
Modeling of Nitrification Kinetics in a Respirometric Biosensor under Suboptimal Conditions

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
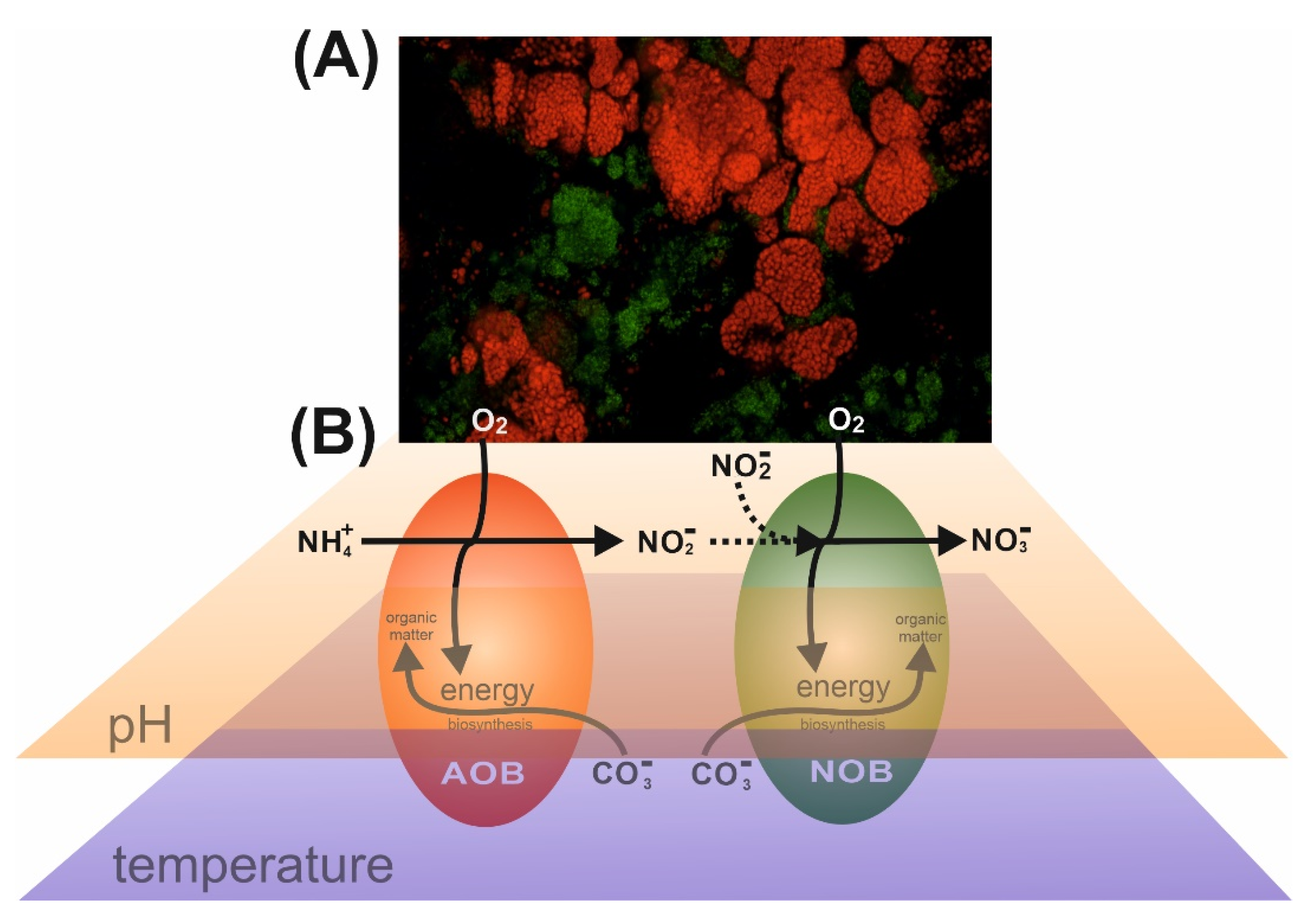
2.1. Immobilization, Growth and Activity Quantification of Bacteria in ABTOW
2.2. Statistical Analysis
3. Results
3.1. Construction of the ABTOW Model
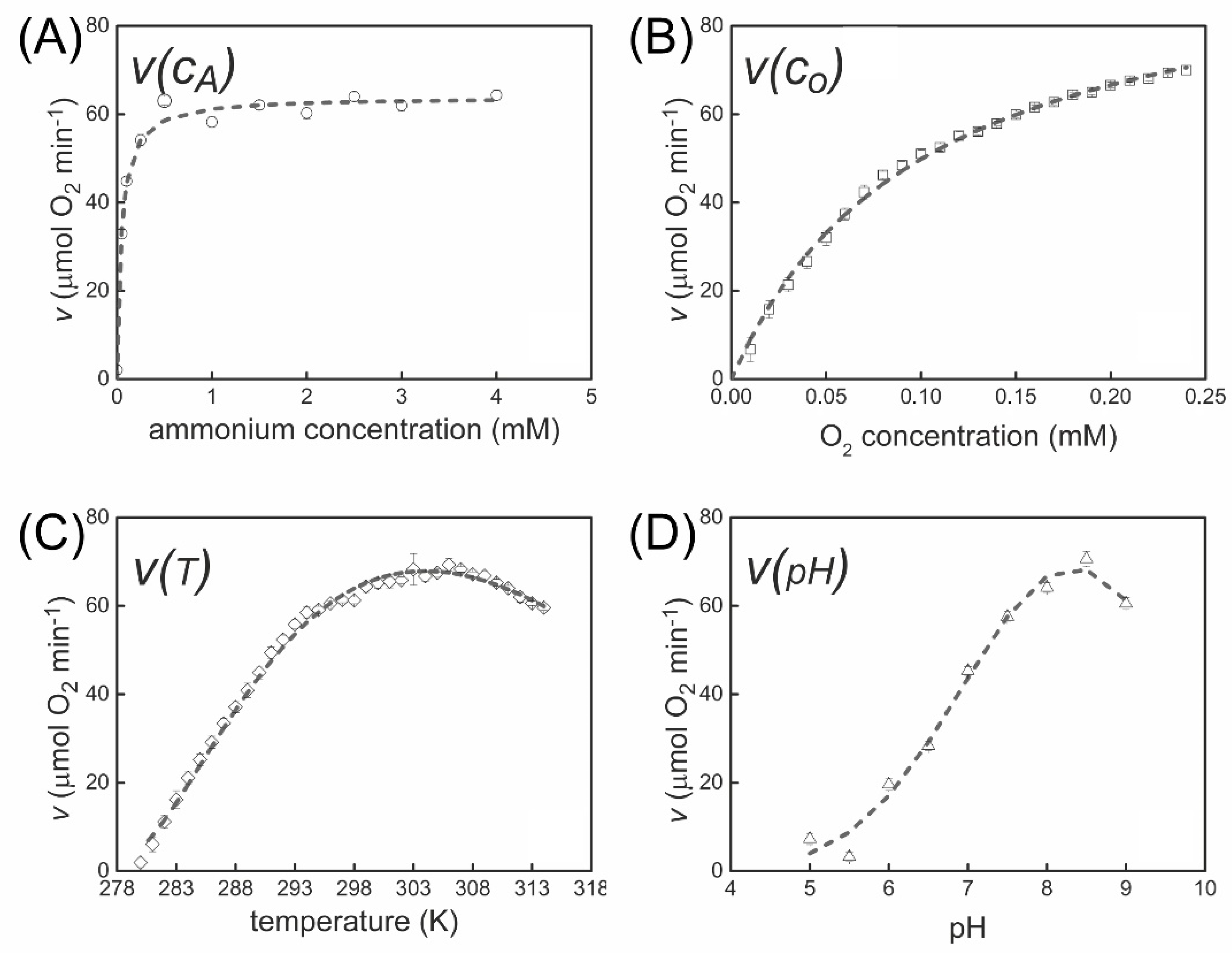
3.2. Dependence of Nitrification Rate on the Substrate Concentration
3.3. Temperature Dependence of Ammonium Oxidation
3.4. Effect of pH on the Ammonium Oxidation
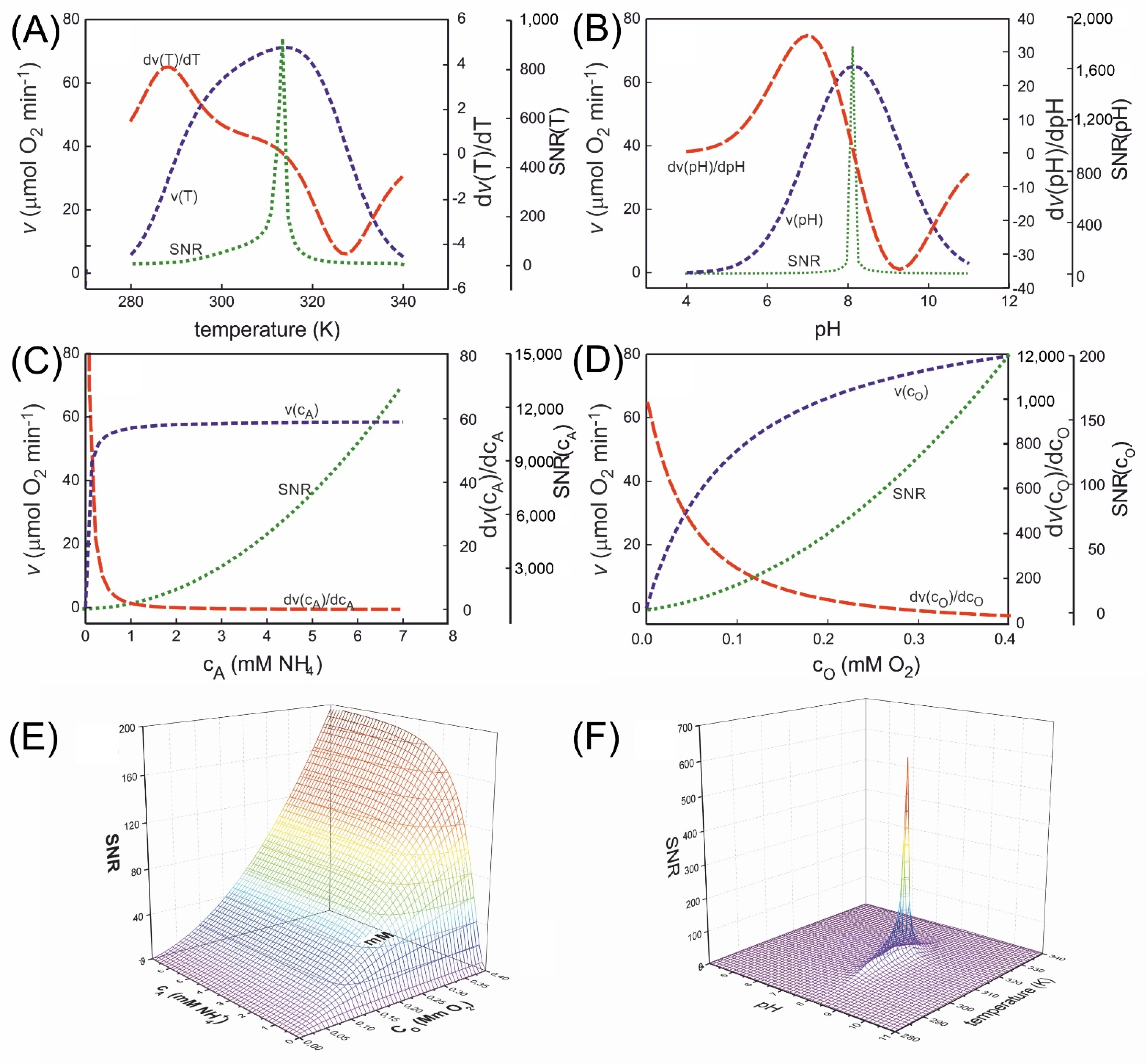
3.5. Dependence of ABTOW Sensitivity on the Stability of Measurement Conditions
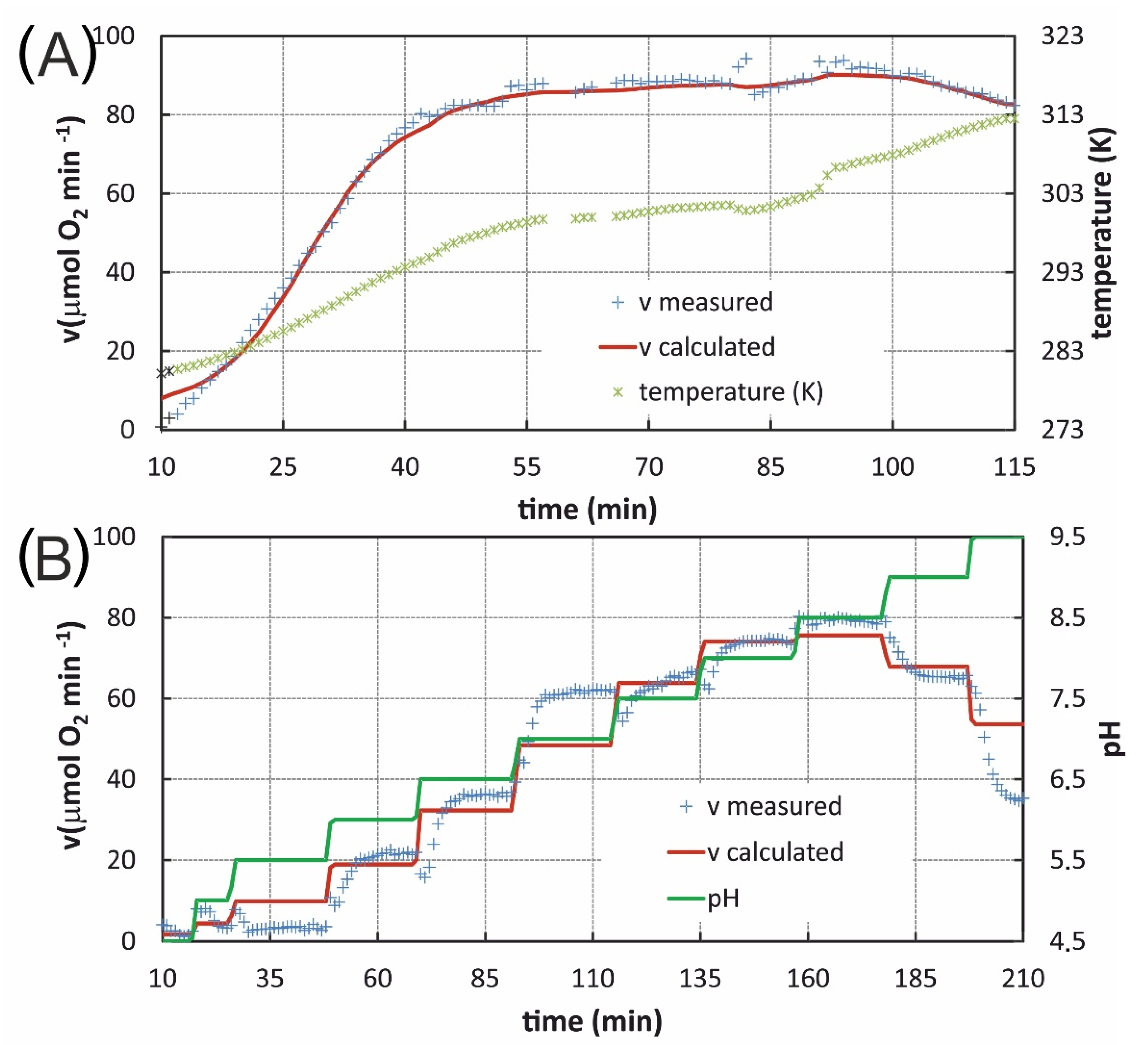
3.6. Performance of the Combined Model in Non-Equilibrium Conditions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woznica, A.; Nowak, A.; Beimfohr, C.; Karczewski, J.; Bernas, T. Monitoring Structure and Activity of Nitrifying Bacterial Biofilm in an Automatic Biodetector of Water Toxicity. Chemosphere 2010, 78, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Woznica, A.; Nowak, A.; Karczewski, J.; Klis, C.; Bernas, T. Automatic Biodetector of Water Toxicity (ABTOW) as a Tool for Examination of Phenol and Cyanide Contaminated Water. Chemosphere 2010, 81, 767–772. [Google Scholar] [CrossRef] [PubMed]
- APHA (Ed.) Standard Methods for the Examination of Water and Wastewater; WPCF: Washington, DC, USA, 1995. [Google Scholar]
- Herschy, R.W. Water Quality for Drinking: WHO Guidelines. In Encyclopedia of Lakes and Reservoirs; World Health Organization: Geneva, Switzerland, 2012; pp. 876–883. [Google Scholar] [CrossRef]
- Eltzov, E.; Slobodnik, V.; Ionescu, R.E.; Marks, R.S. On-Line Biosensor for the Detection of Putative Toxicity in Water Contaminants. Talanta 2015, 132, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Gosset, A.; Ferro, Y.; Durrieu, C. Methods for Evaluating the Pollution Impact of Urban Wet Weather Discharges on Biocenosis: A Review. Water Res. 2016, 89, 330–354. [Google Scholar] [CrossRef]
- Ejeian, F.; Etedali, P.; Mansouri-Tehrani, H.A.; Soozanipour, A.; Low, Z.X.; Asadnia, M.; Taheri-Kafrani, A.; Razmjou, A. Biosensors for Wastewater Monitoring: A Review. Biosens. Bioelectron. 2018, 118, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.M.Z.; Mansour, N. Biosensors for On-Line Water Quality Monitoring-a review. Arab J. Basic Appl. Sci. 2019, 26, 502–518. [Google Scholar] [CrossRef]
- Nichols, D.; Ferguson, S. Bioenergetics, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780123884251. [Google Scholar]
- Spieck, E.; Aamand, J.; Bartosch, S.; Bock, E. Immunocytochemical Detection and Location of the Membrane-Bound Nitrite Oxidoreductase in Cells of Nitrobacter and Nitrospira. FEMS Microbiol. Lett. 1996, 139, 71–76. [Google Scholar] [CrossRef]
- Spieck, E.; Ehrich, S.; Aamand, J.; Bock, E. Isolation and Immunocytochemical Location of the Nitrite-Oxidizing System in Nitrospira Moscoviensis. Arch. Microbiol. 1998, 169, 225–230. [Google Scholar] [CrossRef]
- Woznica, A.; Nowak, A.; Ziemski, P.; Kwasniewski, M.; Bernas, T. Stimulatory Effect of Xenobiotics on Oxidative Electron Transport of Chemolithotrophic Nitrifying Bacteria Used as Biosensing Element. PLoS ONE 2013, 8, e53484. [Google Scholar] [CrossRef]
- Woznica, A.; Karcz, J.; Nowak, A.; Gmur, A.; Bernas, T. Spatial Architecture of Nitrifying Bacteria Biofilm Immobilized on Polyurethane Foam in an Automatic Biodetector for Water Toxicity. Microsc. Microanal. 2010, 16, 550–560. [Google Scholar] [CrossRef]
- Kantartzi, S.G.; Vaiopoulou, E.; Kapagiannidis, A.; Aivasidis, A. Kinetic Characterization of Nitrifying Pure Cultures in Chemostate. Glob. NEST J. 2006, 8, 43–51. [Google Scholar]
- Carrera, J.; Jubany, I.; Carvallo, L.; Chamy, R.; Lafuente, J. Kinetic Models for Nitrification Inhibition by Ammonium and Nitrite in a Suspended and an Immobilized Biomass Systems. Process Biochem. 2004, 39, 1159–1165. [Google Scholar] [CrossRef]
- Jubany, I.; Carrera, J.; Lafuente, J.; Baeza, J.A. Start-up of a Nitrification System with Automatic Control to Treat Highly Concentrated Ammonium Wastewater: Experimental Results and Modeling. Chem. Eng. J. 2008, 144, 407–419. [Google Scholar] [CrossRef]
- Jubany, I.; Lafuente, J.; Baeza, J.A.; Carrera, J. Total and Stable Washout of Nitrite Oxidizing Bacteria from a Nitrifying Continuous Activated Sludge System Using Automatic Control Based on Oxygen Uptake Rate Measurements. Water Res. 2009, 43, 2761–2772. [Google Scholar] [CrossRef]
- Mota, C.; Head, M.A.; Ridenoure, J.A.; Cheng, J.J.; De Los Reyes, F.L. Effects of Aeration Cycles on Nitrifying Bacterial Populations and Nitrogen Removal in Intermittently Aerated Reactors. Appl. Environ. Microbiol. 2005, 71, 8565–8572. [Google Scholar] [CrossRef] [Green Version]
- Wiesmann, U.; Choi, I.S.; Dombrowski, E.M. Nutrient Removal in Fundamentals of Biological Wastewater Treatment Biological; WILEY-VCH: Weinheim, Germany, 2007. [Google Scholar]
- Gerardi, M.H. Nitrification and Denitrification in the Activated Sludge Process; (Google EBook); 2003; ISBN ISBN 0471461318. Available online: https://books.google.vg/books?id=ccIKuXYNlBMC&printsec=frontcover#v=onepage&q&f=false (accessed on 12 May 2022).
- Macdonald, R.M.; Spokes, J.R. A Selective and Diagnostic Medium for Ammonia Oxidising Bacteria. FEMS Microbiol. Lett. 1980, 8, 143–145. [Google Scholar] [CrossRef]
- Jokić, I.; Djurić, Z.; Radulović, K.; Frantlović, M.; Milovanović, G.V.; Krstajić, P.M. Stochastic Time Response and Ultimate Noise Performance of Adsorption--based Microfluidic Biosensors. Biosensors 2021, 11, 194. [Google Scholar] [CrossRef]
- Karcz, J.; Bernas, T.; Nowak, A.; Talik, E.; Woznica, A. Application of Lyophilization to Prepare the Nitrifying Bacterial Biofilm for Imaging with Scanning Electron Microscopy. Scanning 2012, 34, 26–36. [Google Scholar] [CrossRef]
- Zwietering, M.H.; de Koos, J.T.; Hasenack, B.E.; de Witt, J.C.; van’t Riet, K. Modeling of Bacterial Growth as a Function of Temperature. Appl. Environ. Microbiol. 1991, 57, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Rinken, T.; Rinken, A.; Tenno, T.; Järv, J. Calibration of Glucose Biosensors by Using Pre-Steady State Kinetic Data. Biosens. Bioelectron. 1998, 13, 801–807. [Google Scholar] [CrossRef]
- Van Hulle, S.W.; Volcke, E.I.; Teruel, J.L.; Donckels, B.; van Loosdrecht, M.C.; Vanrolleghem, P. a Influence of Temperature and PH on the Kinetics of the Sharon Nitritation Process. J. Chem. Technol. Biotechnol. 2007, 82, 471–480. [Google Scholar] [CrossRef]
- Davidson, E.A.; Samanta, S.; Caramori, S.S.; Savage, K. The Dual Arrhenius and Michaelis-Menten Kinetics Model for Decomposition of Soil Organic Matter at Hourly to Seasonal Time Scales. Glob. Chang. Biol. 2012, 18, 371–384. [Google Scholar] [CrossRef]
- Rosso, L.; Lobry, J.R.; Bajard, S.; Flandrois, J.P. Convenient Model to Describe the Combined Effects of Temperature and pH on Microbial Growth. Appl. Environ. Microbiol. 1995, 61, 610–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equation Parameter | Symbol | Value | Unit |
|---|---|---|---|
| maximum oxygen consumption rate | Vmax | 70.62 ± 6.9 | μmol⋅min−1 |
| O2 Michaelis constant | KO | 0.1 | mM |
| NH4+ Michaelis constant | KA | 0.04 | mM |
| optimal pH | pHopt | 8.15 ± 0.05 | - |
| difference between high pH and low pH when v is equal to 0.5 VpH | U | 1.9 ± 0.09 | - |
| the activation energy of AOB nitrification process | Ea | 183 ± 23 | kJ⋅mol−1 |
| the activation energy of low-temperature enzyme denaturation | El | 177 ± 22 | kJ⋅mol−1 |
| the activation energy of high-temperature enzyme denaturation | Eh | 378 ± 21 | kJ⋅mol−1 |
| the optimal temperature of the process | To | 298 | K |
| the temperature of low-temperature denaturation | Tl | 287 ± 0.7 | K |
| the temperature of high-temperature denaturation | Th | 307 ± 12 | K |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woznica, A.; Karczewski, J.; Klis, C.; Długosz, J.; Ziemski, P.; Nowak, A.; Bernas, T. Modeling of Nitrification Kinetics in a Respirometric Biosensor under Suboptimal Conditions. Water 2022, 14, 2031. https://doi.org/10.3390/w14132031
Woznica A, Karczewski J, Klis C, Długosz J, Ziemski P, Nowak A, Bernas T. Modeling of Nitrification Kinetics in a Respirometric Biosensor under Suboptimal Conditions. Water. 2022; 14(13):2031. https://doi.org/10.3390/w14132031
Chicago/Turabian StyleWoznica, Andrzej, Jerzy Karczewski, Czesław Klis, Jacek Długosz, Przemysław Ziemski, Agnieszka Nowak, and Tytus Bernas. 2022. "Modeling of Nitrification Kinetics in a Respirometric Biosensor under Suboptimal Conditions" Water 14, no. 13: 2031. https://doi.org/10.3390/w14132031
APA StyleWoznica, A., Karczewski, J., Klis, C., Długosz, J., Ziemski, P., Nowak, A., & Bernas, T. (2022). Modeling of Nitrification Kinetics in a Respirometric Biosensor under Suboptimal Conditions. Water, 14(13), 2031. https://doi.org/10.3390/w14132031