
Trophic Patterns of Freshwater Fish across the Balkan Biodiversity Hotspot

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
3. Results
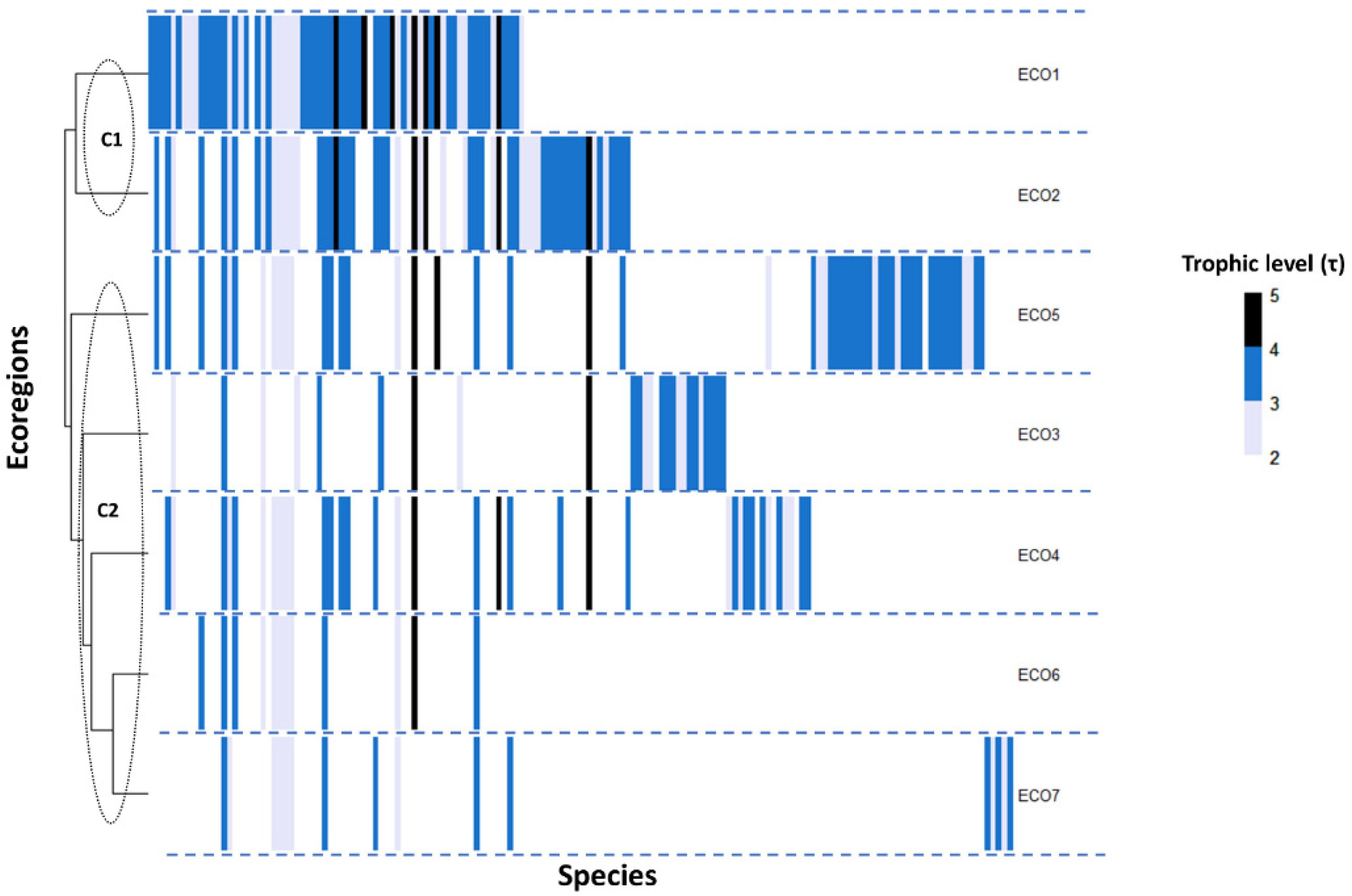
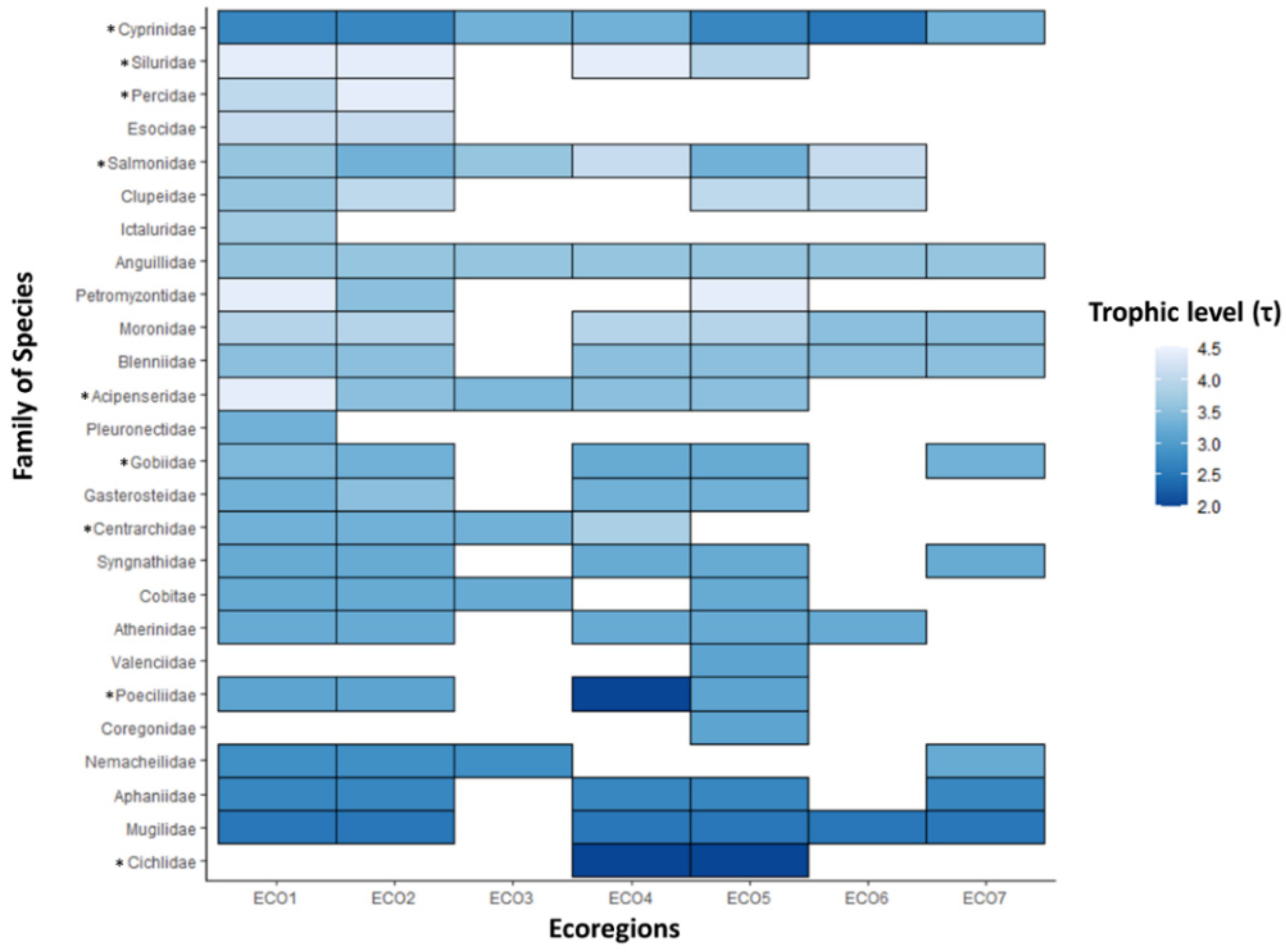
3.1. Trophic Level
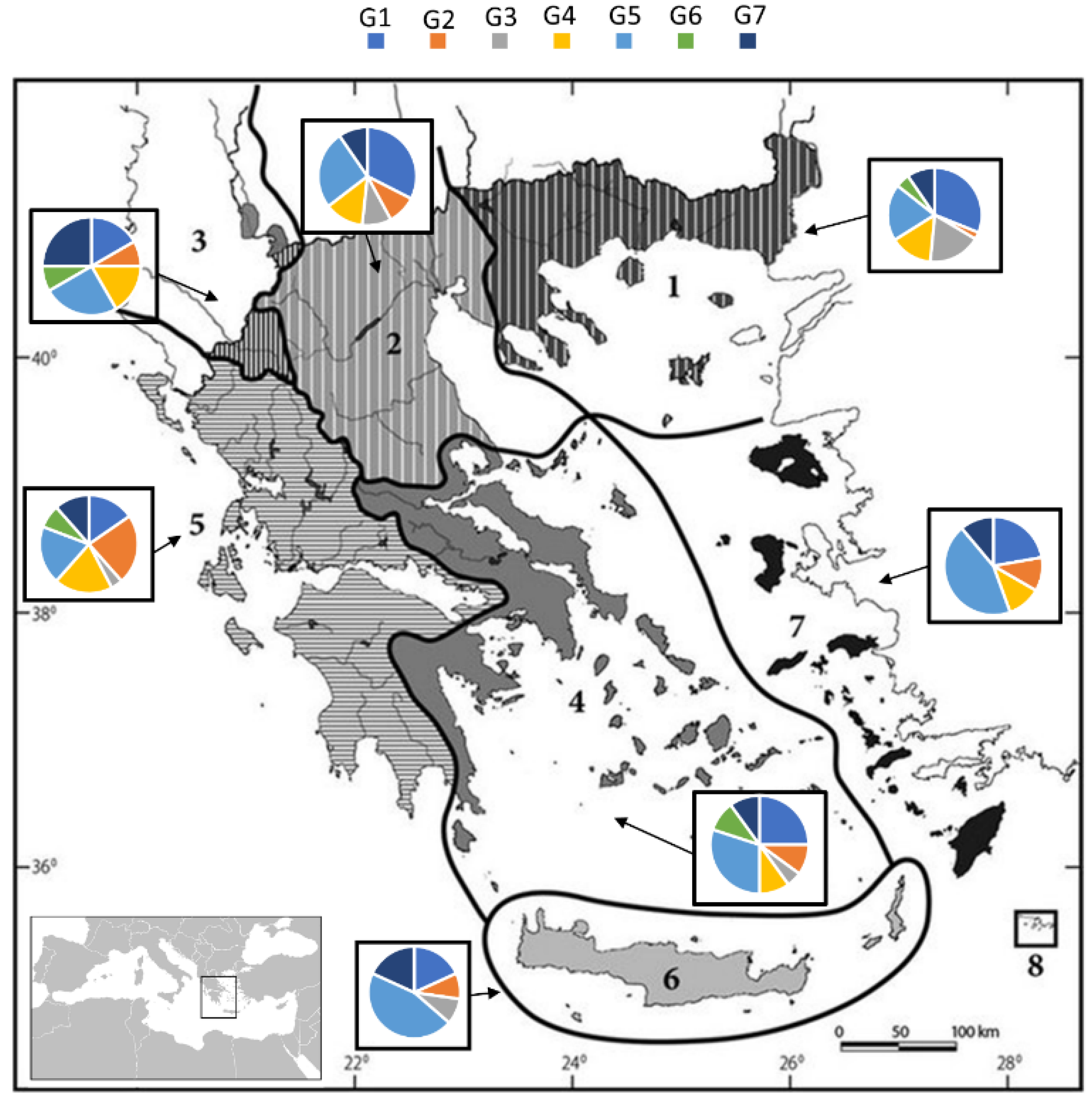
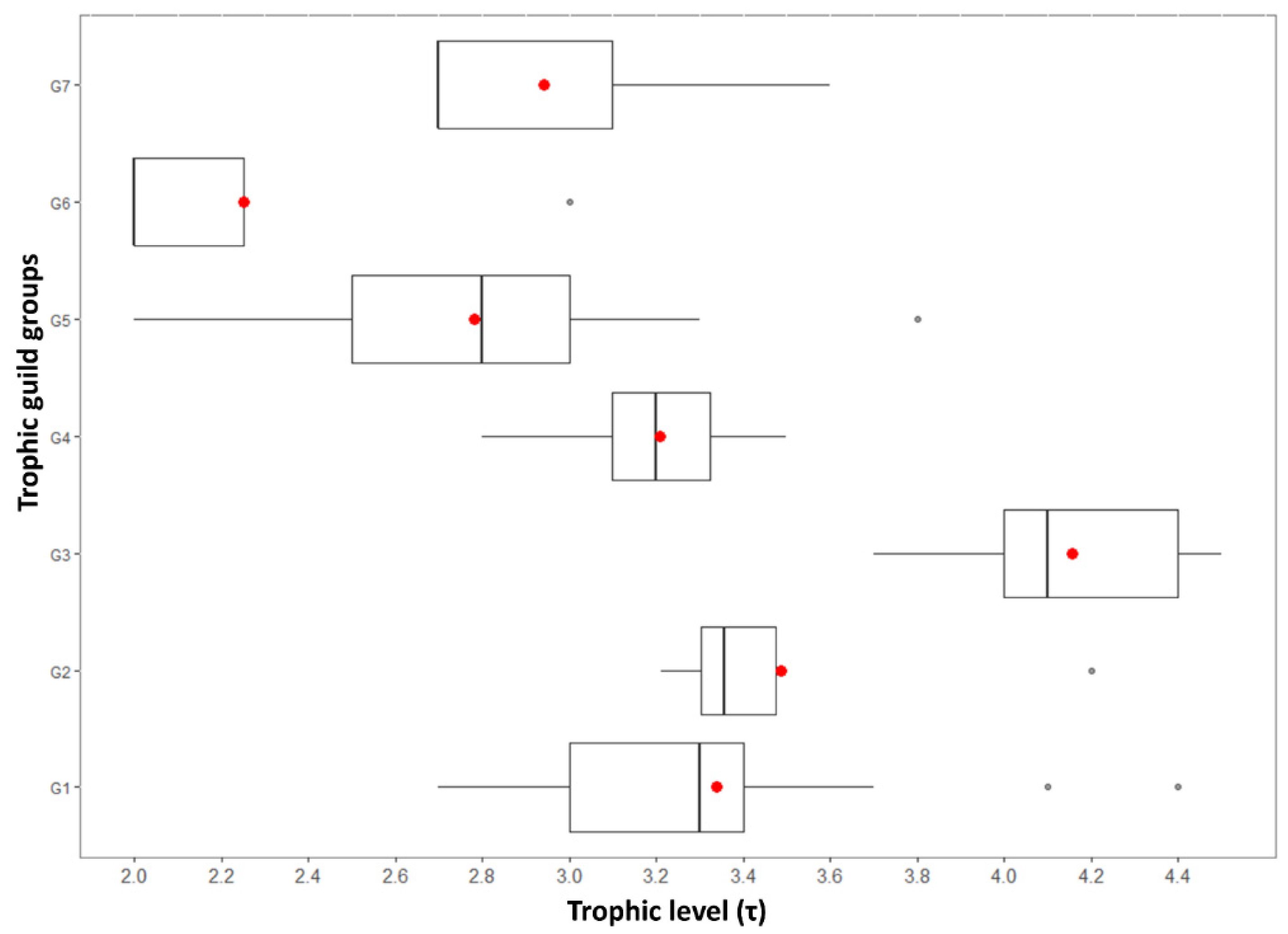
3.2. Trophic Guilds
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2007, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.M.; Cumberlidge, N.; Darwall, W.R.T.; Pollock, C.; Richman, N.I.; Soulsby, A.; Böhm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Angeler, D.G.; Allen, C.R.; Birgé, H.E.; Drakare, S.; McKie, B.G.; Johnson, R.K. Assessing and managing freshwater ecosystems vulnerable to environmental change. Ambio 2014, 43, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winfield, I.J.; Baigún, C.; Balykin, P.A.; Becker, B.; Chen, Y.; Filipe, A.F.; Gerasimov, Y.V.; Godinho, A.L.; Hughes, R.M.; Koehn, J.D.; et al. International Perspectives on the Effects of Climate Change on Inland Fisheries. Fisheries 2016, 41, 399–405. [Google Scholar] [CrossRef]
- WWF. The World’s Forgotten Fishes, 1st ed.; WWF (World Wildlife Fund) International: Gland, Switzerland, 2021; p. 38. Available online: https://wwfint.awsassets.panda.org/downloads/world_s_forgotten_fishes__report_final__1.pdf (accessed on 20 February 2022).
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life, 1st ed.; John Murray: London, UK, 1859; pp. 60–79. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342, 151–164. [Google Scholar] [CrossRef]
- Jeppesen, Ε.; Lauridsen, T.L.; Mitchell, S.F.; Christoffersen, Κ.; Burns, C.W. Trophic structure in the pelagial of 25 shallow New Zealand lakes: Changes along nutrient and fish gradients. J. Plankton Res. 2000, 22, 951–968. [Google Scholar] [CrossRef] [Green Version]
- Gibert, J.P. Temperature directly and indirectly influences food web structure. Sci. Rep. 2019, 9, 5312. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Tammi, J.; Lappalainen, A.; Mannio, J.; Rask, M.; Vuorenmaa, J. Effects of eutrophication on fish and fisheries in Finnish lakes: A survey based on random sampling. Fish. Manag. Ecol. 2001, 6, 173–186. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Møller, P.H.; Sandby, K. Changes in nitrogen retention in shallow eutrophic lakes following a decline in density of cyprinids. Arch. Für Hydrobiol. 1998, 142, 129–151. [Google Scholar] [CrossRef]
- Olin, M.; Rask, M.; Ruuhljärvi, J.; Kurkilahti, M.; Ala-Opas, P.; Ylönen, O. Fish community structure in mesotrophic and eutrophic lakes of southern Finland: The relative abundances of percids and cyprinids along a trophic gradient. J. Fish Biol. 2002, 60, 593–612. [Google Scholar] [CrossRef]
- Minns, C.K.; Kelso, J.R.; Randall, R.G. Detecting the response of fish to habitat alterations in freshwater ecosystems. Can. J. Fish. Aquat. Sci. 1996, 53, 403–414. [Google Scholar] [CrossRef]
- Jones, J.I.; Sayer, C.D. Does the fish-invertebrate-periphyton cascade precipitate plant loss in shallow lakes? Ecology 2003, 84, 2155–2167. [Google Scholar] [CrossRef]
- Terborgh, J.; Estes, J. Trophic Cascades: Predators, Prey, and The Changing Dynamics of Nature; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- de Mutsert, K.; Cowan, J.H., Jr.; Walters, C.J. Using Ecopath with Ecosim to Explore Nekton Community Response to Freshwater Diversion into a Louisiana Estuary. Mar. Coast. Fish. 2012, 4, 104–116. [Google Scholar] [CrossRef]
- Moutopoulos, D.K.; Stoumboudi, M.T.; Ramfos, A.; Tsagarakis, K.; Gritzalis, K.C.; Petriki, O.; Patsia, A.; Barbieri, R.; Machias, A.; Stergiou, K.I.; et al. Food web modelling on the structure and functioning of a Mediterranean lentic system. Hydrobiologia 2018, 822, 259–283. [Google Scholar] [CrossRef]
- Aarts, B.G.; Nienhuis, P.H. Fish zonations and guilds as the basis for assessment of ecological integrity of large rivers. Hydrobiologia 2003, 500, 157–178. [Google Scholar] [CrossRef]
- Noble, R.A.A.; Cowx, I.G.; Goffaux, D.; Kestemont, P. Assessing the health of European rivers using functional ecological guilds of fish communities: Standardising species classification and approaches to metric selection. Fish. Manag. Ecol. 2007, 14, 381–392. [Google Scholar] [CrossRef]
- Logez, M.; Bady, P.; Melcher, A.; Pont, D. A continental-scale analysis of fish assemblage functional structure in European rivers. Ecography 2013, 36, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Darwall, W.; Carrizo, S.; Numa, C.; Barrios, V.; Freyhof, J.; Smith, K. Freshwater Key Biodiversity Areas in the Mediterranean Basin Hotspot: Informing Species Conservation and Development Planning in Freshwater Ecosystems; IUCN (International Union for Conservation of Nature): Gland, Switzerland, 2014; p. 2. [Google Scholar]
- Leonardos, I. The feeding ecology of Aphanius fasciatus (Valenciennes, 1821) in the lagoonal system of Messolongi (western Greece). Sci. Mar. 2008, 72, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Kalogianni, E.; Giakoumi, S.; Andriopoulou, A.; Chatzinikolaou, Y. Feeding ecology of the critically endangered Valencia letourneuxi (Valenciidae). Aquat. Ecol. 2010, 44, 289–299. [Google Scholar] [CrossRef]
- Gkenas, C.; Oikonomou, A.; Economou, A.; Kiosse, F.; Leonardos, I. Life history pattern and feeding habits of the invasive mosquitofish, Gambusia holbrooki, in Lake Pamvotis (NW Greece). J. Biol. Res. 2012, 17, 121–136. [Google Scholar]
- Bobori, D.; Salvarina, I.; Michaloudi, E. Fish dietary patterns in the eutrophic Lake Volvi (East Mediterranean). J. Biol. Res. 2013, 19, 139–149. [Google Scholar]
- Oikonomou, A.; Leprieur, F.; Leonardos, I.D. Ecomorphological diversity of freshwater fishes as a tool for conservation priority setting: A case study from a Balkan hotspot. Environ. Biol. Fishes 2018, 101, 1121–1136. [Google Scholar] [CrossRef]
- Yoğurtçuoğlu, B.; Ekmekçi, F.G.; Karachle, P.K. A review and assessment of fish trophic levels in a large reservoir of central Anatolia, Turkey. Mar. Freshw. Res. 2020, 72, 311–320. [Google Scholar] [CrossRef]
- Bănărescu, P.M. Distribution pattern of the aquatic fauna of the Balkan Peninsula. In Balkan Biodiversity, Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2004; pp. 203–217. [Google Scholar]
- Economidis, P.S.; Bănărescu, P.M. The Distribution and Origins of Freshwater Fishes in the Balkan Peninsula, Especially in Greece. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1991, 76, 257–284. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. Fishbase. Available online: www.fishbase.org (accessed on 1 September 2021).
- Barbieri, R.; Zogaris, S.; Kalogianni, E.; Stoumboudi, M.T.; Chatzinikolaou, Y.; Giakoumi, S.; Kapakos, Y.; Kommatas, D.; Koutsikos, N.; Tachos, V.; et al. Freshwater Fishes and Lampreys of Greece: An Annotated Checklist; Hellenic Centre for Marine Research: Athens, Greece, 2015. Available online: https://imbriw.hcmr.gr/wp-content/uploads/2015/05/FINAL_MARCH_2015_FreshwaterFishlist-1.pdf (accessed on 1 September 2021).
- Koutsikos, N.; Zogaris, S.; Vardakas, L.; Kalantzi, O.I.; Dimitriou, E.; Economou, A.N. Tracking non-indigenous fishes in lotic ecosystems: Invasive patterns at different spatial scales in Greece. Sci. Total Environ. 2019, 659, 389–400. [Google Scholar] [CrossRef]
- Zogaris, S.; Economou, A.N. The Biogeographic Characteristics of the River Basins of Greece. In The Rivers of Greece; Skoulikidis, N., Dimitriou, E., Karaouzas, I., Eds.; Springer: Berlin, Germany, 2017; pp. 53–95. [Google Scholar] [CrossRef]
- Pauly, D.; Froese, R.; Sa-a, P.; Palomares, M.L.; Christensen, V.; Rius, J. Trophlab Manual; ICLARM: Manila, Philippines, 2000. [Google Scholar]
- Gascuel, D.; Guénette, S.; Pauly, D. The trophic-level-based ecosystem modelling approach: Theoretical overview and practical uses. ICES J. Mar. Sci. 2011, 68, 1403–1416. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.R.; Agostinho, C.S.; de Oliveira, R.J.; Marques, E.E. Trophic guilds of fishes in sandbank habitats of a Neotropical river. Neotrop. Ichthyol. 2007, 5, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.H.; Vrijenhoek, R.C. Detrended Correspondence Analysis of Dietary Data. Trans. Am. Fish. Soc. 1998, 117, 29–36. [Google Scholar] [CrossRef]
- Gauch, H.G. Multivariate Analysis in Community Ecology; University Press: Cambridge, UK, 1982; pp. 211–241. [Google Scholar]
- Friedlander, A.M.; Parrish, J.D. Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. J. Exp. Mar. Biol. Ecol. 1998, 224, 1–30. [Google Scholar] [CrossRef]
- Holland, S.M. Detrended Correspondence Analysis (DCA); Department of Geology, University of Georgia: Athens, GA, USA, 2008; Available online: https://strata.uga.edu/software/pdf/dcaTutorial.pdf (accessed on 1 September 2021).
- Kaufman, L.; Rousseeuw, P.J. Finding Groups in Data: An Introduction to Cluster Analysis. In Wiley Series in Probability and Statistics; Barnett, V., Bradley, R.A., Hunter, J.S., Kadane, J.B., Kendall, D.G., Smith, F.M., Stigler, S.M., Teugels, J., Watson, G.S., Eds.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 1990; p. 87. [Google Scholar] [CrossRef]
- Tibshirani, R.; Walther, G.; Hastie, T. Estimating the number of clusters in a data set via the gap statistic. J. R. Stat. Soc. 2001, 63, 411–423. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 September 2021).
- Economou, A.N.; Giakoumi, S.; Vardakas, L.; Barbieri, R.; Stoumboudi, M. Τh.; Zogaris, S. The freshwater ichthyofauna of Greece—An update based on a hydrographic basin survey. Mediterr. Mar. Sci. 2007, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Shumka, S.; Meulenbroek, P.; Schiemer, F.; Sanda, R. Fishes of the River Vjosa—An annotated Checklist. Acta ZooBot Austria 2018, 155, 163–176. [Google Scholar]
- Marić, D. The Skadar/Shkodra Lake Environment. In The Handbook of Environmental Chemistry; Pešić, V., Karaman, G., Kostianoy, A.G., Eds.; Springer International Publishing: New York City, NY, USA, 2018; pp. 363–381. [Google Scholar] [CrossRef]
- Persson, L.; Diehl, S.; Johansson, L.; Andersson, G.; Hamrin, S.F. Shifts in fish communities along the productivity gradient of temperate lakes-patterns and the importance of size-structured interactions. J. Fish Biol. 1991, 38, 281–293. [Google Scholar] [CrossRef]
- Root, R.B. The niche exploitation pattern of the blue-grey gnatcatcher. Ecol. Monogr. 1967, 37, 317–350. [Google Scholar] [CrossRef]
- Specziár, A.; Rezsu, E.T. Feeding guilds and food resource partitioning in a lake fish assemblage: An ontogenetic approach. J. Fish Biol. 2009, 75, 247–267. [Google Scholar] [CrossRef]
- Jacobsen, L.; Perrow, M.R.; Landkildehus, F.; Hjørne, M.; Lauridsen, T.L.; Berg, S. Interactions between piscivores, zooplanktivores and zooplankton in submerged macrophytes: Preliminary observations from enclosure and pond experiments. Hydrobiologia 1997, 342/343, 197–205. [Google Scholar] [CrossRef]
- Tátrai, I.; Mátyás, K.; Korponai, J.; Paulovits, G.; Pekár, F. Management of fish communities and its impacts on the lower trophic levels in shallow ecosystems in Hungary. Hydrobiologia 2003, 506, 489–496. [Google Scholar] [CrossRef]
- Blanco, S.; Romo, S.; Villena, M.J.; Martínez, S. Fish communities and food web interactions in some shallow Mediterranean lakes. Hydrobiologia 2003, 506, 473–480. [Google Scholar] [CrossRef]
- de Mérona, B.; Vigouroux, R. Diet changes in fish species from a large reservoir in South America and their impact on the trophic structure of fish assemblages (Petit-Saut Dam, French Guiana). Ann. Limnol. Int. J. Limnol. 2006, 42, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Delariva, R.L.; Hahn, N.S.; Kashiwaqui, E.A.L. Diet and trophic structure of the fish fauna in a subtropical ecosystem: Impoundment effects. Neotrop. Ichthyol. 2013, 11, 891–904. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, K.; Pedersen, J.; Christoffersen, K.; Jensen, F. Ecological consequences of food partitioning for the fish population structure in a eutrophic lake. Hydrobiologia 1994, 291, 35–45. [Google Scholar] [CrossRef]
- Tolonen, K.T.; Karjalainen, J.; Staff, S.; Leppä, M. Individual and population-level food consumption by cyprinids and percids in a mesotrophic lake. Ecol. Freshw. Fish 2000, 9, 153–162. [Google Scholar] [CrossRef]
- Rakaj, N.; Flloko, A. Conservation status of freshwater fish of Albania. Biol. Conserv. 1995, 72, 195–199. [Google Scholar] [CrossRef]
- Oikonomou, A.; Leprieur, F.; Leonardos, I.D. Biogeography of freshwater fishes of the Balkan Peninsula. Hydrobiologia 2014, 738, 205–220. [Google Scholar] [CrossRef]
- Albrecht, C.; Wolff, C.; Glöer, P.; Wilke, T. Concurrent evolution of ancient sister lakes and sister species: The freshwater gastropod genus Radix in lakes Ohrid and Prespa. Hydrobiologia 2008, 615, 157–167. [Google Scholar] [CrossRef]
- Zogaris, S.; Economou, A.N.; Dimopoulos, P. Ecoregions in the Southern Balkans: Should Their Boundaries Be Revised? Environ. Manag. 2009, 43, 682–697. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D. Functional Organization of Stream Fish Assemblages in Relation to Hydrological Variability. Ecology 1995, 76, 606–627. [Google Scholar] [CrossRef]
- Wilson, S.K.; Bellwood, D.R.; Choat, J.H.; Furnas, M.J. Detritus in the epilithic algal matrix and its use by coral reef fishes. Oceanogr. Mar. Biol. 2003, 41, 279–309. [Google Scholar]
- Amarasinghe, U.S.; Welcomme, R.L. An Analysis of Fish Species Richness in Natural Lakes. Environ. Biol. Fishes 2002, 65, 327–339. [Google Scholar] [CrossRef]
- Oikonomou, A.; Stefanidis, K. α- and β-Diversity Patterns of Macrophytes and Freshwater Fishes are Driven by Different Factors and Processes in Lakes of the Unexplored Southern Balkan Biodiversity Hotspot. Water 2020, 12, 1984. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Michaloudi, E.; Sommer, U. Modifying the PEG model for Mediterranean lakes—No biological winter and strong fish predation. Freshw. Biol. 2014, 59, 1136–1144. [Google Scholar] [CrossRef] [Green Version]
- Perrow, M.R.; Jowit, A.J.D.; Stansfield, J.H.; Phillips, G.L. The practical importance of the interactions between fish, zooplankton and macrophytes in shallow lake restoration. Hydrobiologia 1999, 395, 199–210. [Google Scholar] [CrossRef]
- Meerhoff, M.; Clemente, J.M.; De Mello, F.T.; Iglesias, C.; Pedersen, A.R.; Jeppesen, E. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes? Glob. Chang. Biol. 2007, 13, 1888–1897. [Google Scholar] [CrossRef]
- Mozsár, A.; Sály, P.; Antal, L.; Nagy, S.A.; Boros, G. Organismal stoichiometry at the temporal scale: Seasonal variability shapes interspecific differences in fish. Freshw. Biol. 2018, 64, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Mozsár, A.; Boros, G.; Sály, P.; Antal, L.; Nagy, S.A. Relationship between Fulton’s condition factor and proximate body composition in three freshwater fish species. J. Appl. Ichthyol. 2014, 31, 315–320. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Ecoregion | τmean | N |
|---|---|---|---|
| 1 | Thrace | 3.24 | 67 |
| 2 | Macedonia-Thessaly | 3.23 | 58 |
| 3 | Southeastern Adriatic | 3.15 | 26 |
| 4 | Western Aegean | 3.1 | 38 |
| 5 | Ionian | 3.15 | 57 |
| 6 | Crete | 3.12 | 12 |
| 7 | Eastern Aegean | 3.01 | 17 |
| Number of Optimal Clusters () | Method | ||
|---|---|---|---|
| Average Silhouette Width | Gap Statistic (GS) | Increase Rate of GS | |
| 1min | 0 | 0.141 | 20.9% |
| 2 | 0.192 | 0.178 | 34.8% |
| 3 | 0.289 | 0.273 | 22.1% |
| 4 | 0.334 | 0.350 | 6.3% |
| 5 | 0.339 | 0.374 | 14.4% |
| 6 | 0.390 | 0.436 | 11.8% |
| 7 | 0.403 | 0.495 | −2.4% |
| 8 | 0.340 | 0.483 | 4.3% |
| 9 | 0.352 | 0.505 | 4.9% |
| 10max | 0.351 | 0.531 | |
| Trophic Guild | Algae | Detritus | Aquatic Insects | Terrestrial Insects | Micro-Crustaceans | Other Invertebrates | Nekton | Plant Matter |
|---|---|---|---|---|---|---|---|---|
| G1 | 1.09 | 5.45 | 12.59 | 2.71 | 57.10 * | 17.71 | 0.78 | 2.54 |
| G2 | 0.00 | 0.00 | 15.48 | 7.62 | 24.29 * | 5.23 | 36.70 * | 10.65 |
| G3 | 0.00 | 0.00 | 1.25 | 3.17 | 0.18 | 0.75 | 94.06 * | 0.02 |
| G4 | 2.86 | 0.00 | 65.09 * | 0.00 | 7.34 | 22.71 | 0.66 | 1.30 |
| G5 | 9.09 | 35.15 * | 0.30 | 0.70 | 38.63 * | 5.18 | 0.00 | 10.10 |
| G6 | 79.15 * | 20.85 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| G7 | 0.22 | 29.85 * | 3.02 | 27.10 * | 15.76 | 1.19 | 3.17 | 19.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vagenas, G.; Oikonomou, A.; Karachle, P.K.; Petriki, O.; Stoumboudi, M.T. Trophic Patterns of Freshwater Fish across the Balkan Biodiversity Hotspot. Water 2022, 14, 1112. https://doi.org/10.3390/w14071112
Vagenas G, Oikonomou A, Karachle PK, Petriki O, Stoumboudi MT. Trophic Patterns of Freshwater Fish across the Balkan Biodiversity Hotspot. Water. 2022; 14(7):1112. https://doi.org/10.3390/w14071112
Chicago/Turabian StyleVagenas, Georgios, Anthi Oikonomou, Paraskevi K. Karachle, Olga Petriki, and Maria Th. Stoumboudi. 2022. "Trophic Patterns of Freshwater Fish across the Balkan Biodiversity Hotspot" Water 14, no. 7: 1112. https://doi.org/10.3390/w14071112
APA StyleVagenas, G., Oikonomou, A., Karachle, P. K., Petriki, O., & Stoumboudi, M. T. (2022). Trophic Patterns of Freshwater Fish across the Balkan Biodiversity Hotspot. Water, 14(7), 1112. https://doi.org/10.3390/w14071112