Exploring the Cumulative Selectivity of Polycyclic Aromatic Hydrocarbons in Phytoplankton, Water, and Sediment in Typical Urban Water Bodies

Abstract
:1. Introduction
2. Materials and Methods
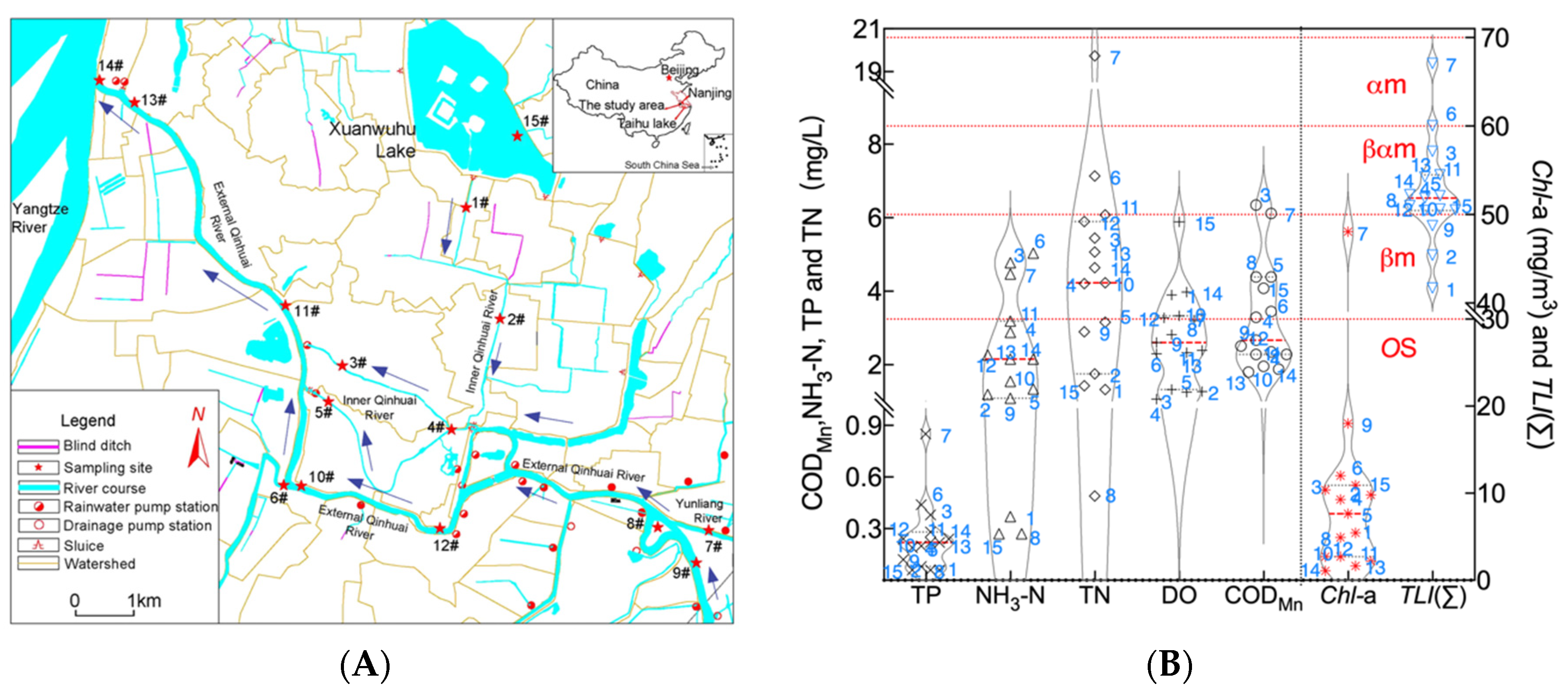
2.1. Sample Collection and Pretreatment
2.2. Data Analysis
3. Results
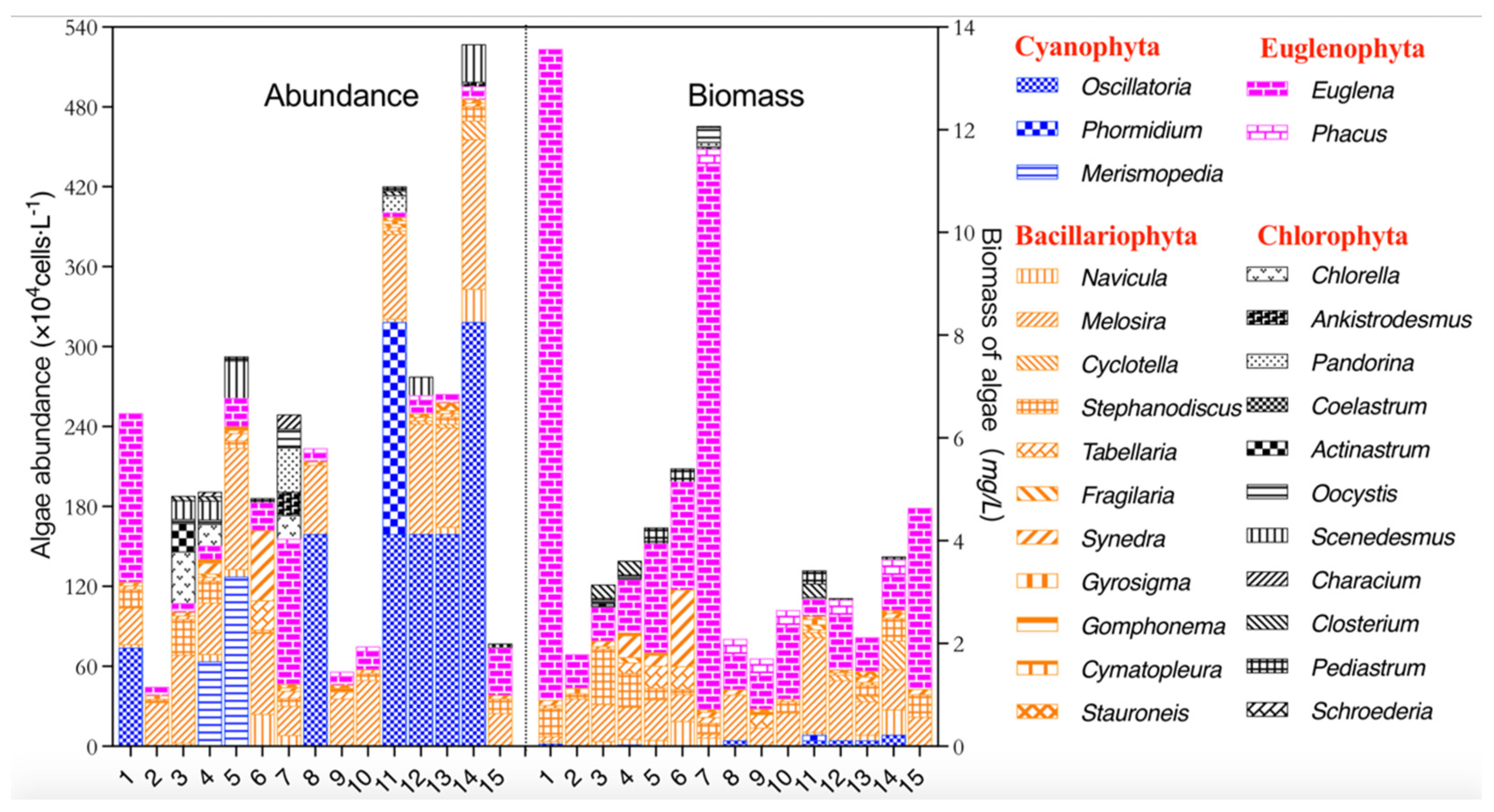
3.1. Characteristics of Phytoplankton Community Composition in Autumn
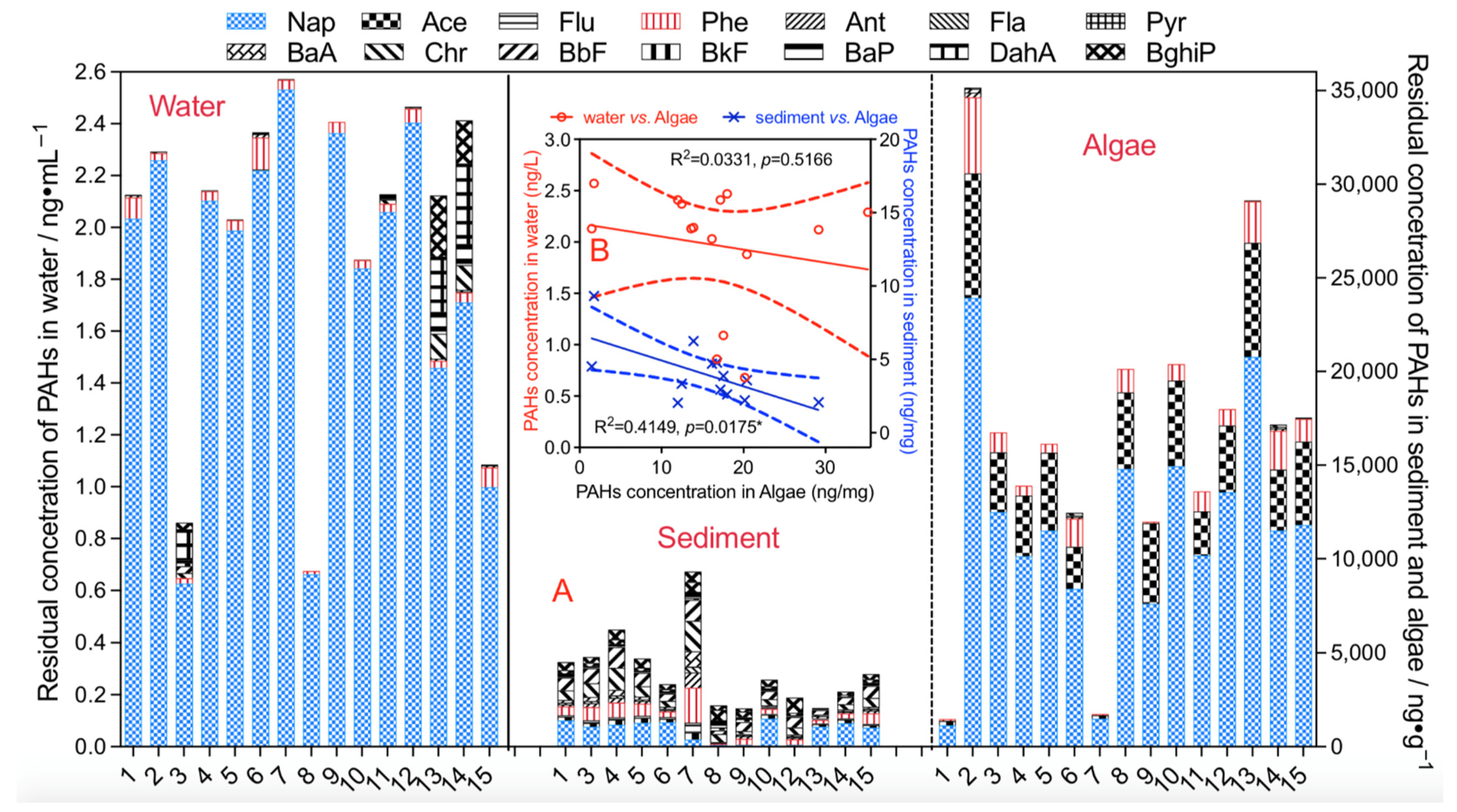
3.2. Residual Characteristics of PAHs in Phytoplankton, Water, and Sediment
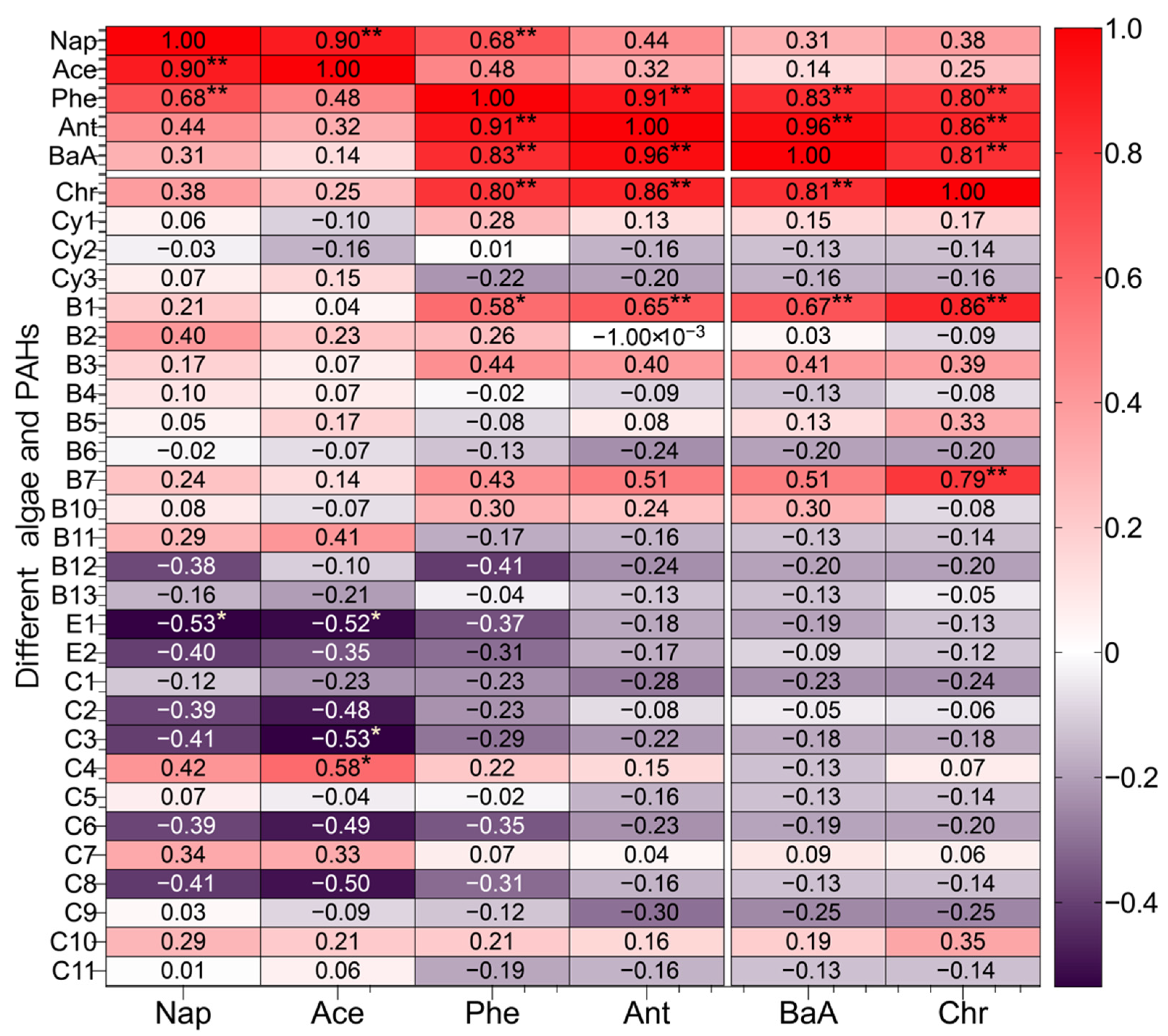
3.3. Relationship Analysis between Phytoplankton Community and Residual PAHs
3.3.1. Correlation Analysis
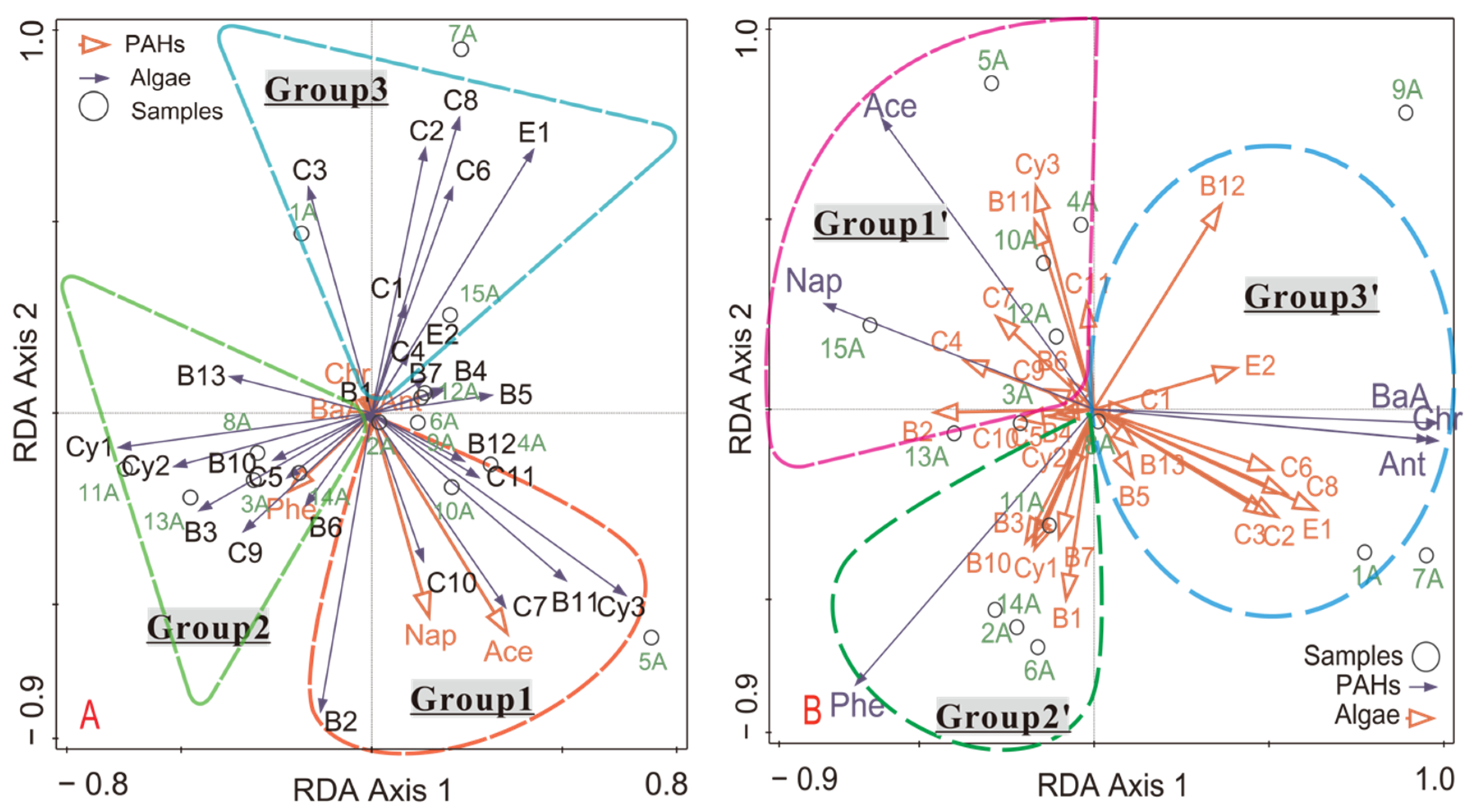
3.3.2. Redundancy Analysis (RDA)
3.3.3. Stepwise Multiple Regression Analysis
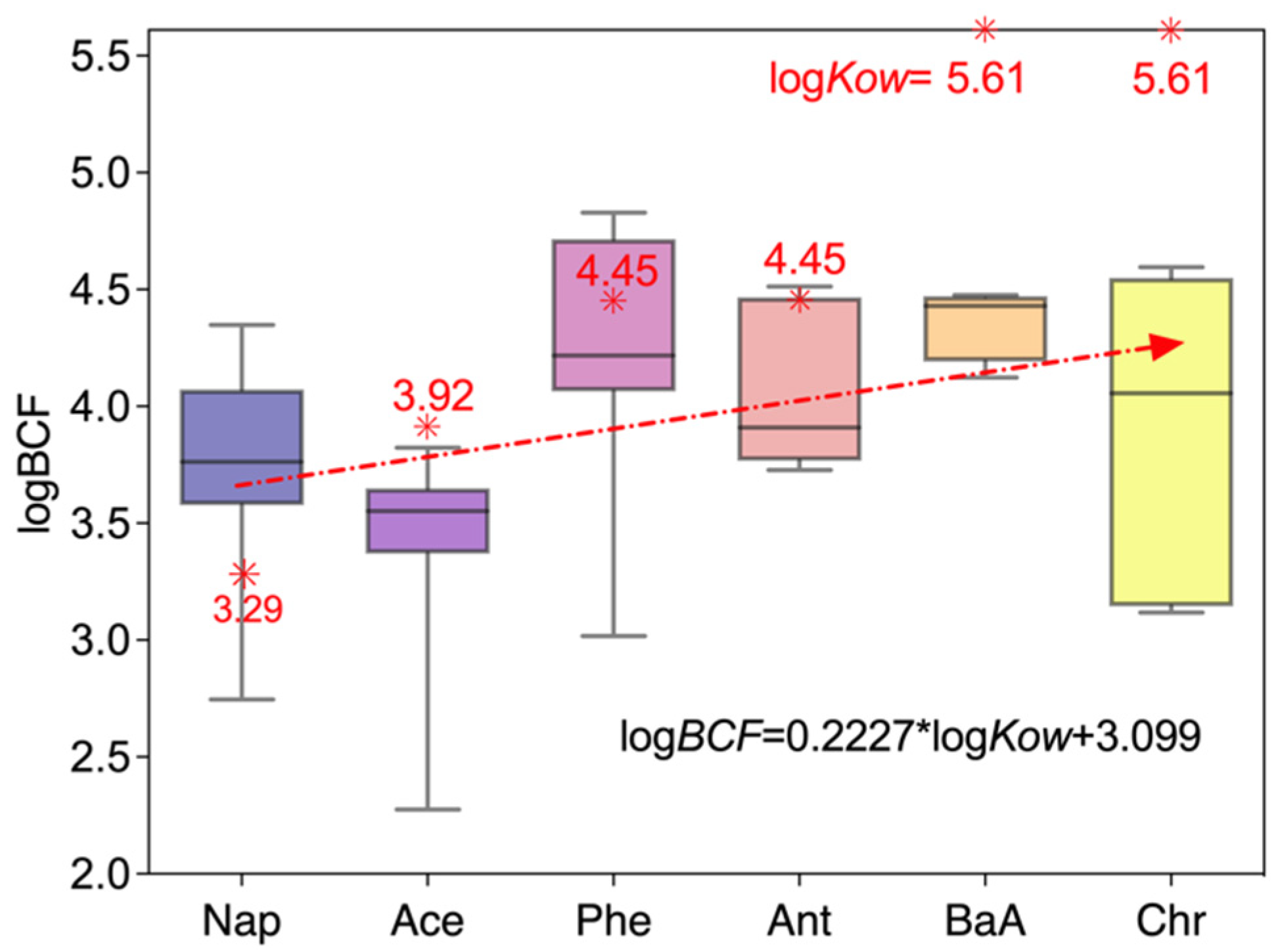
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Zhou, S.; Song, J.; Wu, S. Human health risks of polycyclic aromatic hydrocarbons in the urban soils of Nanjing, China. Sci. Total Environ. 2018, 612, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, Y.; Li, Y.; Sheng, G.; Yan, G. Accumulation and biodegradation of anthracene by Chlorella protothecoides under different trophic conditions. Chin. J. Appl. Ecol. 2002, 13, 145–150. [Google Scholar]
- Hung, W.-N.; Chiou, C.T.; Lin, T.-F. Lipid–water partition coefficients and correlations with uptakes by algae of organic compounds. J. Hazard. Mater. 2014, 279, 197–202. [Google Scholar] [CrossRef]
- Watanabe, K.H.; Lin, H.-I.; Bart, H.L.; Martinat, P.; Means, J.C.; Kunas, M.L.; Grimm, D.A. Bayesian estimation of kinetic rate constants in a food-web model of polycyclic aromatic hydrocarbon bioaccumulation. Ecol. Model. 2005, 181, 229–246. [Google Scholar] [CrossRef]
- Koelmans, A.A. Limited Reversibility of Bioconcentration of Hydrophobic Organic Chemicals in Phytoplankton. Environ. Sci. Technol. 2014, 48, 7341–7348. [Google Scholar] [CrossRef] [PubMed]
- Pathak, B.; Gupta, S.; Verma, R. Biosorption and Biodegradation of Polycyclic Aromatic Hydrocarbons (PAHs) by Microalgae. In Green Adsorbents for Pollutant Removal: Fundamentals and Design; Crini, G., Lichtfouse, E., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 215–247. [Google Scholar] [CrossRef]
- Baghour, M. Algal Degradation of Organic Pollutants. In Handbook of Ecomaterials; Martínez, L.M.T., Kharissova, O.V., Kharisov, B.I., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 565–586. [Google Scholar] [CrossRef]
- Casserly, D.M.; Davis, E.M.; Downs, T.D.; Guthrie, R.K. Sorption of organics by Selenastrum capricornutum. Water Res. 1983, 17, 1591–1594. [Google Scholar] [CrossRef]
- Swackhamer, D.L.; Skoglund, R.S. Bioaccumulation of PCBs by algae: Kinetics versus equilibrium. Environ. Toxicol. Chem. 1993, 12, 831–838. [Google Scholar] [CrossRef]
- Geyer, H.; Viswanathan, R.; Freitag, D.; Korte, F. Relationship between water solubility of organic chemicals and their bioaccumulation by the alga Chlorella. Chemosphere 1981, 10, 1307–1313. [Google Scholar] [CrossRef]
- Mailhot, H. Prediction of Algal Bioaccumulation and Uptake Rate of Nine Organic Compounds by Ten Physicochemical Properties. Environ. Sci. Technol. 1987, 21, 1009–1013. [Google Scholar] [CrossRef]
- Sijm, D.T.H.M.; Middelkoop, J.; Vrisekoop, K. Algal density dependent bioconcentration factors of hydrophobic chemicals. Chemosphere 1995, 31, 4001–4012. [Google Scholar] [CrossRef]
- Fan, C.W.; Reinfelder, J. Phenanthrene accumulation kinetics in marine diatoms. Environ. Sci. Technol. 2003, 37, 3405–3412. [Google Scholar] [CrossRef] [PubMed]
- García de Llasera, M.P.; Olmos-Espejel, J.d.J.; Díaz-Flores, G.; Montaño-Montiel, A. Biodegradation of benzo(a)pyrene by two freshwater microalgae Selenastrum capricornutum and Scenedesmus acutus: A comparative study useful for bioremediation. Environ. Sci. Pollut. Res. 2016, 23, 3365–3375. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Anzion, S.F.M.; Lijklema, L. Dynamics of Organic Micropollutant Biosorption to Cyanobacteria and Detritus. Environ. Sci. Technol. 1995, 29, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Martínez, F.; Jarillo, J.A.; Orús, M.I. Interactions between trichlorfon and three Chlorophyceae. Bull. Environ. Contam. Toxicol. 1991, 46, 599–605. [Google Scholar] [CrossRef]
- Soto, C.; Hellebust, J.A.; Hutchinson, T.C. Effect of naphthaene and aqueous crude oil extracts on the green flagellate Chlamydomonas angulosa II. Photosynthesis and the uptake and release of naphthalene. Can. J. Bot. 1975, 53, 118–126. [Google Scholar] [CrossRef]
- Lei, A.-P.; Hu, Z.-L.; Wong, Y.-S.; Tam, N.F.-Y. Removal of fluoranthene and pyrene by different microalgal species. Bioresour. Technol. 2007, 98, 273–280. [Google Scholar] [CrossRef]
- Kirso, U.; Irha, N. Role of algae in fate of carcinogenic polycyclic aromatic hydrocarbons in the aquatic environment. Ecotoxicol. Environ. Saf. 1998, 41, 83–89. [Google Scholar] [CrossRef]
- Hong, Y.-W.; Yuan, D.-X.; Lin, Q.-M.; Yang, T.-L. Accumulation and biodegradation of phenanthrene and fluoranthene by the algae enriched from a mangrove aquatic ecosystem. Mar. Pollut. Bull. 2008, 56, 1400–1405. [Google Scholar] [CrossRef]
- Chung, M.K.; Tsui, M.T.K.; Cheung, K.C.; Tam, N.F.Y.; Wong, M.H. Removal of aqueous phenanthrene by brown seaweed Sargassum hemiphyllum: Sorption-kinetic and equilibrium studies. Sep. Purif. Technol. 2007, 54, 355–362. [Google Scholar] [CrossRef]
- Chan, S.M.N.; Luan, T.; Wong, M.H.; Tam, N.F.Y. Removal and biodegradation of polycyclic aromatic hydrocarbons by Selenastrum capricornutum. Environ. Toxicol. Chem. 2006, 25, 1772–1779. [Google Scholar] [CrossRef]
- Ke, L.; Luo, L.; Wang, P.; Luan, T.; Tam, N.F.-Y. Effects of metals on biosorption and biodegradation of mixed polycyclic aromatic hydrocarbons by a freshwater green alga Selenastrum capricornutum. Bioresour. Technol. 2010, 101, 6950–6961. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cai, F.; Luan, T.; Lin, L.; Chen, B. Pyrene metabolites by bacterium enhancing cell division of green alga Selenastrum capricornutum. Sci. Total Environ. 2019, 689, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wang, P.; Lin, L.; Luan, T.; Ke, L.; Tam, N.F.Y. Removal and transformation of high molecular weight polycyclic aromatic hydrocarbons in water by live and dead microalgae. Process Biochem. 2014, 49, 1723–1732. [Google Scholar] [CrossRef]
- Warshawsky, D.; Cody, T.; Radike, M.; Reilman, R.; Schumann, B.; LaDow, K.; Schneider, J. Biotransformation of benzo[a]pyrene and other polycyclic aromatic hydrocarbons and heterocyclic analogs by several green algae and other algal species under gold and white light. Chem. Biol. Interact. 1995, 97, 131–148. [Google Scholar] [CrossRef]
- Cerniglia, C.E. Biodegradation of polycyclic aromatic hydrocarbons. Curr. Opin. Biotechnol. 1993, 4, 331–338. [Google Scholar] [CrossRef]
- Semple, K.T.; Cain, R.B.; Schmidt, S. Biodegradation of aromatic compounds by microalgae. FEMS Microbiol. Lett. 1999, 170, 291–300. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiencke, C.; Sandermann, H. Activity of phase I and phase II detoxication enzymes in Antarctic and Arctic macroalgae. Mar. Environ. Res. 1999, 48, 23–36. [Google Scholar] [CrossRef]
- Warshawsky, D.; Keenan, T.H.; Reilman, R.; Cody, T.E.; Radike, M.J. Conjugation of benzo[a]pyrene metabolites by freshwater green alga Selenastrum capricornutum. Chem. Biol. Interact. 1990, 74, 93–105. [Google Scholar] [CrossRef]
- Thies, F.; Backhaus, T.; Bossmann, B.; Grimme, L.H. Xenobiotic Biotransformation in Unicellular Green Algae (Involvement of Cytochrome P450 in the Activation and Selectivity of the Pyridazinone Pro-Herbicide Metflurazon). Plant Physiol. 1996, 112, 361. [Google Scholar] [CrossRef]
- Shubert, L.E. Algae as Ecological Indicators; Shubert, L.E., Ed.; Academic Press Inc.: London, UK, 1984; p. 434. [Google Scholar] [CrossRef]
- Zhao, Z.; Mi, T.; Xia, L.; Yan, W.; Jiang, Y.; Gao, Y. Understanding the patterns and mechanisms of urban water ecosystem degradation: Phytoplankton community structure and water quality in the Qinhuai River, Nanjing City, China. Environ. Sci. Pollut. Res. 2013, 20, 5003–5012. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Zhao, Z.; Xia, L.; Jiang, X.; Gao, Y. Effects of water-saving irrigation on the residues and risk of polycyclic aromatic hydrocarbon in paddy field. Sci. Total Environ. 2018, 618, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, Z.; Qiao, H.; Liu, F. Assessment of eutrophication and water quality in the estuarine area of Lake Wuli, Lake Taihu, China. Sci. Total Environ. 2019, 650, 1392–1402. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.; Ma, P.; Hu, Z.; Zhou, G. Study on the evaluation and treatment of lake eutrophication by means of algae biology. J. Saf. Environ. 2005, 5, 87–91. [Google Scholar]
- Zhang, H.; Chen, R.; Li, F.; Chen, L. Effect of flow rate on environmental variables and phytoplankton dynamics: Results from field enclosures. Chin. J. Oceanol. Limnol. 2015, 33, 430–438. [Google Scholar] [CrossRef]
- Li, F.; Zhang, H.; Zhu, Y.; Xiao, Y.; Chen, L. Effect of flow velocity on phytoplankton biomass and composition in a freshwater lake. Sci. Total Environ. 2013, 447, 64–71. [Google Scholar] [CrossRef]
- Tao, Y.; Yu, J.; Liu, X.; Xue, B.; Wang, S. Factors affecting annual occurrence, bioaccumulation, and biomagnification of polycyclic aromatic hydrocarbons in plankton food webs of subtropical eutrophic lakes. Water Res. 2018, 132, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Yu, J.; Xue, B.; Yao, S.; Wang, S. Precipitation and temperature drive seasonal variation in bioaccumulation of polycyclic aromatic hydrocarbons in the planktonic food webs of a subtropical shallow eutrophic lake in China. Sci. Total Environ. 2017, 583, 447–457. [Google Scholar] [CrossRef]
- Tao, Y. Eutrophication-induced regime shifts reduced sediment burial ability for polycyclic aromatic hydrocarbons: Evidence from Lake Taihu in China. Chemosphere 2021, 271, 129709. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, Y.; Cao, J.; Wu, Z.; Yao, S.; Xue, B. Climate change has weakened the ability of Chinese lakes to bury polycyclic aromatic hydrocarbons. Environ. Pollut. 2019, 255, 113288. [Google Scholar] [CrossRef]
- Kottuparambil, S.; Agusti, S. Cell-by-cell estimation of PAH sorption and subsequent toxicity in marine phytoplankton. Chemosphere 2020, 259, 127487. [Google Scholar] [CrossRef]
- Ding, Q.; Gong, X.; Jin, M.; Yao, X.; Zhang, L.; Zhao, Z. The biological pump effects of phytoplankton on the occurrence and benthic bioaccumulation of hydrophobic organic contaminants (HOCs) in a hypereutrophic lake. Ecotoxicol. Environ. Saf. 2021, 213, 112017. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Fu, H.; Xu, Z.; Zhu, D.; Qu, X. Prediction of hydrophobic organic compound partition to algal organic matter through the growth cycle of Microcystis aeruginosa. Environ. Pollut. 2021, 289, 117827. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-W.; Zeng, E.Y.; Qiu, H.; Yu, K.; Cai, S. Bioconcentration of polybrominated diphenyl ethers and organochlorine pesticides in algae is an important contaminant route to higher trophic levels. Sci. Total Environ. 2017, 579, 1885–1893. [Google Scholar] [CrossRef] [PubMed]
- Szczybelski, A.S.; van den Heuvel-Greve, M.J.; Kampen, T.; Wang, C.; van den Brink, N.W.; Koelmans, A.A. Bioaccumulation of polycyclic aromatic hydrocarbons, polychlorinated biphenyls and hexachlorobenzene by three Arctic benthic species from Kongsfjorden (Svalbard, Norway). Mar. Pollut. Bull. 2016, 112, 65–74. [Google Scholar] [CrossRef]
- Duan, D.; Huang, Y.; Cheng, H.; Ran, Y. Relationship of polycyclic aromatic hydrocarbons with algae-derived organic matter in sediment cores from a subtropical region. J. Geophys. Res. Biogeosci. 2015, 120, 2243–2255. [Google Scholar] [CrossRef] [Green Version]
- She, Z.; Huang, X. Research Methods of Freshwater Plankton; Science Press: Beijing, China, 1991. [Google Scholar]
- Wallberg, P.; Andersson, A. Determination of adsorbed and absorbed polychlorinated biphenyls (PCBs) in seawater microorganisms. Mar. Chem. 1999, 64, 287–299. [Google Scholar] [CrossRef]
- Tao, Y.; Yu, J.; Lei, G.; Xue, B.; Zhang, F.; Yao, S. Indirect influence of eutrophication on air–water exchange fluxes, sinking fluxes, and occurrence of polycyclic aromatic hydrocarbons. Water Res. 2017, 122, 512–525. [Google Scholar] [CrossRef]
- Tao, Y.; Liu, D. Trophic status affects the distribution of polycyclic aromatic hydrocarbons in the water columns, surface sediments, and plankton of twenty Chinese lakes. Environ. Pollut. 2019, 252, 666–674. [Google Scholar] [CrossRef]
- Ben Othman, H.; Lanouguère, É.; Got, P.; Sakka Hlaili, A.; Leboulanger, C. Structural and functional responses of coastal marine phytoplankton communities to PAH mixtures. Chemosphere 2018, 209, 908–919. [Google Scholar] [CrossRef]
- Kottuparambil, S.; Agusti, S. PAHs sensitivity of picophytoplankton populations in the Red Sea. Environ. Pollut. 2018, 239, 607–616. [Google Scholar] [CrossRef]
- Jaiswal, K.K.; Kumar, V.; Vlaskin, M.S.; Nanda, M. Impact of pyrene (polycyclic aromatic hydrocarbons) pollutant on metabolites and lipid induction in microalgae Chlorella sorokiniana (UUIND6) to produce renewable biodiesel. Chemosphere 2021, 285, 131482. [Google Scholar] [CrossRef] [PubMed]
- Stange, K.; Swackhamer, D.L. Factors affecting phytoplankton species-specific differences in accumulation of 40 polychlorinated biphenyls (PCBs). Environ. Toxicol. Chem. 1994, 13, 1849–1860. [Google Scholar] [CrossRef]
- Halling-Sørensen, B.; Nyholm, N.; Kusk, K.O.; Jacobsson, E. Influence of Nitrogen Status on the Bioconcentration of Hydrophobic Organic Compounds to Selenastrum capricornutum. Ecotoxicol. Environ. Saf. 2000, 45, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.C.; Xu, F.L.; Dawson, R.; Chen, S.H.; Tao, S. Adsorption and absorption of polycyclic aromatic hydrocarbons to rice roots. Environ. Pollut. 2007, 148, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Javid, H.; Zhao, Z.; Pang, Y.; Xia, L.; Hussain, I.; Jiang, X. Effects of Different Water Seasons on the Residual Characteristics and Ecological Risk of Polycyclic Aromatic Hydrocarbons in Sediments from Changdang Lake, China. J. Chem. 2016, 2016, 8545816. [Google Scholar]
- Tam, N.F.Y.; Ke, L.; Wang, X.H.; Wong, Y.S. Contamination of polycyclic aromatic hydrocarbons in surface sediments of mangrove swamps. Environ. Pollut. 2001, 114, 255–263. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Genus Name | Eutrophication Level | Algae Abundance | Code | Genus Name | Eutrophication Level | Algae Abundance |
|---|---|---|---|---|---|---|---|
| Cyanophyta | Euglenophyta | ||||||
| Cy1 | Oscillatoria | αm | ++++ | E1 | Euglena | ps, αm, βm | ++ |
| Cy2 | Phormidium | ps | ++++ | E2 | Phacus | βm, os | + |
| Cy3 | Merismopedia | αm | +++ | Chlorophyta | |||
| Bacillariophyta | C1 | Chlorella | ps, αm | ++ | |||
| B1 | Navicula | βαm | + | C2 | Ankistrodesmus | αm, βm | ++ |
| B2 | Melosira | βαm | +++ | C3 | Pandorina | βm, os | ++ |
| B3 | Cyclotella | βαm | + | C4 | Coelastrum | βm, os | + |
| B4 | Stephanodiscus | αm | ++ | C5 | Actinastrum | βm, os | ++ |
| B5 | Tabellaria | os | + | C6 | Oocystis | βm, os | + |
| B6 | Fragilaria | βm | + | C7 | Scenedesmus | αm, βm | ++ |
| B7 | Synedra | βαm | + | C8 | Characium | os, βm | ++ |
| B10 | Gyrosigma | βαm | + | C9 | Closterium | αm, βm | + |
| B11 | Gomphonema | βαm | + | C10 | Pediastrum | βm, os | + |
| B12 | Cymatopleura | αm, βm | + | C11 | Schroederia | ps | + |
| B13 | Stauroneis | os | + | ||||
| Species Matrix vs. Environmental Matrix | Ordination Axes | AX1 | AX2 | AX3 | AX4 | Total Variance |
|---|---|---|---|---|---|---|
| Algae vs. PAHs | Eigenvalues | 0.210 | 0.118 | 0.220 | 0.129 | 1.000 |
| Cumulative percentage variance of fitted response data | 45.70 | 71.30 | 71.80 | 73.60 | ||
| PAHs vs. algae | Eigenvalues | 0.608 | 0.3259 | 0.0521 | 0.0122 | 1.000 |
| Cumulative percentage variance of fitted response data | 60.8 | 93.39 | 98.6 | 99.83 |
| Algae vs. PAHs | PAHs vs. Algae | |||
|---|---|---|---|---|
| Axes | SPEC AX1 | SPEC AX2 | SPEC AX1 | SPEC AX2 |
| Nap | 0.1235 | −0.4714 | −0.7713 | 0.2783 |
| Ace | 0.2914 | −0.5094 | −0.6067 | 0.7643 |
| Phe | −0.1766 | −0.1804 | −0.6816 | −0.7247 |
| Ant | 0.0052 | −0.0169 | 0.9831 | −0.0828 |
| BaA | −0.007 | −0.0548 | 0.9832 | −0.0362 |
| Chr | 0.0011 | −0.0192 | 0.9559 | −0.0792 |
| Cy1 | −0.6637 | −0.0902 | −0.1693 | −0.3668 |
| Cy2 | −0.5175 | −0.1398 | −0.074 | −0.1767 |
| Cy3 | 0.6662 | −0.4762 | −0.1661 | 0.5866 |
| B1 | 0.0056 | 0.0086 | −0.0798 | −0.4991 |
| B2 | −0.134 | −0.7793 | −0.4556 | −0.0083 |
| B3 | −0.4524 | −0.255 | −0.1951 | −0.3489 |
| B4 | 0.187 | 0.0642 | −0.1098 | −0.0116 |
| B5 | 0.3154 | 0.0465 | 0.1103 | −0.1785 |
| B6 | −0.1734 | −0.2423 | −0.0702 | 0.0765 |
| B7 | 0.1868 | 0.0579 | −0.0999 | −0.3387 |
| B10 | −0.2584 | −0.1233 | −0.1745 | −0.3535 |
| B11 | 0.5094 | −0.4376 | −0.1695 | 0.4963 |
| B12 | 0.2422 | −0.126 | 0.3625 | 0.5375 |
| B13 | −0.3717 | 0.0951 | 0.1176 | −0.0955 |
| E1 | 0.4241 | 0.6874 | 0.6355 | −0.2645 |
| E2 | 0.0978 | 0.1568 | 0.4065 | 0.1085 |
| C1 | 0.0897 | 0.2804 | 0.1076 | −0.0033 |
| C2 | 0.1405 | 0.6918 | 0.5239 | −0.2825 |
| C3 | −0.166 | 0.5891 | 0.4964 | −0.2784 |
| C4 | 0.1423 | 0.0922 | −0.3705 | 0.1283 |
| C5 | −0.222 | −0.1693 | −0.1219 | −0.0209 |
| C6 | 0.2121 | 0.5879 | 0.5077 | −0.1591 |
| C7 | 0.3507 | −0.5079 | −0.2782 | 0.2434 |
| C8 | 0.23 | 0.7712 | 0.5496 | −0.2223 |
| C9 | −0.3358 | −0.3092 | −0.1357 | 0.052 |
| C10 | 0.1354 | −0.3871 | −0.2099 | −0.0264 |
| C11 | 0.2808 | −0.1685 | −0.0217 | 0.2809 |
| Matrix | Group | PAHs | Algae type | Sample Sites |
|---|---|---|---|---|
| Algae vs. PAHs | 1 | Nap, Ace | Cy3, B2, B11, B12, C7,C10, C11, B12 | 4A, 5A, 10A |
| 2 | Phe | Cy1, Cy2, B1, B3, B6, B10, B13, C5, C9 | 2A,3A, 8A, 11A, 13A, 14A | |
| 3 | BaA, Chr, Ant | B1, B5, B7, E1, E2, C1, C2, C3, C4, C6, C8, B4 | 1A, 6A, 7A, 9A, 12A, 15A | |
| PAHs vs. algae | 1 | Nap, Ace | Cy3, B2, B6, B11, C4,C7,C9,C10, C11, B4 | 4A, 5A, 10A,12A, 15A |
| 2 | Phe | Cy1, Cy2, B1, B3, B7, B10, C5 | 2A, 3A, 6A, 8A, 11A, 13A,14A | |
| 3 | BaA, Chr, Ant | B1, B5, B12, B13, E1, E2, C1, C2, C3, C6, C8 | 1A, 7A, 9A |
| Group | PAHs | Algae | Algae Entered | Regression Equation | R | F | p |
|---|---|---|---|---|---|---|---|
| Group I | Nap | Abu (1) | E1, | CNap = 40.797 − 0.172 × E1 | 0.533 | 5.174 | 0.041 |
| B12 | CNap = 43.890 − 0.202 × E1 − 6.778 × B12 | 0.724 | 6.609 | 0.012 | |||
| C4 | CNap = 42.579 − 0.209 × E1 − 6.244 × B12 + 5.974 × C4 | 0.832 | 8.250 | 0.004 | |||
| C11 | CNap = 42.645 − 0.212 × E1 − 11.023 × B12 + 5.980 × C4 + 7.447 × C11 | 0.913 | 12.571 | 0.001 | |||
| B11 | CNap = 41.541 − 0.208 × E1 − 10.604 × B12 + 6.272 × C4 + 7.442 × C11 + 4.620 × B11 | 0.946 | 15.374 | 0.000 | |||
| Bio (2) | E1, B12, Cy3, C4, B5 | CNap = 40.068 − 2.123 × E1 − 595.594 × B12 + 751.959 × Cy3 + 13,544.526 × C4 +19.661 × B5 | 0.964 | 23.525 | 0.000 | ||
| Ace | Abu | C4, | CAce = 10.048 + 3.248 × C4 | 0.583 | 6.685 | 0.023 | |
| E1, | CAce = 11.195 + 3.464 × C4 − 0.070 × E1 | 0.809 | 11.346 | 0.002 | |||
| B11 | CAce = 11.195 + 3.634 × C4 − 0.068 × E1 + 3.136 × B11 | 0.920 | 20.232 | 0.000 | |||
| Bio | C4, E1, B11 | CAce = 11.195 + 7268.369 × C4 − 0.685 × E1 + 156.789 × B11 | 0.920 | 20.232 | 0.000 | ||
| Group II | Phe | Abu | B1 | CPhe = 2.607 + 0.179 × B1 | 0.575 | 6.426 | 0.025 |
| Bio | B1, B10, C4, C3 | CPhe = 1.710 + 12.401 × B1 + 41.121 × B10 + 2379.072 × C4 − 36.288 × C3 | 0.913 | 12.472 | 0.001 | ||
| Group III | Ant | Abu | B1 | CAnt = 0.027 + 0.015 × B1 | 0.657 | 9.857 | 0.008 |
| Bio | B1 | CAnt = 0.025 + 0.738 × B1 | 0.647 | 9.345 | 0.009 | ||
| BaA | Abu | B1 | CBaA = 0.005 + 0.013 × B1 | 0.678 | 11.070 | 0.005 | |
| B10 | CBaA = −0.032 + 0.015 × B1 + 0.080 × B10 | 0.815 | 11.860 | 0.001 | |||
| Cy2 | CBaA = −0.036 + 0.015 × B1 + 0.164 × B10 − 0.003 × Cy2 | 0.961 | 44.868 | 0.000 | |||
| C6 | CBaA = −0.025 + 0.016 × B1 + 0.160 × B10 − 0.003 × Cy2 − 0.008 × C6 | 0.979 | 57.138 | 0.000 | |||
| Cy3 | CBaA = −0.015 + 0.015 × B1 + 0.156 × B10 − 0.003 × Cy2 − 0.008 × C6 − 0.001 × Cy3 | 0.987 | 70.048 | 0.000 | |||
| B13 | CBaA = −0.002 + 0.016 × B1 + 0.151 × B10 − 0.002 × Cy2 − 0.008 × C6 − 0.001 × Cy3 − 0.011 × B13 | 0.993 | 93.543 | 0.000 | |||
| B4 | CBaA = 0.015 + 0.016 × B1 + 0.146 × B10 − 0.002 × Cy2 − 0.007 × C6 − 0.001 × Cy3 − 0.014 × B13 − 0.002 × B4 | 0.996 | 132.037 | 0.000 | |||
| Bio | B1, B10, Cy2, C6 | CBaA = −0.028 + 0.783 × B1 + 4.023 × B10 − 3.826 × Cy2 − 0.412 × C6 | 0.978 | 54.740 | 0.000 | ||
| Chr | Abu | B1 | CChr = −0.018 + 0.023 × B1 | 0.862 | 37.670 | 0.000 | |
| B7 | CChr = −0.015 + 0.015 × B1 + 0.007 × B7 | 0.924 | 35.079 | 0.000 | |||
| B3 | CChr = −0.012 − 0.003 × B1 + 0.016 × B7 − 0.034 × B3 | 0.954 | 36.843 | 0.000 | |||
| B7,B3 | CChr = −0.014 + 0.015 × B7 + 0.030 × B3 | 0.953 | 59.420 | 0.000 | |||
| C4 | CChr = −0.028 + 0.015 × B7 + 0.031 × B3 + 0.048 × C4 | 0.970 | 58.397 | 0.000 | |||
| Bio | B1, B7, C1, B5 | CChr = −0.029 + 0.766 × B1 + 0.349 × B7 − 11.653 × C1 − 0.406 × B5 | 0.959 | 28.535 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, L.; Zhao, Z.; Lang, Z.; Qin, Z.; Zhu, Y. Exploring the Cumulative Selectivity of Polycyclic Aromatic Hydrocarbons in Phytoplankton, Water, and Sediment in Typical Urban Water Bodies. Water 2022, 14, 3145. https://doi.org/10.3390/w14193145
Xia L, Zhao Z, Lang Z, Qin Z, Zhu Y. Exploring the Cumulative Selectivity of Polycyclic Aromatic Hydrocarbons in Phytoplankton, Water, and Sediment in Typical Urban Water Bodies. Water. 2022; 14(19):3145. https://doi.org/10.3390/w14193145
Chicago/Turabian StyleXia, Liling, Zhenhua Zhao, Zihan Lang, Zhirui Qin, and Yuelong Zhu. 2022. "Exploring the Cumulative Selectivity of Polycyclic Aromatic Hydrocarbons in Phytoplankton, Water, and Sediment in Typical Urban Water Bodies" Water 14, no. 19: 3145. https://doi.org/10.3390/w14193145
APA StyleXia, L., Zhao, Z., Lang, Z., Qin, Z., & Zhu, Y. (2022). Exploring the Cumulative Selectivity of Polycyclic Aromatic Hydrocarbons in Phytoplankton, Water, and Sediment in Typical Urban Water Bodies. Water, 14(19), 3145. https://doi.org/10.3390/w14193145