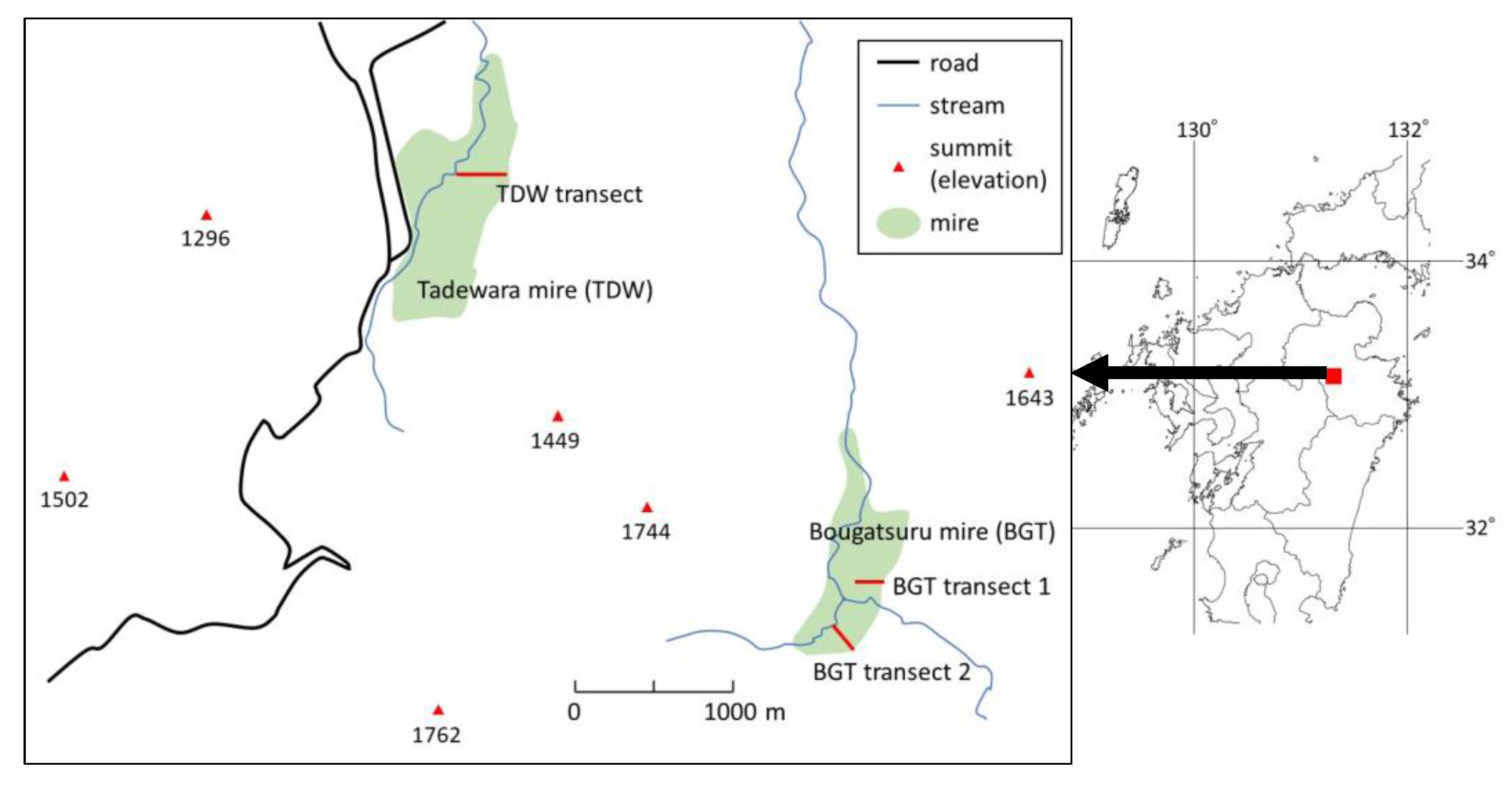
Figure 1.
Map showing the Tadewara mire (TDW) and Bougatsuru mire (BGT) in south-western Japan. TDW transect, BGT transect 1, and BGT transect 2 are investigated transects with a length of 160 m, 70 m, and 90 m, respectively.
Figure 1.
Map showing the Tadewara mire (TDW) and Bougatsuru mire (BGT) in south-western Japan. TDW transect, BGT transect 1, and BGT transect 2 are investigated transects with a length of 160 m, 70 m, and 90 m, respectively.
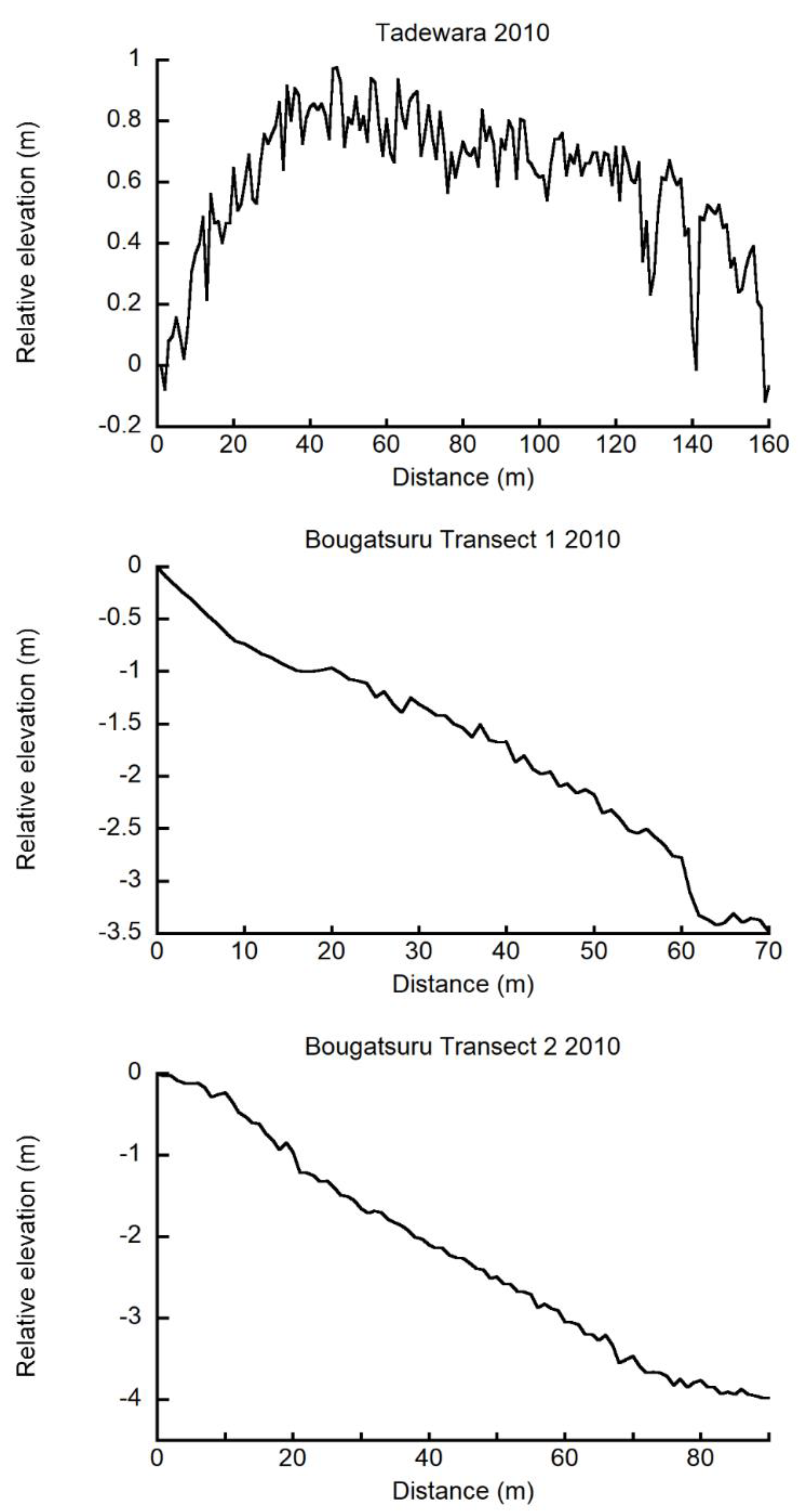
Figure 2.
Elevation of ground surface relative to the origin (0 m site) of Tadewara transect (TDW transect), Bougatsuru transect 1 (BGT transect 1), and Bougatsuru transect 2 (BGT transect 2). Data were collected in July 2010.
Figure 2.
Elevation of ground surface relative to the origin (0 m site) of Tadewara transect (TDW transect), Bougatsuru transect 1 (BGT transect 1), and Bougatsuru transect 2 (BGT transect 2). Data were collected in July 2010.
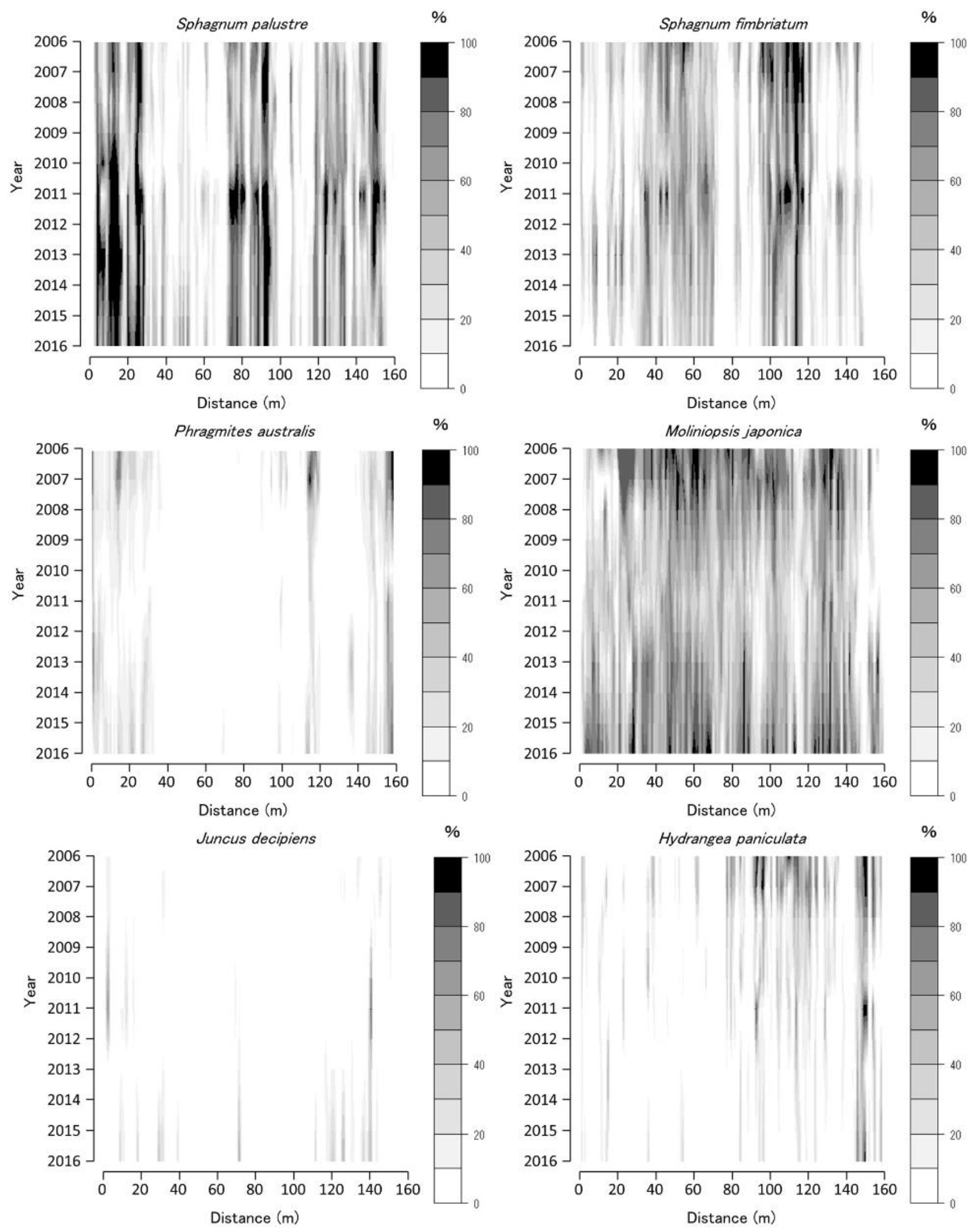
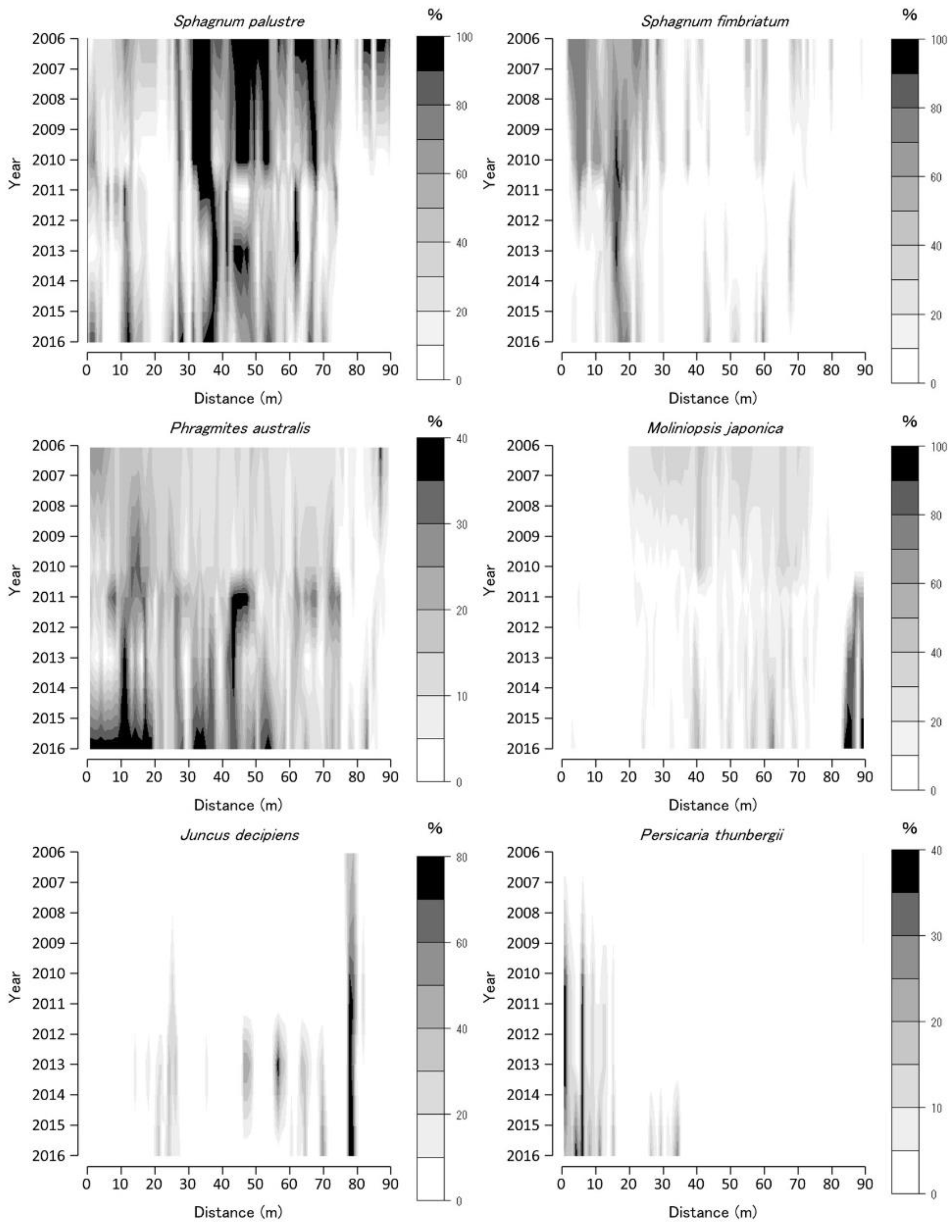
Figure 3.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, Juncus decipiens, and Hydrangea paniculata along the Tadewara transect (TDW transect). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2007, 2008, 2010, 2011, 2013, and 2016.
Figure 3.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, Juncus decipiens, and Hydrangea paniculata along the Tadewara transect (TDW transect). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2007, 2008, 2010, 2011, 2013, and 2016.
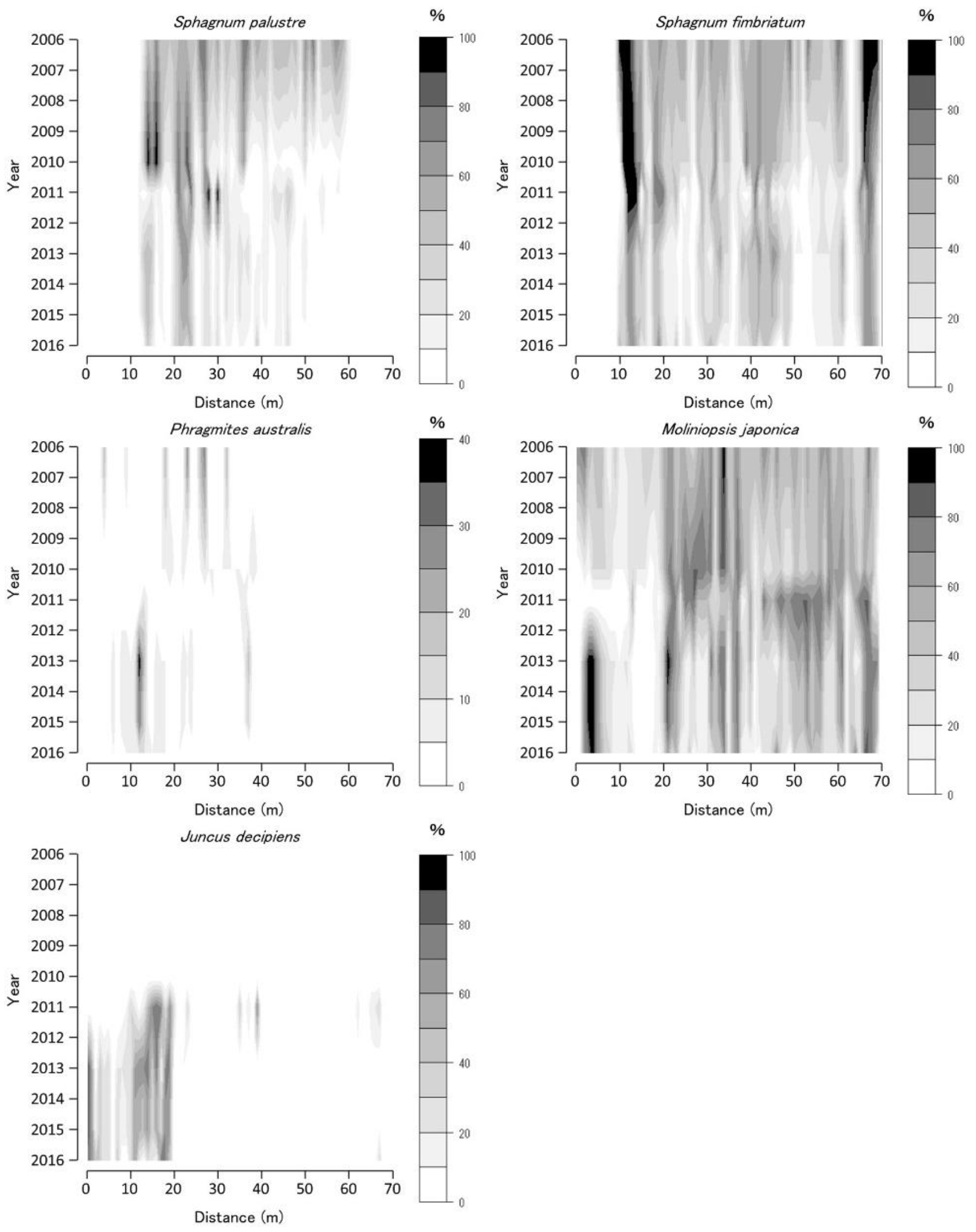
Figure 4.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, and Juncus decipiens along Bougatsuru transect 1 (BGT transect 1). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2010, 2011, 2013, and 2016.
Figure 4.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, and Juncus decipiens along Bougatsuru transect 1 (BGT transect 1). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2010, 2011, 2013, and 2016.
Figure 5.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, Juncus decipiens, and Persicalia thunbergii along Bougatsuru transect 2 (BGT transect 2). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2010, 2011, 2013, and 2016.
Figure 5.
Coverage of Sphagnum palustre, Sphagnum fimbriatum, Phragmites australis, Moliniopsis japonica, Juncus decipiens, and Persicalia thunbergii along Bougatsuru transect 2 (BGT transect 2). Data were collected at every 1 × 1 m2 quadrat placed sequentially on the transect. Countered figures of coverage on the axes of year and position were drawn by interpolating data collected in 2006, 2010, 2011, 2013, and 2016.
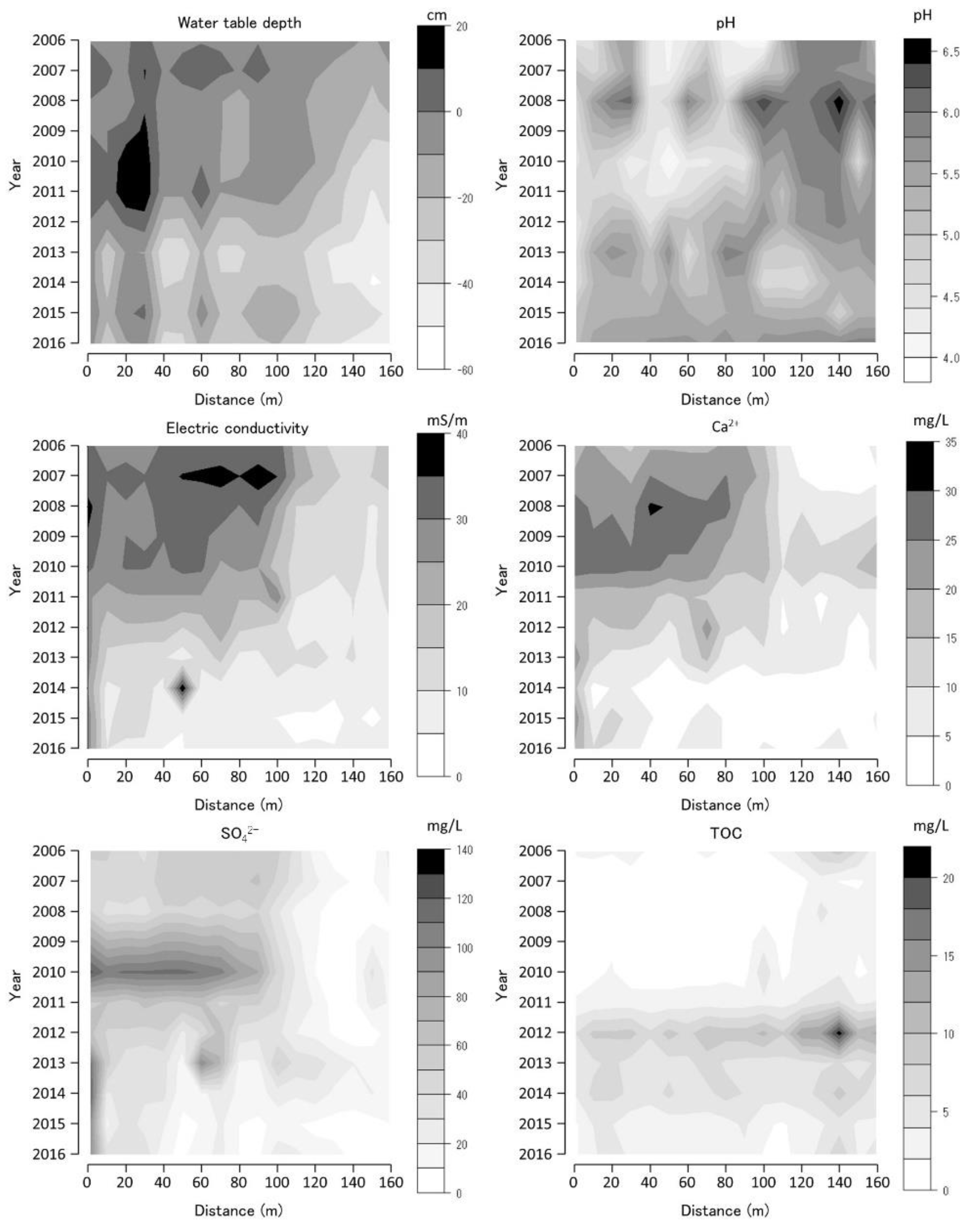
Figure 6.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along the Tadewara transect (TDW transect). Data were collected at 17 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2009).
Figure 6.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along the Tadewara transect (TDW transect). Data were collected at 17 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2009).
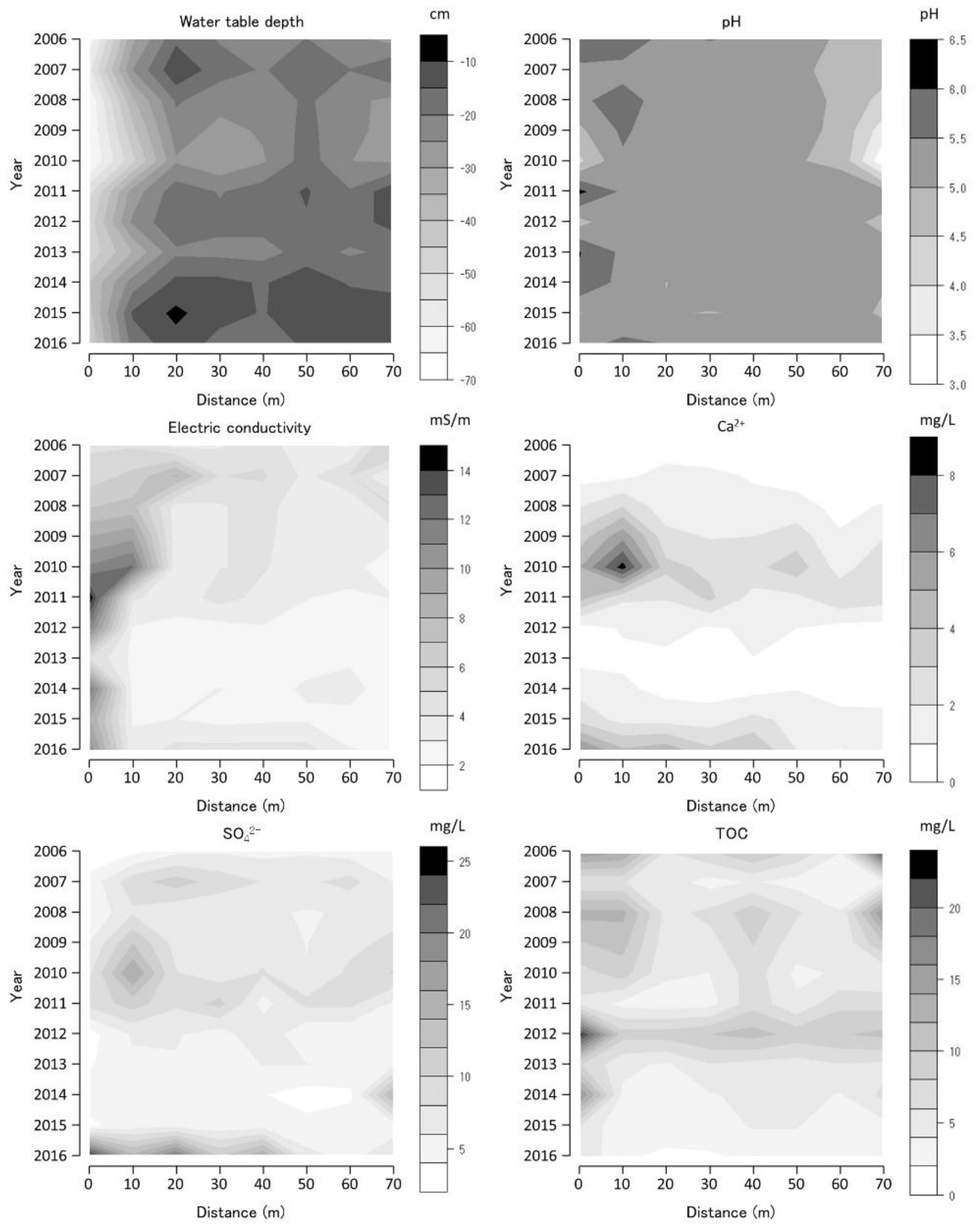
Figure 7.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along Bougatsuru transect 1 (BGT transect 1). Data were collected at 8 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2008 and 2009).
Figure 7.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along Bougatsuru transect 1 (BGT transect 1). Data were collected at 8 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2008 and 2009).
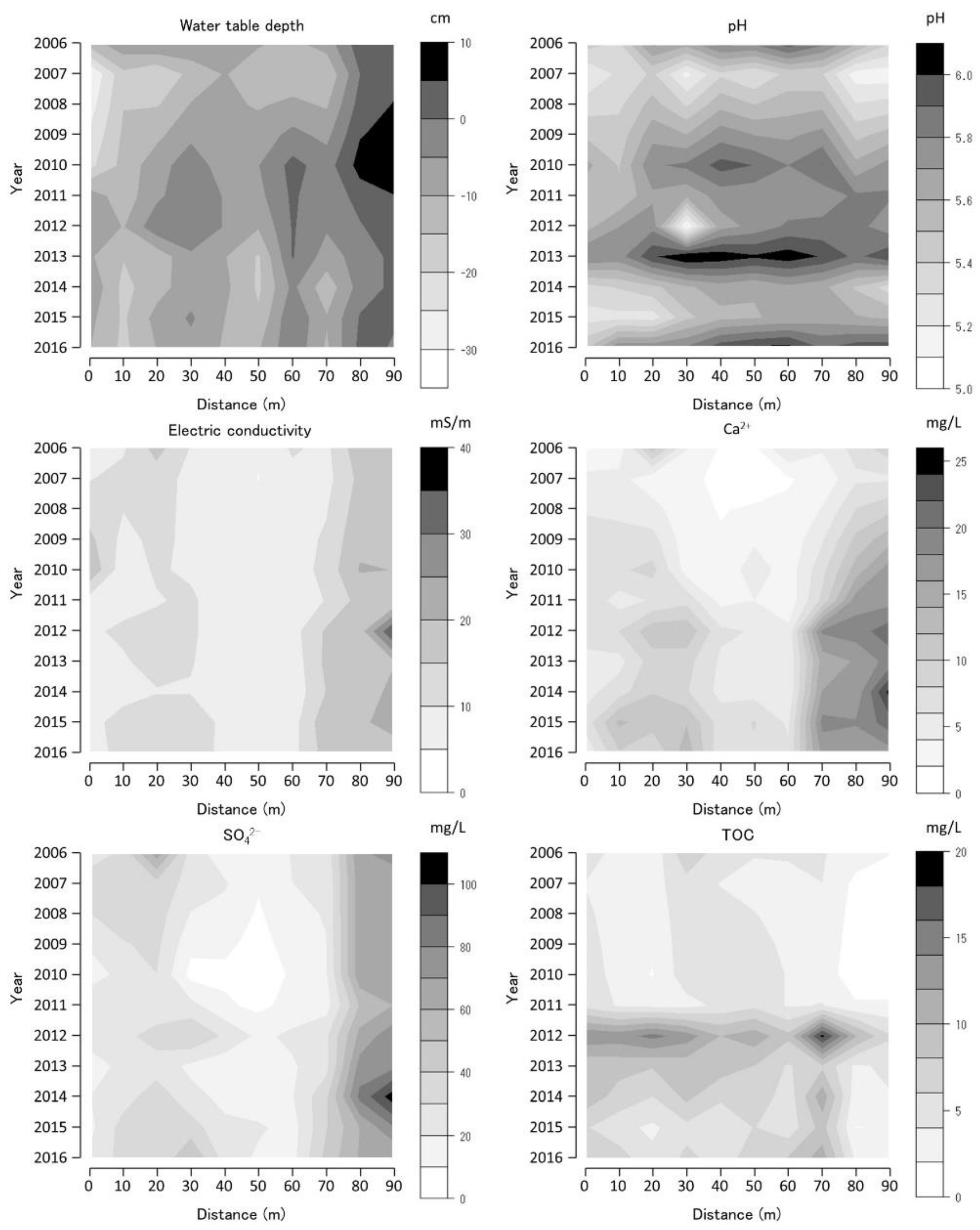
Figure 8.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along Bougatsuru transect 2 (BGT transect 2). Data were collected at 10 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2008 and 2009).
Figure 8.
Water-table depth (WTD), pH, electric conductivity (EC), calcium ion concentration (Ca2+), sulfate ion concentration (SO42−), and total organic carbon (TOC) along Bougatsuru transect 2 (BGT transect 2). Data were collected at 10 sites of water environment monitoring with 10 m intervals along the transect. Countered figures of chemical variables on the axes of year and position were drawn by interpolating missing data (in 2008 and 2009).
Table 1.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along the Tadewara transect (TDW transect). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red-colored cells show the increase in coverage and blue-colored cells show the decrease in coverage from the beginning to the end of each period.
Table 1.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along the Tadewara transect (TDW transect). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red-colored cells show the increase in coverage and blue-colored cells show the decrease in coverage from the beginning to the end of each period.
| Tadewara Mire from 2006 to 2011 |
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 | 110 | 120 | 130 | 140 | 150 | 160 |
| Sphagnumfimbriatum | | | | | | | | * | * | | | | | * | | | |
| Sphagnumpalustre | | | | | | | | | ** | | | | * | ** | * | | |
| Moliniopsisjaponica | | | * | ** | ** | ** | ** | ** | ** | ** | ** | | | * | * | * | |
| Phragmitesaustralis | | | ** | * | | | | | | | * | | * | | | * | |
| Juncusdecipiens | | * | | | | | | | | | | | | | | | |
| Hydrangeapaniculata | | | | | | | | | * | * | * | ** | * | * | | * | |
| Tadewara Mire from 2011 to 2016 |
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 | 110 | 120 | 130 | 140 | 150 | 160 |
| Sphagnumfimbriatum | | | | | * | ** | * | * | * | | | | | | ** | | |
| Sphagnumpalustre | | | * | | | ** | | | | | | | | | | * | |
| Moliniopsisjaponica | * | ** | * | * | * | | ** | | * | * | ** | * | | * | * | * | * |
| Phragmitesaustralis | | | ** | | | | | | | | | | | | | | |
| Juncusdecipiens | | | | | | | | | | | | | * | * | | | |
| Hydrangeapaniculata | * | | | | | | | | | | | * | | | | | |
Table 2.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along Bougatsuru transect 1 (Bougatsuru transect 1). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red-colored cells show the increase in coverage and blue-colored cells show the decrease in coverage from the beginning to the end of each period.
Table 2.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along Bougatsuru transect 1 (Bougatsuru transect 1). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red-colored cells show the increase in coverage and blue-colored cells show the decrease in coverage from the beginning to the end of each period.
| Bougatsuru Mire Transect 1 from 2006 to 2011 |
|---|
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 |
|---|
| Sphagnumfimbriatum | | | | * | | ** | * | |
| Sphagnumpalustre | | | | | ** | ** | * | |
| Moliniopsisjaponica | * | * | * | | | ** | | |
| Phragmitesaustralis | | | | | * | | | |
| Juncusdecipiens | | * | * | | | | | |
| Bougatsuru Mire Transect 1 from 2011 to 2016 |
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 |
| Sphagnumfimbriatum | | | * | | | | | |
| Sphagnumpalustre | | | | * | | * | | |
| Moliniopsisjaponica | | * | | * | * | ** | * | |
| Phragmitesaustralis | | * | * | | | | | |
| Juncusdecipiens | * | | | | | | | |
Table 3.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along the Bougatsuru transect 2 (Bougatsuru transect 2). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red colored cells show the increase in coverage and blue colored cells show decrease in coverage from the beginning to the end of each period.
Table 3.
Significance levels of difference of coverage of abundant species between 2006 and 2011 and between 2011 and 2016 at water environment monitoring sites along the Bougatsuru transect 2 (Bougatsuru transect 2). Five 1 × 1 m2 quadrats of both sides of each water environment monitoring site (one side at both end sites) were assigned to each site. Significance of difference was tested by Wilcoxon test. Red colored cells show the increase in coverage and blue colored cells show decrease in coverage from the beginning to the end of each period.
| Bougatsuru Mire Transect 2 from 2006 to 2011 |
|---|
| Distance(m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 |
|---|
| Sphagnumfimbriatum | | * | | ** | * | | * | * | | |
| Sphagnumpalustre | * | | ** | | | ** | ** | ** | | * |
| Moliniopsisjaponica | | | * | ** | ** | ** | ** | * | | * |
| Phragmitesaustralis | * | * | * | ** | * | | * | ** | * | |
| Juncusdecipiens | | | | * | | | | | | |
| Persicariathunbergii | * | * | | | | | | | | |
| Bougatsuru mire transect 2 from 2011 to 2016 |
| Distance(m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 |
| Sphagnumfimbriatum | | | | | | | * | | | |
| Sphagnumpalustre | | | * | | | ** | | | | |
| Moliniopsisjaponica | | | | | * | | * | | | |
| Phragmitesaustralis | * | * | * | | | | | | | * |
| Juncusdecipiens | | | * | | | | | | | |
| Persicariathunbergii | | | | * | | | | | | |
Table 4.
Significance levels of regression of water environmental variables and elapsed days in Tadewara transect (TDW transect) from 2006 to 2011 and from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
Table 4.
Significance levels of regression of water environmental variables and elapsed days in Tadewara transect (TDW transect) from 2006 to 2011 and from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
| Tadewara Mire from 2006 to 2011 |
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 | 110 | 120 | 130 | 140 | 150 | 160 |
| WTD | ** | | ** | ** | ** | ** | * | ** | * | ** | | * | * | ** | ** | ** | ** |
| pH | | | * | *** | | | | | ** | | ** | | | | | ** | |
| EC | * | ** | ** | | *** | *** | *** | *** | *** | *** | | *** | *** | *** | * | ** | * |
| Cl− | ** | ** | *** | ** | ** | *** | *** | *** | *** | *** | ** | | | ** | ** | *** | *** |
| NO3− | *** | | | | * | | | | | | | | | | | ** | * |
| SO42− | * | | | *** | * | * | | | | | | * | | | | * | |
| NH4+ | | | | | | | | | | | | | ** | | * | | |
| Na+ | * | ** | ** | * | ** | ** | ** | ** | *** | *** | ** | | | *** | *** | *** | *** |
| K+ | | * | ** | * | | ** | *** | | ** | *** | | | * | * | *** | ** | *** |
| Mg2+ | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | ** | | * | ** |
| Ca2+ | | | | | | | * | | *** | *** | * | | * | | *** | ** | |
| TOC | | * | | | ** | ** | ** | ** | ** | *** | | * | | | | | |
| TP | *** | ** | ** | ** | ** | * | ** | * | *** | *** | * | ** | ** | ** | ** | | *** |
| TN | | * | | | | | | | * | * | | | | | | | |
| Tadewara Mire from 2011 to 2016 |
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 | 110 | 120 | 130 | 140 | 150 | 160 |
| WTD | | ** | ** | ** | ** | *** | ** | ** | ** | ** | | ** | ** | * | | | |
| pH | | | *** | *** | *** | *** | *** | ** | ** | | | * | | | | | |
| EC | | *** | *** | *** | *** | | *** | *** | *** | *** | *** | *** | *** | *** | ** | | ** |
| Cl− | | *** | * | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | | *** | *** | *** |
| NO3− | *** | ** | * | ** | | ** | ** | * | ** | ** | * | ** | ** | ** | ** | ** | ** |
| SO42− | ** | ** | | * | *** | *** | | *** | *** | *** | * | | | | | * | ** |
| NH4+ | | | | | | | * | | | | | | | | | * | |
| Na+ | ** | ** | | ** | *** | *** | *** | *** | *** | *** | *** | * | * | | | ** | |
| K+ | * | | | | ** | | | * | | * | | * | * | | | * | |
| Mg2+ | ** | | | ** | *** | *** | ** | *** | *** | *** | ** | | | | | | |
| Ca2+ | | ** | | *** | *** | *** | ** | *** | *** | *** | ** | | * | | | | |
| TOC | ** | | | * | * | | | * | | | *** | *** | *** | ** | | ** | * |
| TP | | ** | * | ** | | * | | | | | | | | | | | |
| TN | | | | | | | | | ** | | * | | | | | | * |
Table 5.
Significance levels of regression of water environmental variables and elapsed days in Bougatsuru transect 1 (BGT transect 1) from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
Table 5.
Significance levels of regression of water environmental variables and elapsed days in Bougatsuru transect 1 (BGT transect 1) from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
| Bougatsuru Mire Transect 1 from 2011 to 2016 |
|---|
| Distance (m) | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 |
|---|
| WTD | | ** | * | | | | ** | |
| pH | *** | | | | | | | |
| EC | | | | | | | | |
| Cl– | | | | | | * | | ** |
| NO3– | | * | * | ** | ** | *** | * | * |
| SO42− | | | | | | | | |
| NH4+ | | * | | | | | | |
| Na+ | | | | | | | | * |
| K+ | | | | | | * | ** | ** |
| Mg2+ | | | | | | | | |
| Ca2+ | | | | | | | | |
| TOC | | *** | *** | *** | *** | ** | ** | ** |
| TP | | | *** | | | | | |
| TN | | | | | | | | |
Table 6.
Significance levels of regression of water environmental variables and elapsed days in Bougatsuru transect 2 (BGT transect 2) from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
Table 6.
Significance levels of regression of water environmental variables and elapsed days in Bougatsuru transect 2 (BGT transect 2) from 2011 to 2016. Red-colored cells mean significant positive slopes and blue-colored cells mean significant negative slopes.
| Bougatsuru Mire Transect 2 from 2011 to 2016 |
|---|
| Distance (m) | Spring | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 |
|---|
| WTD | – | | | | ** | ** | ** | | ** | | *** |
| pH | | * | | | | | | | | | |
| EC | | | * | | * | * | * | ** | *** | | |
| Cl– | | *** | *** | ** | * | * | * | * | * | * | |
| NO3– | | | * | * | | | | ** | * | | |
| SO42− | | | | | ** | ** | *** | | *** | | |
| NH4+ | | * | | | | * | | | * | | |
| Na+ | ** | | * | | ** | | ** | | *** | * | |
| K+ | * | ** | | | | | * | | * | | |
| Mg2+ | *** | | *** | ** | *** | *** | *** | *** | *** | ** | * |
| Ca2+ | | * | * | | ** | ** | ** | *** | *** | | |
| TOC | *** | | | | | | | * | | | |
| TP | | | | | | | | | | | |
| TN | | * | | * | | | | | | | |
Table 7.
Significance levels of correlation between coverage of each abundant species and water environmental variables in transects of the TDW transect, BGT transect 1, and BGT transect 2. Significance levels of Spearman’s’ correlation coefficient were shown. Red-colored cells mean significant positive correlations and blue-colored cells mean significant negative correlations.
Table 7.
Significance levels of correlation between coverage of each abundant species and water environmental variables in transects of the TDW transect, BGT transect 1, and BGT transect 2. Significance levels of Spearman’s’ correlation coefficient were shown. Red-colored cells mean significant positive correlations and blue-colored cells mean significant negative correlations.
| | WTD | pH | EC | Cl− | NO3− | SO42− | NH4+ | Na+ | K+ | Mg2+ | Ca2+ | TOC | TP | TN |
|---|
| Sphagnum fimbriatum | | | | | | | | | | | ** | | | |
| Sphagnum palustre | | | | | | | | | | | | | | |
| Moliniopsis japonica | | * | | | | ** | | | | * | | ** | | ** |
| Phragmites australis | | | | | | | | ** | | ** | | | | |
| Juncus decipiens | | | | | | | | | | | | | | |
| Hydrangea paniculata | | | | | | | | ** | ** | ** | | | | ** |
| Persicaria thunbergii | | | | | | | | | | | | | | |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}