

Modeling Stygofauna Resilience to the Impact of the Climate Change in the Karstic Groundwaters of South Italy


Abstract
:1. Introduction
2. Materials and Methods
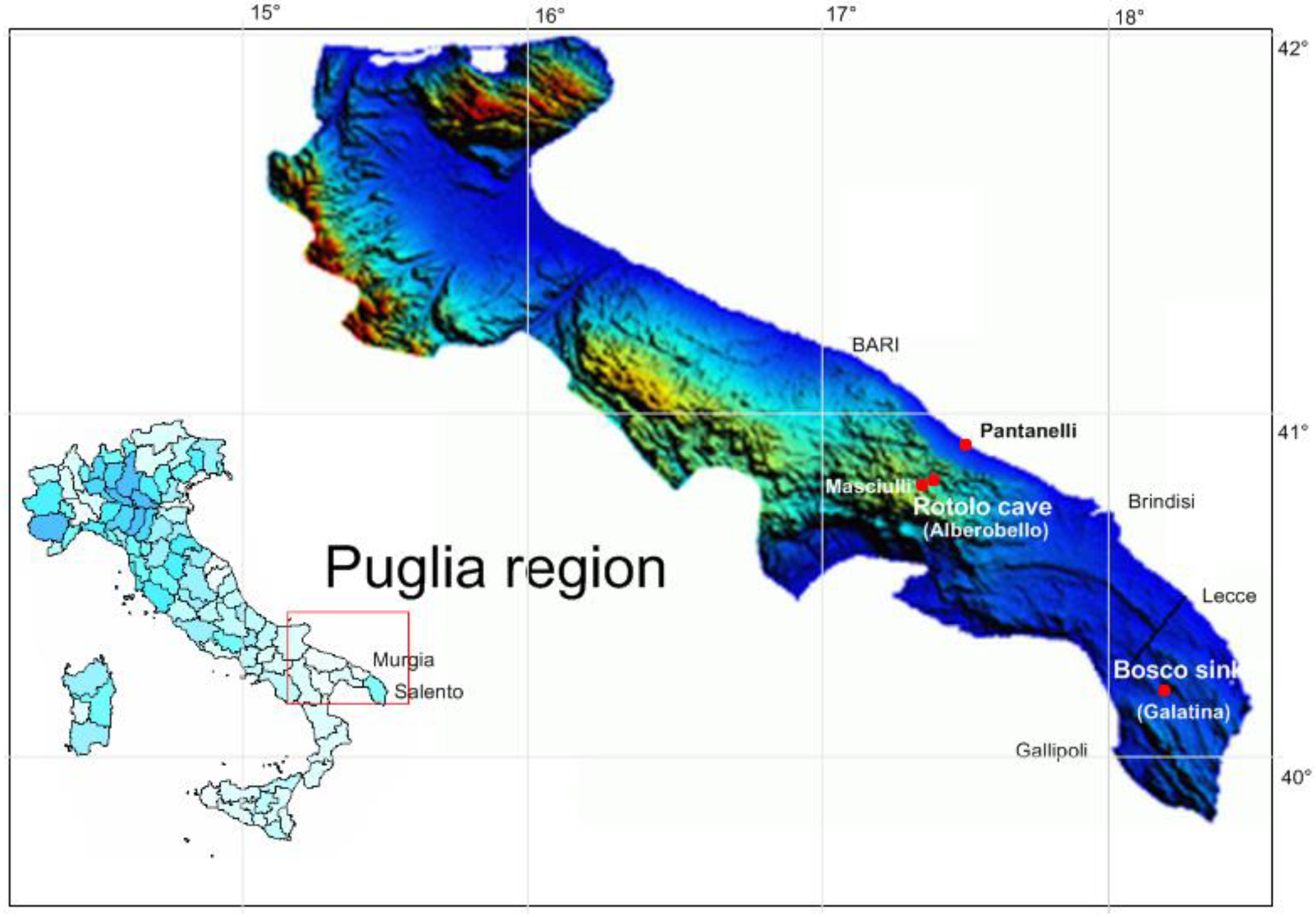
2.1. Sampled Habitats
2.2. Assessment of Increases in Groundwater Temperature and Salinity
3. Results
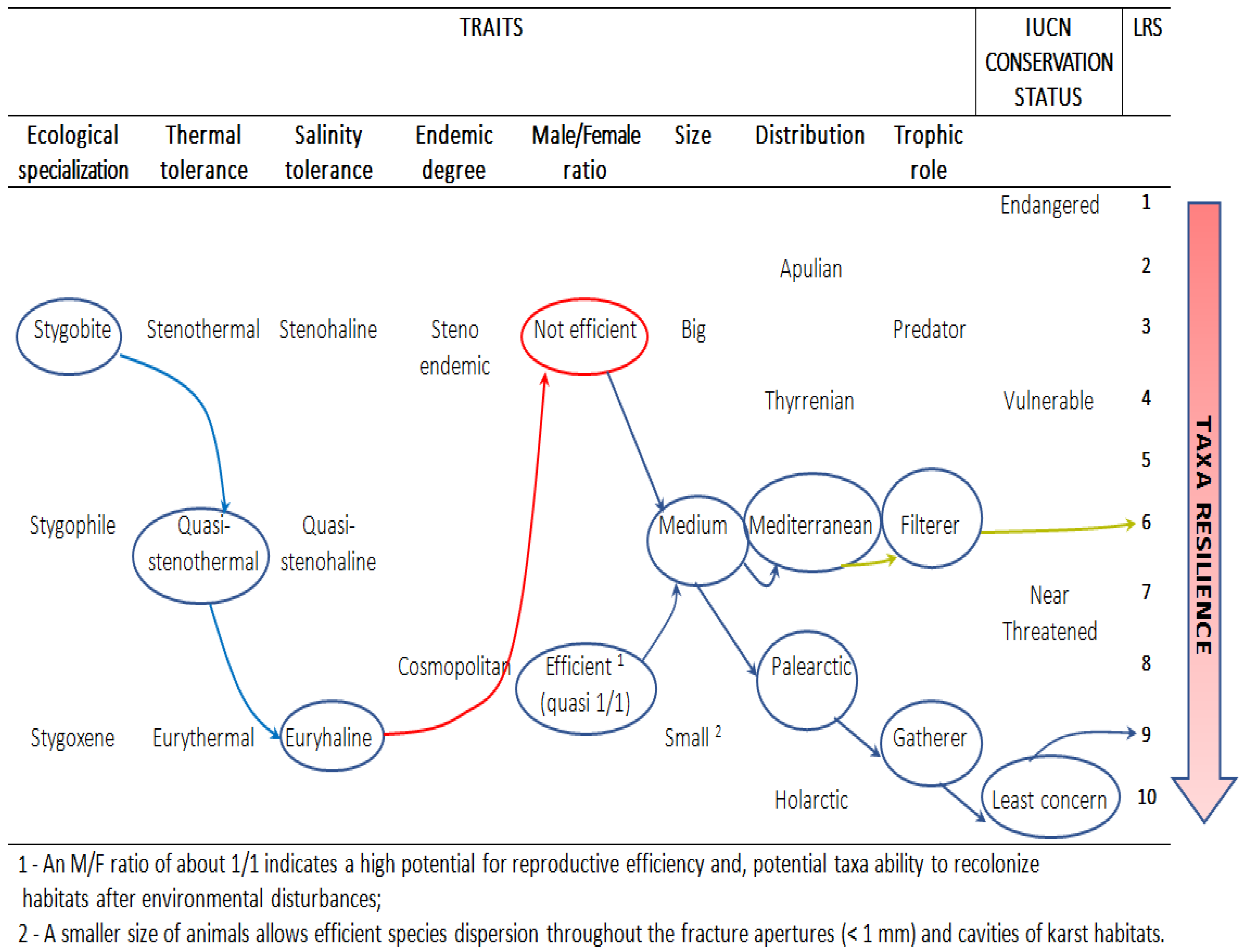
3.1. Resilience of Stygofauna to Climate Change
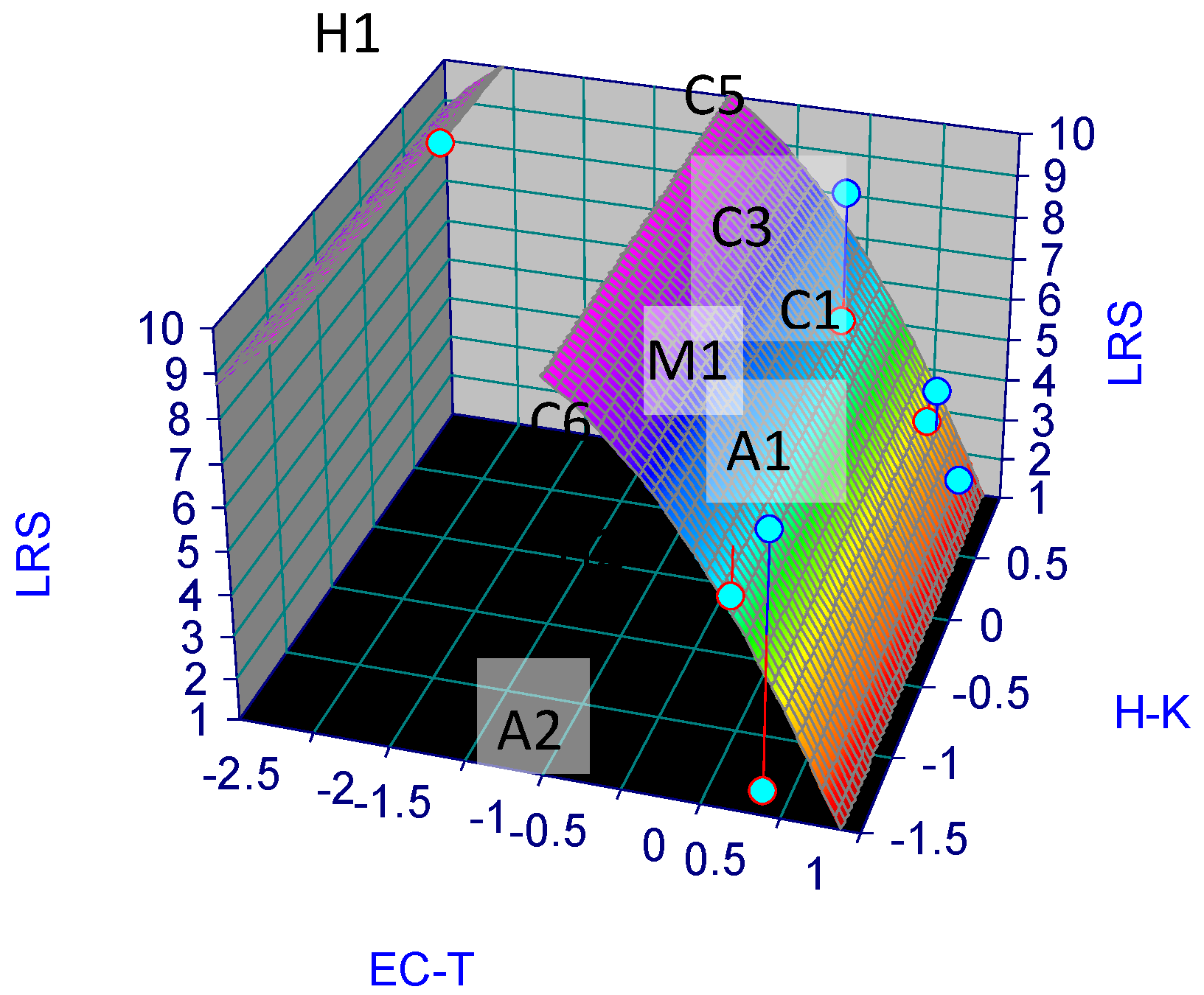
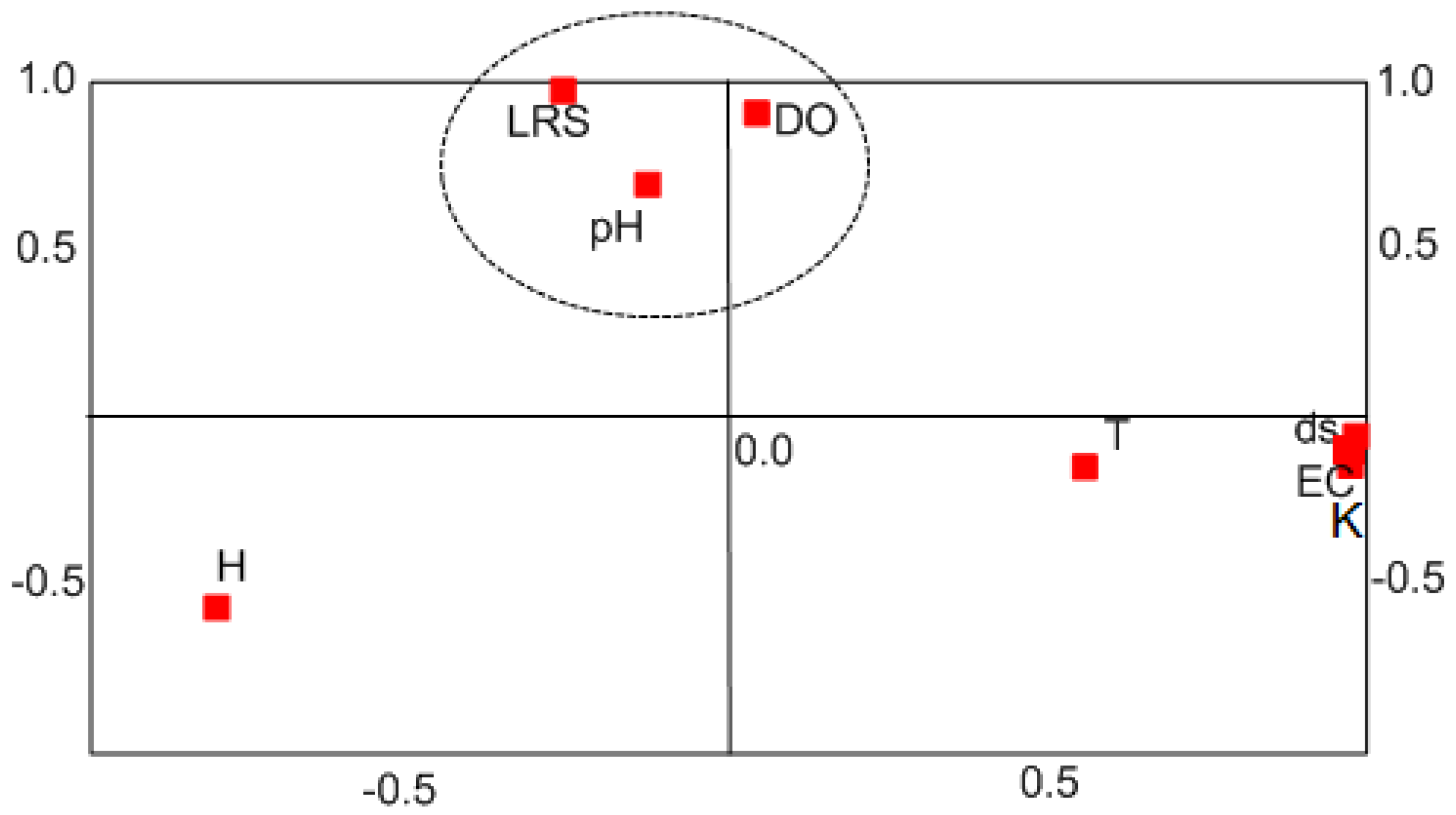
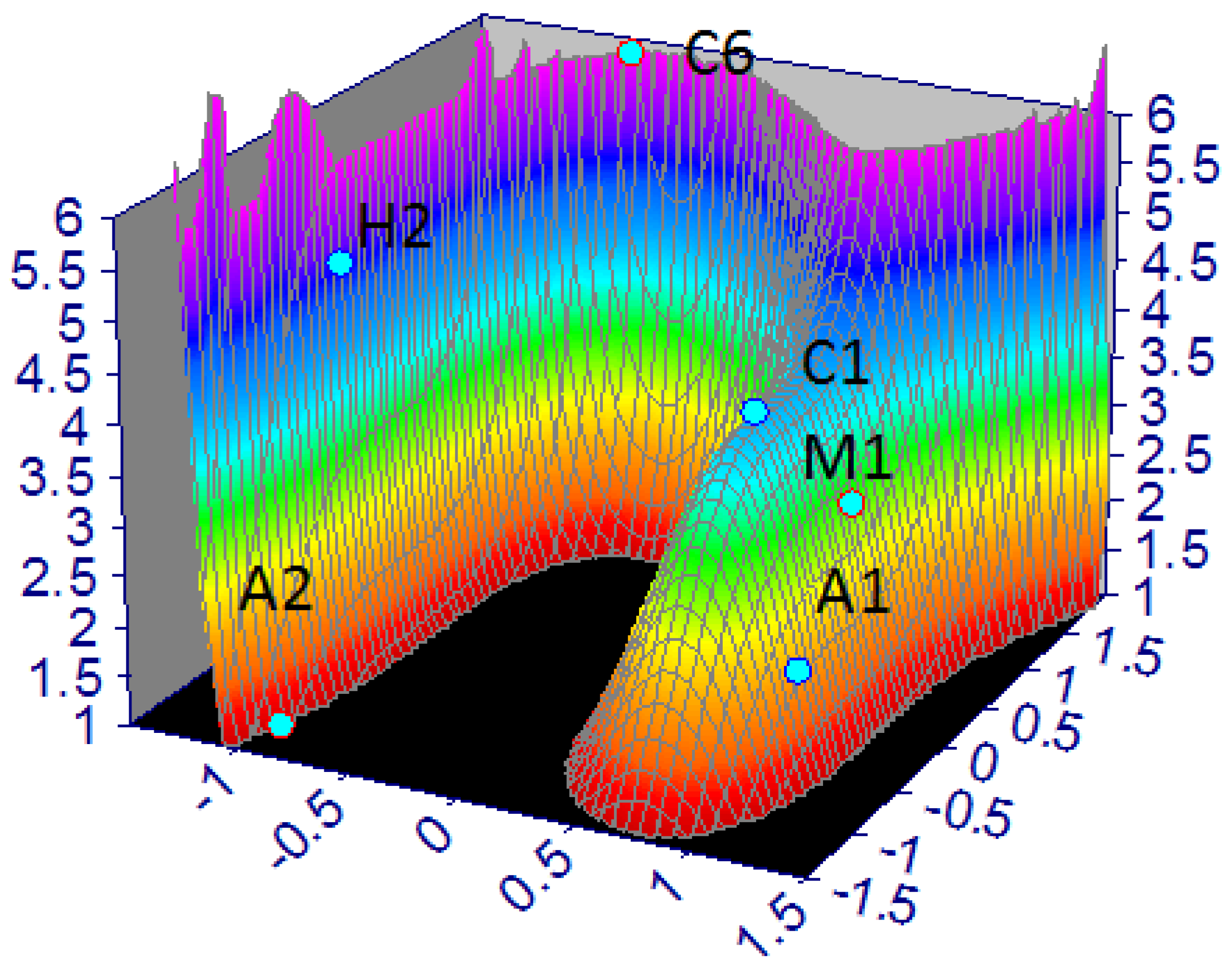
3.2. Mathematical Model of Stygofauna Resilience
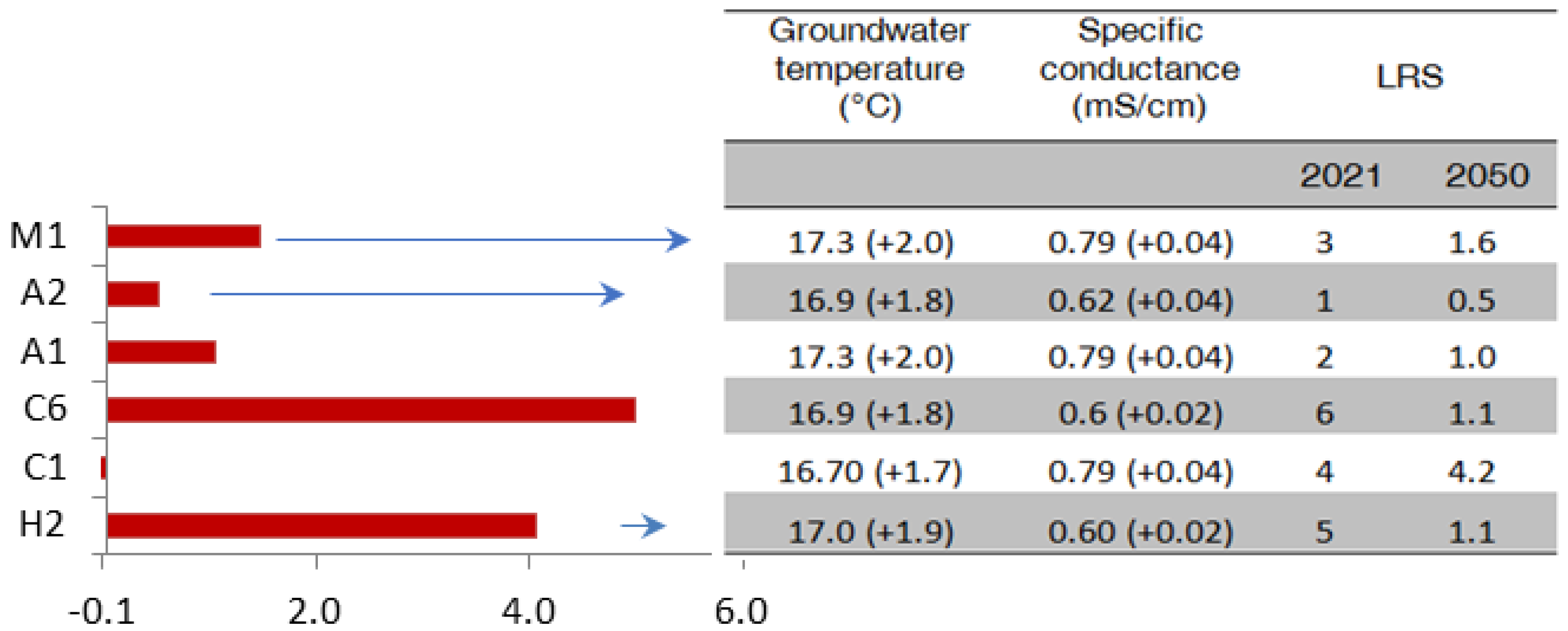
3.3. Projections of Stygofaunal Resilience for 2050
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Domínguez-Villar, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Christman, M.C.; Culver, D.C. The relationship between cave biodiversity and available habitat. J. Biogeogr. 2001, 28, 367–380. [Google Scholar] [CrossRef]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Danielopol, D.L.; Rouch, R.; Baltanas, A. Taxonomic diversity of groundwater harpacticoida (Copepoda, Crustacea) in southern France—A contribution to characterise hotspot diversity sites. Vie Milieu-Life Environ. 2002, 52, 1–15. [Google Scholar]
- Zagmajster, M.; Malard, F.; Eme, D.; Culver, D.C. Subterranean biodiversity patterns from global to regional scales. In Cave Ecology; Moldovan, O.T., Kovàc, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 201. [Google Scholar] [CrossRef]
- Walters, A.D.; Cannizzaro, A.G.; Trujillo, D.A.; Berg, D.J. Addressing the Linnean shortfall in a cryptic species complex. Zool. J. Linn. Soc. 2021, 192, 277–305. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.P.; Griebler, C.; Halse, S.; Humphreys, W.F.; et al. Scientists’ Warning on the Conservation of Subterranean Ecosystems. BioScience 2019, 69, 641–650. [Google Scholar] [CrossRef]
- Giere, O. Introduction to Meiobenthology. In Meiobenthology; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar] [CrossRef]
- Billi, A.; Valle, A.; Brilli, M.; Faccenna, C.; Funiciello, R. Fracture-controlled fluid circulation and dissolutional weathering in sinkhole-prone carbonate rocks from central Italy. J. Struct. Geol. 2007, 29, 385–395. [Google Scholar] [CrossRef]
- Hancock, P.J.; Boulton, A.J.; Humphreys, W.F. Aquifers and hyporheic zones: Towards an ecological understanding of groundwater. Hydrogeol. J. 2005, 13, 98–111. [Google Scholar] [CrossRef]
- Humphreys, W.F. Aquifers: The ultimate groundwater-dependent ecosystems. Aust. J. Bot. 2006, 54, 115–132. [Google Scholar] [CrossRef]
- Achurra, A.; Rodriguez, P. Biodiversity of groundwater oligochaetes from a karst unit in northern Iberian Peninsula: Ranking subterranean sites for conservation management. Hydrobiologia 2008, 605, 159. [Google Scholar] [CrossRef]
- Pipan, T.; Navodnik, V.; Janzekovic, F.; Novak, T. Studies of the fauna of percolation water of Huda luknja, a cave in isolated karst in northeast Slovenia. Acta Carsologica 2008, 37, 141–151. [Google Scholar] [CrossRef]
- Masciopinto, C.; Semeraro, F.; La Mantia, R.; Inguscio, S.; Rossi, E. Stygofauna abundance and distribution in the fissures and caves of the Nardò (S Italy) fractured aquifer subject to reclaimed water injections. Geomicrobiol. J. 2006, 23, 267–278. [Google Scholar] [CrossRef]
- Masciopinto, C.; Liso, I.S. Assessment of the impact of sea-level rise due to climate change on coastal groundwater discharge. Sci. Total Environ. 2015, 569–570, 672–680. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Fiasca, B.; Tabilio Di Camillo, A.; Murolo, A.; Di Cicco, M.; Galassi, D.M.P. The weighted groundwater health index(wGHI) by Korbel and Hose (2017) in European groundwater bodies in nitrate vulnerable zones. Ecol. Indic. 2020, 116, 106525. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Borgoni, R.; Ambrosini, R.; Cifoni, M.; Galassi, D.; Petitta, M. Occurrence of volatile organic compounds in shallow alluvial aquifers of a Mediterranean region: Baseline scenario and ecological implications. Sci. Total Environ. 2015, 538, 712–723. [Google Scholar] [CrossRef]
- Blowes, S.A.; Supp, S.R.; Antão, L.H.; Bates, A.; Bruelheide, H.; Chase, J.M.; Moyes, F.; Magurran, A.; McGill, B.; Myers-Smith, I.H.; et al. The geography of biodiversity change in marine and terrestrial assemblages. Science 2019, 366, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Castellarini, F.; Malard, F.; Dole-Olivier, M.-J.; Gibert, J. Modelling the distribution of stygobionts in the Jura Mountains (Eastern France). Implications for the protection of groundwaters. Divers. Distrib. 2007, 13, 213–224. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, ecology and evolution of groundwater copepods. Freshw. Biol. 2009, 54, 691–708. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Galassi, D. Effect of temperature rising on the stygobitic crustacean species Diacyclops belgicus: Does global warming affect groundwater populations? Water 2017, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- Peck, L.S.; Clark, M.S.; Morley, S.A.; Massey, A.; Rossetti, H. Animal temperature limits and ecological relevance: Effects of size, activity and rates of change. Funct. Ecol. 2009, 23, 248–256. [Google Scholar] [CrossRef]
- SPSS 1995. SPSS for Windows; Release 6.1 1996; SPSS Inc.: Chicago, IL, USA, 1995; Volume 60611. [Google Scholar]
- Maggiore, M.; Pagliarulo, P. Circolazione Idrica ed Equilibri Idrogeologici Negli Acquiferi della Puglia. Geologi e Territorio; Supplemento al n. 1/2004; Periodico dell’ Ordine dei Geologi della Puglia: Bari, Italy, 2004. [Google Scholar]
- Parise, M.; Benedetto, L. Surface Landforms and Speleological Investigation for a Better Understanding of Karst Hydrogeological Processes: A History of Research in Southeastern Italy; Geological Society: London, UK, 2018; Special Publications 466:1; pp. 137–153. [Google Scholar]
- Bauer, S.; Liedl, R.; Sauter, M. Modeling the influence of epikarst evolution on karst aquifer genesis: A time variant recharge boundary condition for joint karst-epikarst development. Water Resour. Res. 2005, 41, W09416. [Google Scholar] [CrossRef]
- Cvetkov, L. Un filet phréatobiologique. Bull. De L’institut De Zool. Et Musée Sofia 1968, 22, 215–219. [Google Scholar]
- Watson, N.T.N.; Duggan, I.C.; Hogg, I.D. Assessing the diversity of New Zealand freshwater harpacticoid copepods (Crusta cea: Copepoda) using mitochondrial DNA (COI) barcodes. N. Z. J. Zool. 2015, 42, 57–67. [Google Scholar] [CrossRef]
- Dussart, B. Les Copépodes des Eaux Continentales; Editions N. Boublée & Cie: Paris, France, 1967; Volume I. [Google Scholar]
- Dussart, B. Les Copépodes des Eaux Continentales; Editions N. Boublée & Cie: Paris, France, 1969; Volume II. [Google Scholar]
- Dussart, B.; Defaye, D. World Directory of Crustacea Copepoda of Inland Waters, II—Cyclopiformes, Backhuys; Publishers BV: Leiden, The Netherlands, 2006; p. 354. [Google Scholar]
- Messouli, M.; Coineau, N.; Boutin, C. Revision, Phylogeny and Biogeography of the Groundwater Amphipods Salentinellidae. I. Description of Salentinella anae nov. sp. from Spain with Remarks on the Genera Salentinella and Parasalentinella. Zool. Sci. 2002, 19, 147–1154. [Google Scholar] [CrossRef] [PubMed]
- Nouvel, H.; Casanova, J.-P.; Lagardère, J.-P. Ordre des Mysidacés (Mysidacea Boas 1883). In Traité de Zoologie. Anatomie, Systématique, Biologie. Tome VII, Fascicule IIIA. Crustacés Péracarides; Forest, J., Ed.; Memoires de l’Institut Oceanographique Fondation Albert Ier: Prince de Monaco, Monaco, 1999; Volume 19, pp. 39–86. [Google Scholar]
- Ruffo, S. Hadzia minuta n. sp. (Hadziidae) e Salentinella gracillima n.gen., n. sp. (Gammaridae) nuovi anfipodi troglobi dell’Italia meridionale. Bollettino Soc. Nat. Napoli. 1947, 56, 178–188. [Google Scholar]
- Thorp, J.H.; Rogers, D.C. Thorp & Covich’s Freshwater Invertebrates, 4th ed.; Ecology and General Biology; Academic Press: Cambridge, MA, USA, 2015; Volume I. [Google Scholar]
- Wellborn, G.A.; Witt, J.D.S.; Cothran, R.D. Class Malacostraca, Superorders Peracarida and Syncarida; Thorp and Covich’s Freshwater Invertebrates; Academic Press: Cambridge, MA, USA, 2015; pp. 781–796. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change). Climate Change 2013: The Physical Science Basis; Working Group I Contribution to the IPCC Fifth Assessment Report; Cambridge University Press: Cambridge, UK, 2013; Available online: www.ipcc.ch/report/ar5/wg1 (accessed on 9 June 2022).
- Kano, Y.; Kase, T. Genetic exchange between anchialine cave populations by means of larval dispersal: The case of a new gastropod species Neritilia cavernicola. Zool. Scr. 2004, 33, 423–437. [Google Scholar] [CrossRef]
- Gonzalez, B.C.; Martínez, A.; Borda, E.; Iliffe, T.M.; Fontaneto, D.; Worsaae, K. Genetic spatial structure of an anchialine cave annelid indicates connectivity within—But not between—Islands of the Great Bahama bank. Mol. Phylogenetics Evol. 2017, 109, 259–270. [Google Scholar] [CrossRef]
- Latella, L.; Ruffo, S.; Stoch, F. The project CKmap (Checklist and Distribution of the Italian Fauna): Methods and Informatical Techniques. Memorie del Museo Civico di Storia Naturale di Verona, 15–19. Accademia.edu. 2007. Available online: https://scholar.google.it/scholar?q=IUCN+(Latella+et+al.,+2007&hl=it&as_sdt=0&as_vis=1&oi=scholart (accessed on 9 June 2022).
- Minelli, A.; Ruffo, S. Checklist delle Specie della Fauna Italiana. Calderini, Bologna, Fascicoli 1–110; La Posta, S., Ed.; Bologna, Municipality of Verona: Verona, Italy, 1995. [Google Scholar]
- Ruffo, S.; Stoch, F. Checklist e distribuzione della fauna italiana. Memorie del Museo Civico di Storia Naturale di Verona, II Serie, Sezione Scienze della Vita 16; Municipality of Verona: Verona, Italy, 2005. [Google Scholar]
- Vesentini, G. Arpacticoidi (Crostacei Copepodi) del lago Maggiore, del lago di Mergozzo e di piccole acque viciniori. Mem. Dell’istitituto Ital. Di Idrobiol. 1968, 24, 197–224. [Google Scholar]
- Shapouri, M.; Cancela da Fonseca, L.; Iepure, S.; Stigter, T.; Ribeiro, L.; Silva, A. The variation of stygofauna along a gradient of salinization in a coastal aquifer: The variation of stygofauna along a gradient of salinization in coastal aquifer. Hydrol. Res. 2015, 47, 89–103. [Google Scholar] [CrossRef]
- Castaño-Sánchez, A.; Hose, G.C.; Reboleira, A.S.P.S. Salinity and temperature increase impact groundwater crustaceans. Sci. Rep. 2020, 10, 12328. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, G. A New Species of Pseudocyclopiidae (Crustacea, Copepoda, Calanoida) From an Anchialine Environment of South-Eastern Italy. Front. Ecol. Evol. 2022, 10, 857161. [Google Scholar] [CrossRef]
- Ciccarese, G.; Pesce, G.L. La Zinzulusa: 200 anni dopo. Thalass. Salentina 1999, 23, 79–88. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Locations | Longitude (N) | Latitude (E) | Hydraulic Conductivity (cm/s) | Distance from Sea (km) | Flow Gradient (%) | pH | Temperature (°C) | Electric Conductivity (mS/cm) | O2 (mg/L) | Flow Head (m a.s.l.) | Soil Elevation(m a.s.l.) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Masciulli (Alberobello, BA) | 40° 49′ 20″ | 17° 15′ 04″ | 0.15 | 11.3 | 0.31 | 7.6 | 15.7 | 0.65 | 5 | 35 | 305 |
| Rotolo cave (Alberobello, BA) | 40° 49′ 30″ | 17° 15′ 08″ | 0.16 | 11 | 0.32 | 7.8 | 15.1 | 0.58 | 5.4 | 35 | 304 |
| Pantanelli (Monopoli, BA) | 42° 53′ 40″ | 17° 23′ 04″ | 0.2 | 0.2 | 0.05 | 8.1 | 11.0 | 8.3 | 7 | 0.1 | 0.5 |
| Bosco sinkhole (Noha, Galatina, LE) | 40° 10′ 1.5″ | 18° 10′ 15″ | 0.29 | 13.4 | 0.02 | 7.6 | 15.0 | 0.752 | 6 | 2.5 | 75 |
| Order | Species | 2018–2021 | Category | Ecological Status Concern | Adult/Young | Fem./Male Length (mm) | Male/Female | Endemic Degree | Coro Type | Trophic Role | Temp. Tolerance | Salinity Tolerance |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crustacea Copepoda Harpacticoida | H1 | Ponds of epikarst in Rotolo cave | Sf | Low | 22/0 | 0.6–0.68/0.58 | 11/11 | Low | EU | Gatherer | High | High |
| H2 | Shaft outflow in Rotolo cave | Sb | 36/0 | 0.42 | - | High | Italy | - | Low | Low (fresh water) | ||
| H3 | Aquifer (Masciulli well) | Sg | Low | 1/0 | 0.7/0.5 | 0/1 | Low | Olartic | Gat. | High | High | |
| Crustacea Copepoda Cyclopoida | C1 | Aquifer (Bosco) | Sb | Vulnerable | 7 | 0.62–0.64/0.50-0.9 | - | High | Med sea | Gat. | Low | Strong |
| C2 | Ponds and aquifer in Rotolo cave | Sg | Low | 67/5 | 0.9–1.4/1 | 37/30 | Low | Olartic | Pred./gat | Very high | High | |
| C3 | Ponds and aquifer in Rotolo cave | Sf | Low | 4/0 | 0.7–1/0.7–0.9 | 3/1 | Low | EU | Gatherer | Quasi low | High | |
| C4 | Aquifer (Bosco) | Sg | Low | 1 | 0.9–1/0.7–0.9 | 0/1 | Low | Asia-EU | Gat. | High | High | |
| C5 | Aquifer (Pantanelli) | Sf | Low | 1/12 | 0.9–1.6/0.8–1.0 | 1/0 | Low | Olartic | Pred./gat. | Very high | High | |
| C6 | Ponds in epikarst of Rotolo cave | Sb | Low | 4 | - | 4/1 | - | - | - | High | High | |
| Amphipoda | A1 | Aquifer (Bosco) | Sb | Endangered | 6/1 | 1.5–2.0 | - | High | Med Sea | Gat. | High | Quasi low |
| A2 | Aquifer (Rotolo cave) | Sb | Quasi threatened | 1/0 | 2.8–4.7 | 1 | High | Italy | Gat. | - | Low (fresh water) | |
| Mysidacea | M1 | Aquifer (Bosco) | Sb | Endangered | 9 | 6.5–13 | 9 | High | Italy | Fit./Saproph age | High | High |
| LRS (Z) | X (EC-T) | Y (H-K) | |
|---|---|---|---|
| H1 | 9 | −0.011 | 0.629 |
| H2 | 5 | 0.061 | −1.232 |
| C1 | 4 | 0.584 | 0.816 |
| C3 | 7 | 0.330 | −1.346 |
| C5 | 10 | −2.565 | 0.212 |
| C6 | 6 | −0.012 | 0.613 |
| A1 | 2 | 0.776 | 0.770 |
| A2 | 1 | 0.330 | −1.346 |
| M1 | 3 | 0.507 | 0.884 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabilio Di Camillo, A.; Masciopinto, C. Modeling Stygofauna Resilience to the Impact of the Climate Change in the Karstic Groundwaters of South Italy. Water 2022, 14, 2715. https://doi.org/10.3390/w14172715
Tabilio Di Camillo A, Masciopinto C. Modeling Stygofauna Resilience to the Impact of the Climate Change in the Karstic Groundwaters of South Italy. Water. 2022; 14(17):2715. https://doi.org/10.3390/w14172715
Chicago/Turabian StyleTabilio Di Camillo, Agostina, and Costantino Masciopinto. 2022. "Modeling Stygofauna Resilience to the Impact of the Climate Change in the Karstic Groundwaters of South Italy" Water 14, no. 17: 2715. https://doi.org/10.3390/w14172715
APA StyleTabilio Di Camillo, A., & Masciopinto, C. (2022). Modeling Stygofauna Resilience to the Impact of the Climate Change in the Karstic Groundwaters of South Italy. Water, 14(17), 2715. https://doi.org/10.3390/w14172715