
The Importance of Groundwater Quality and Other Habitat Parameters for Effective Active Protection of an Endangered Plant Species in Eastern Poland

 ,
,  , , and
, , and
Abstract
:1. Introduction
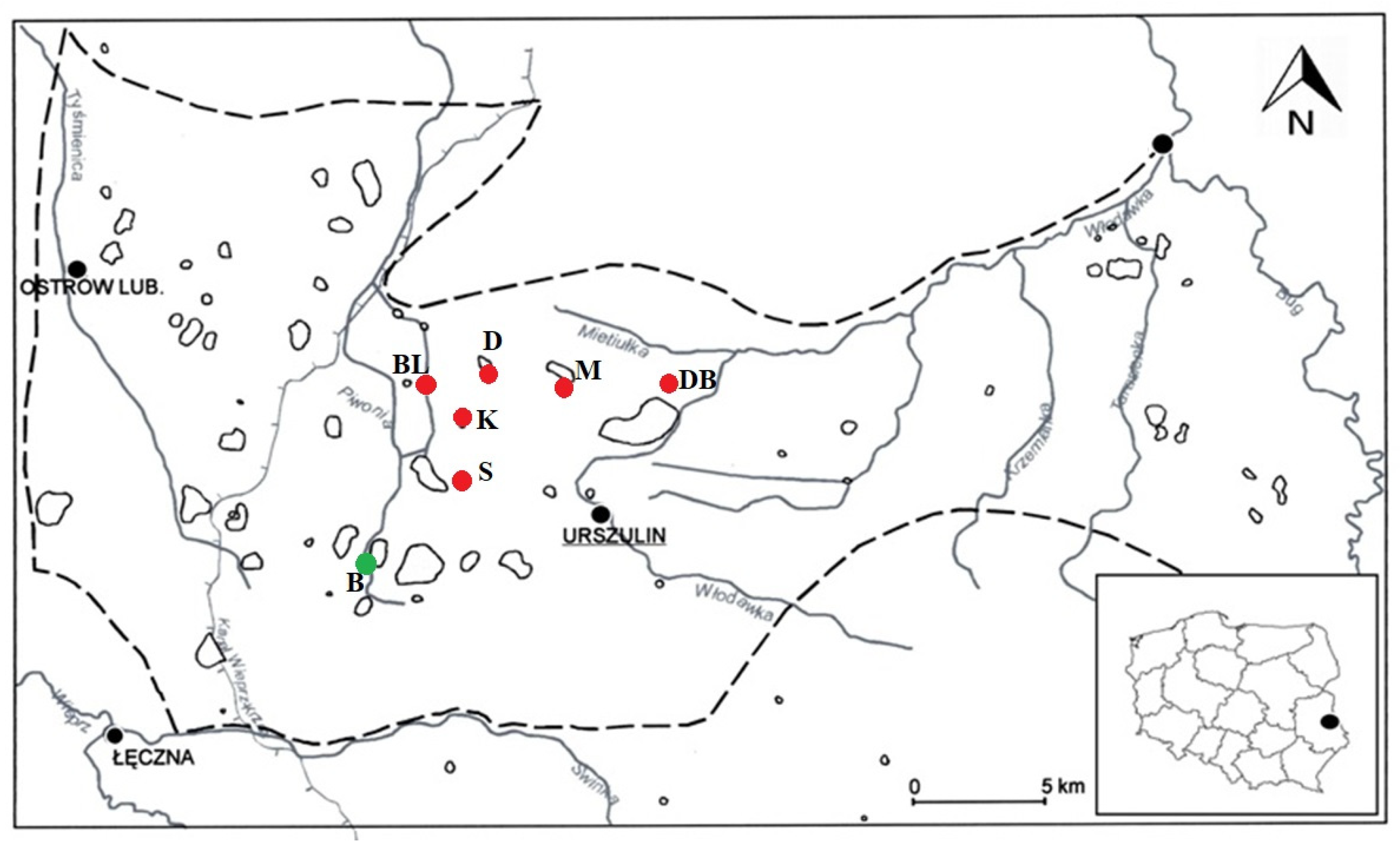
2. Materials and Methods
3. Results
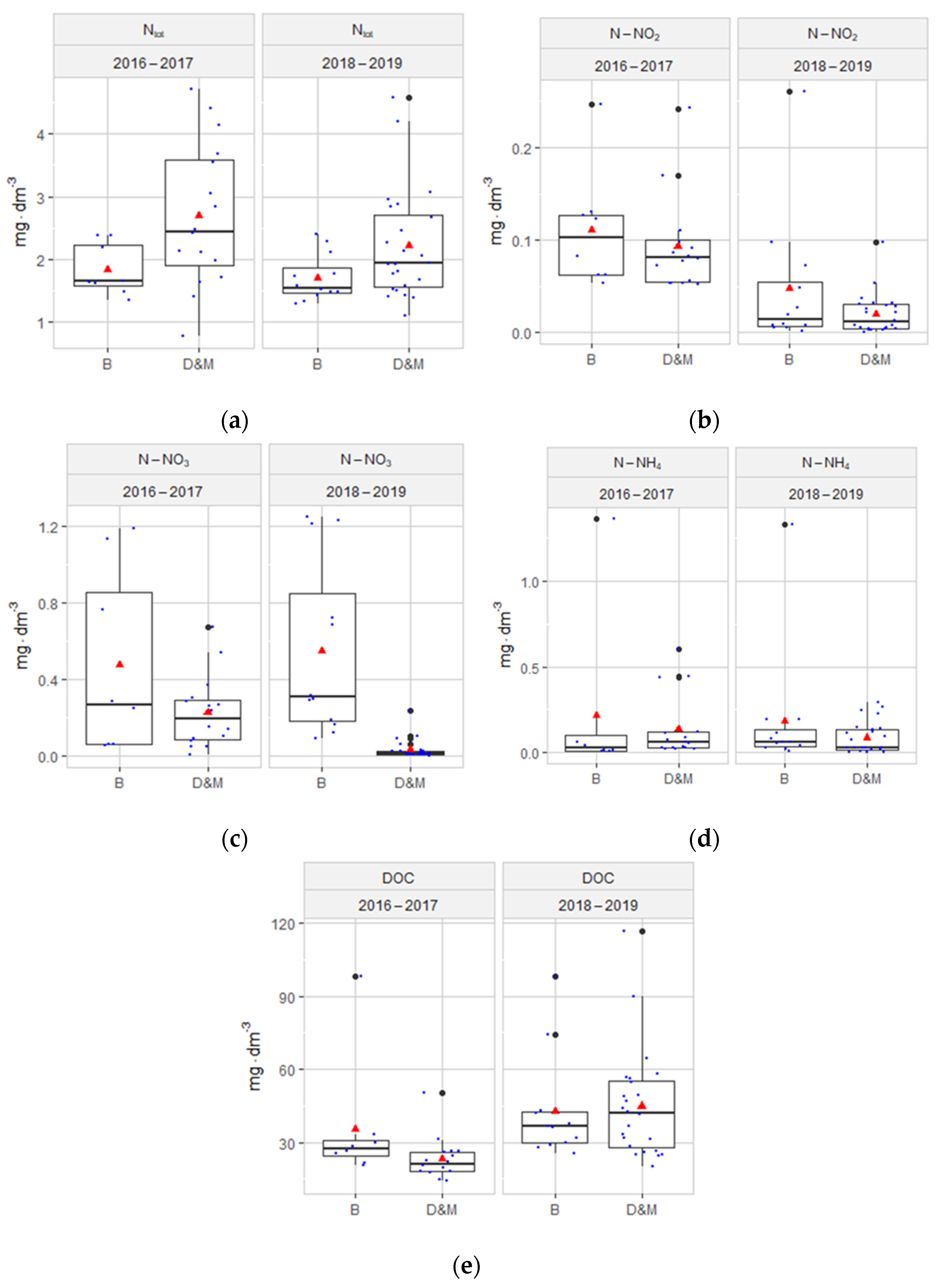
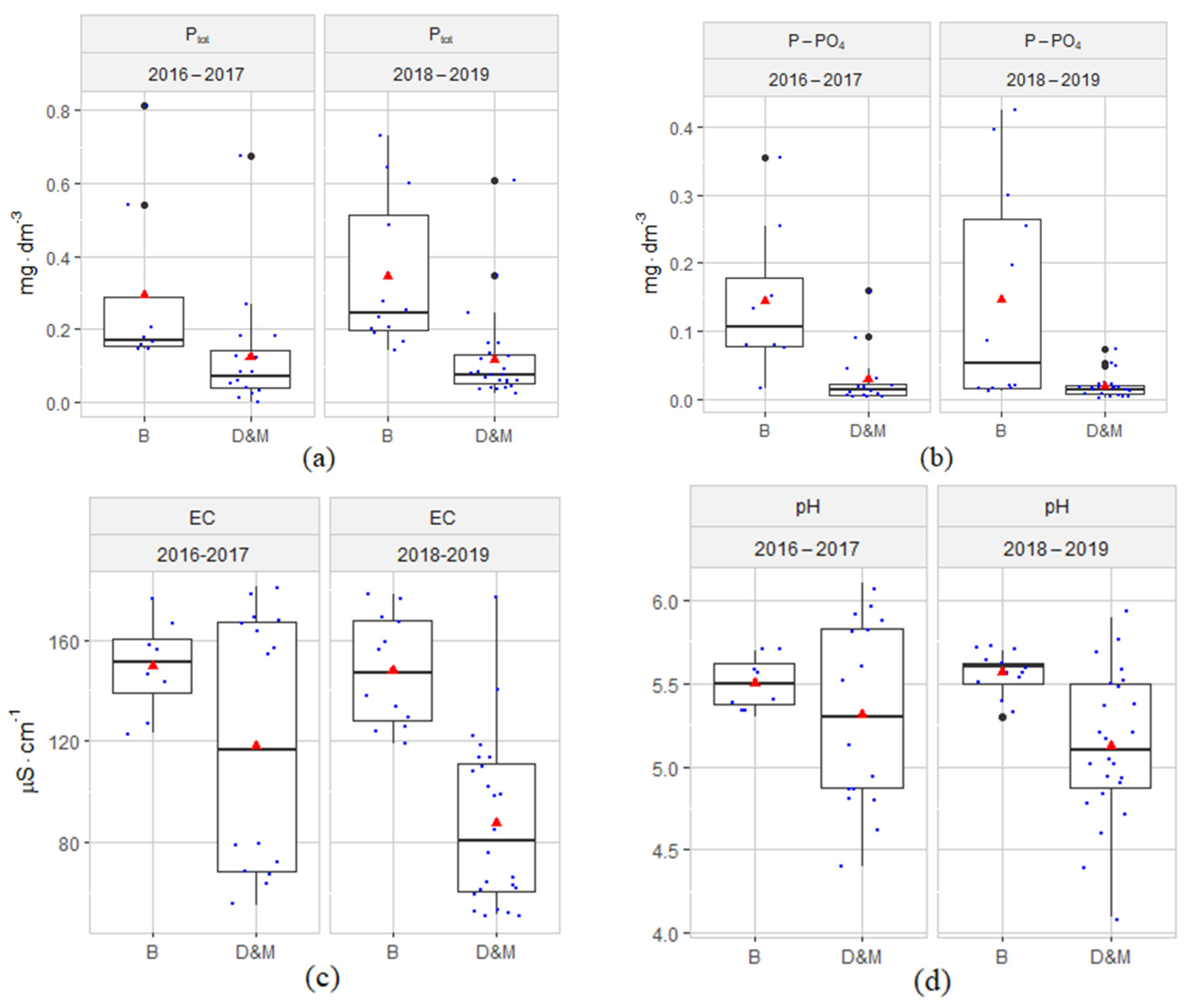
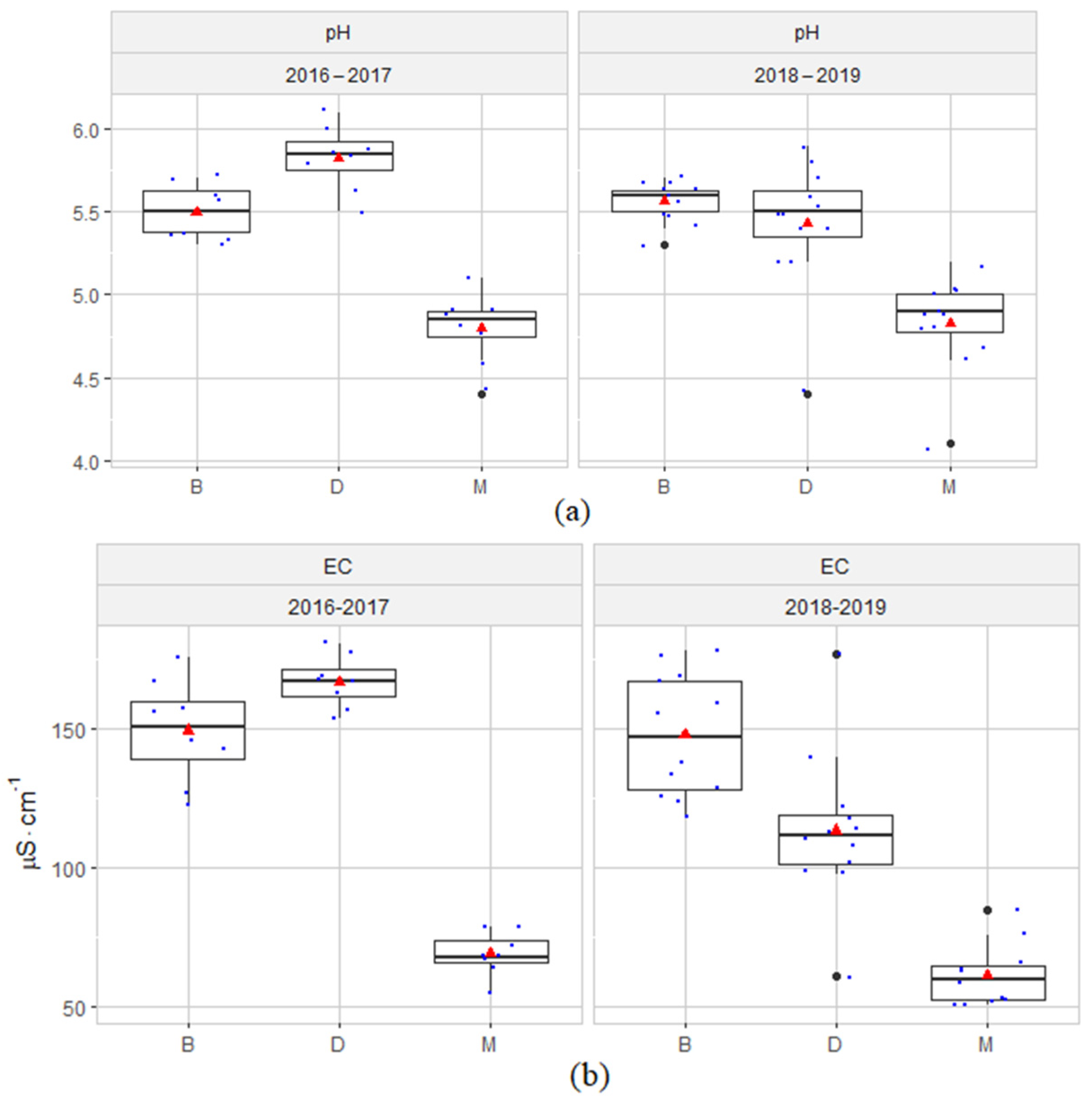
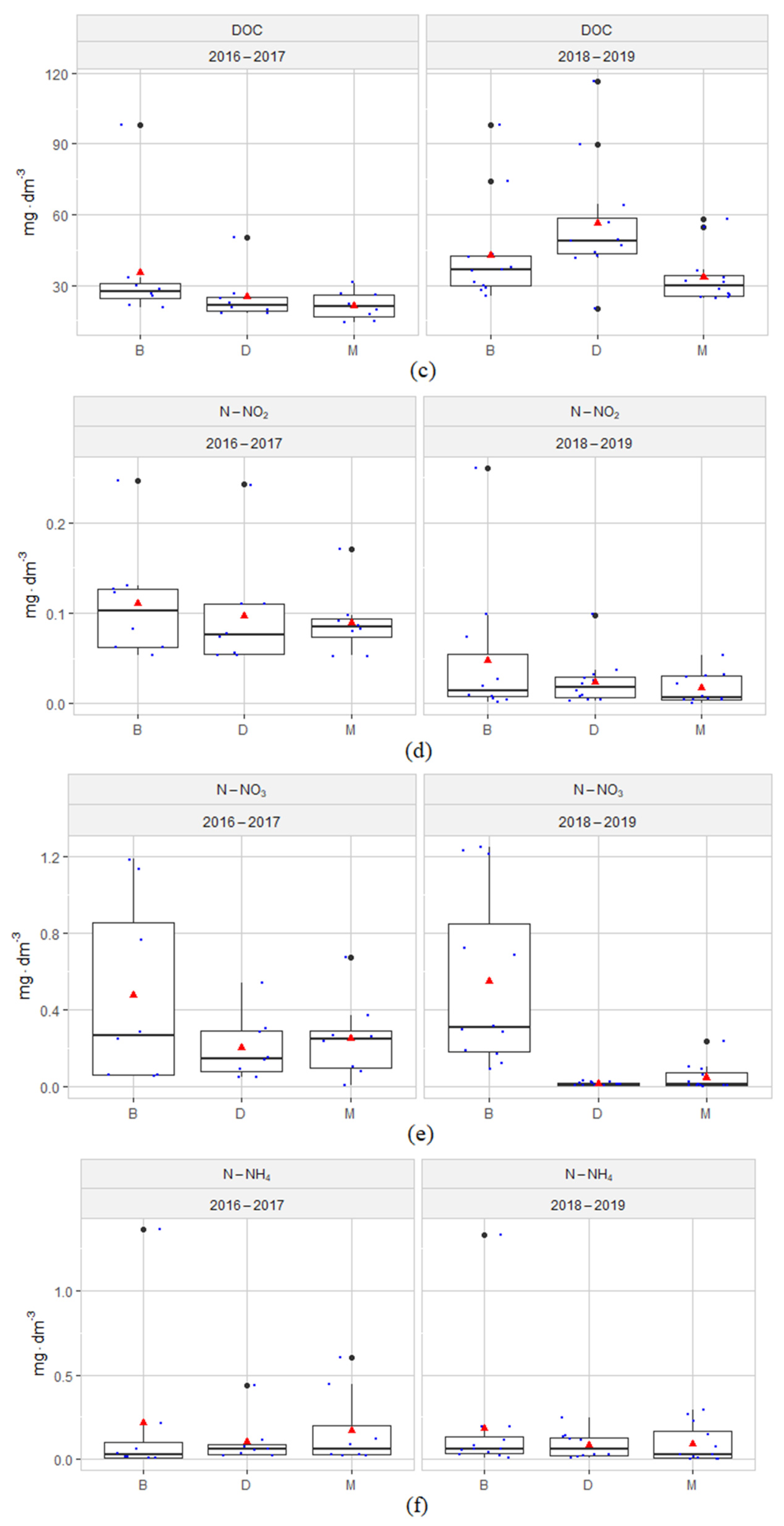
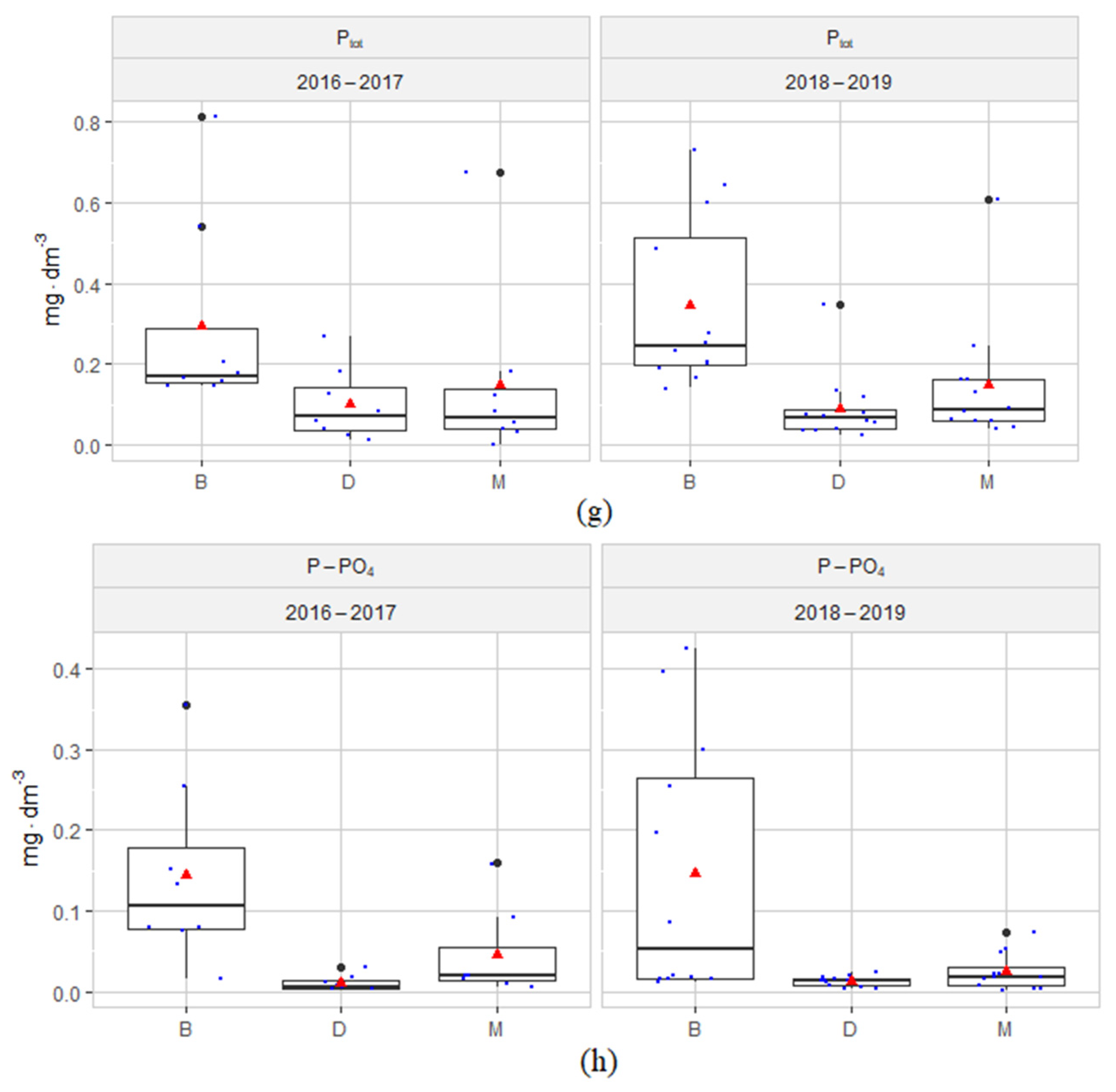
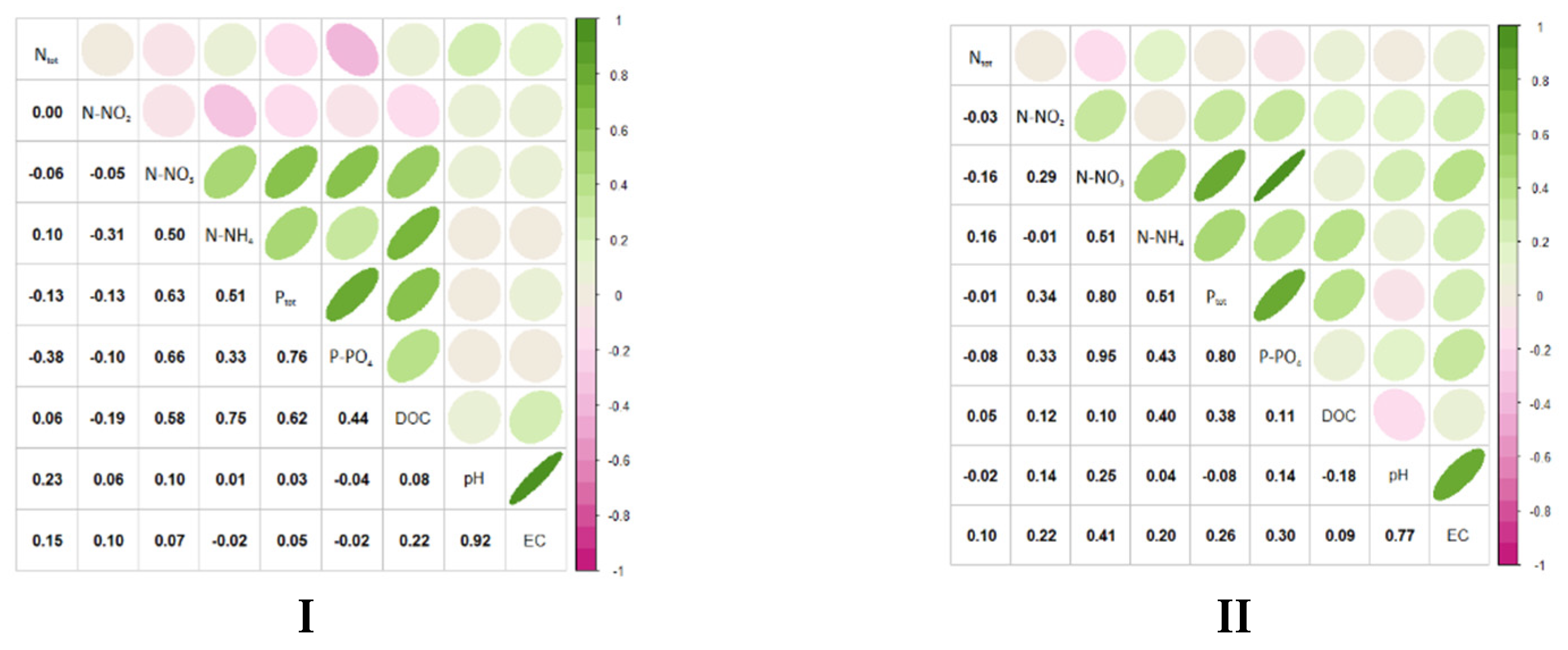
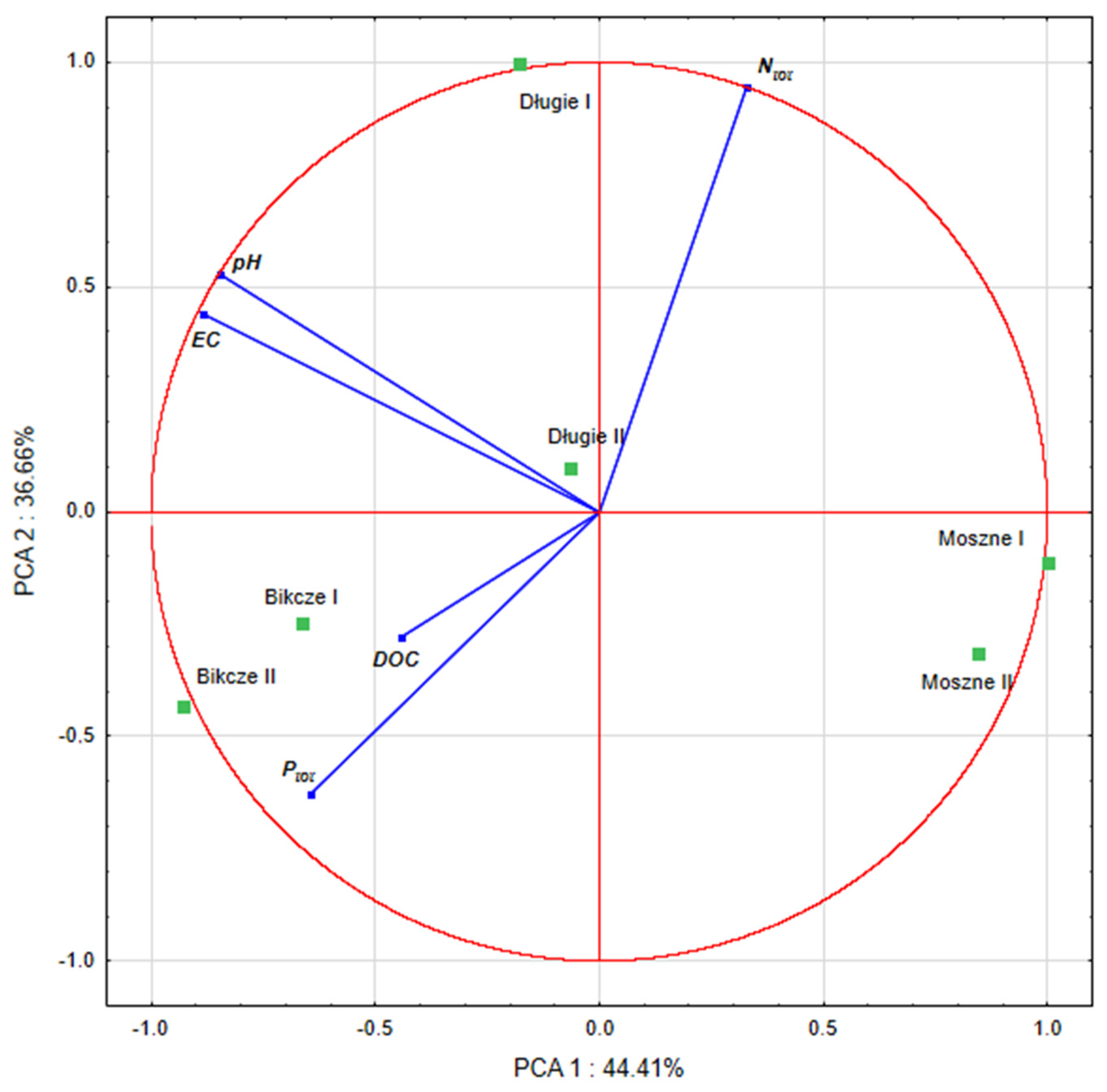
3.1. Hydrochemical Conditions of the Habitats
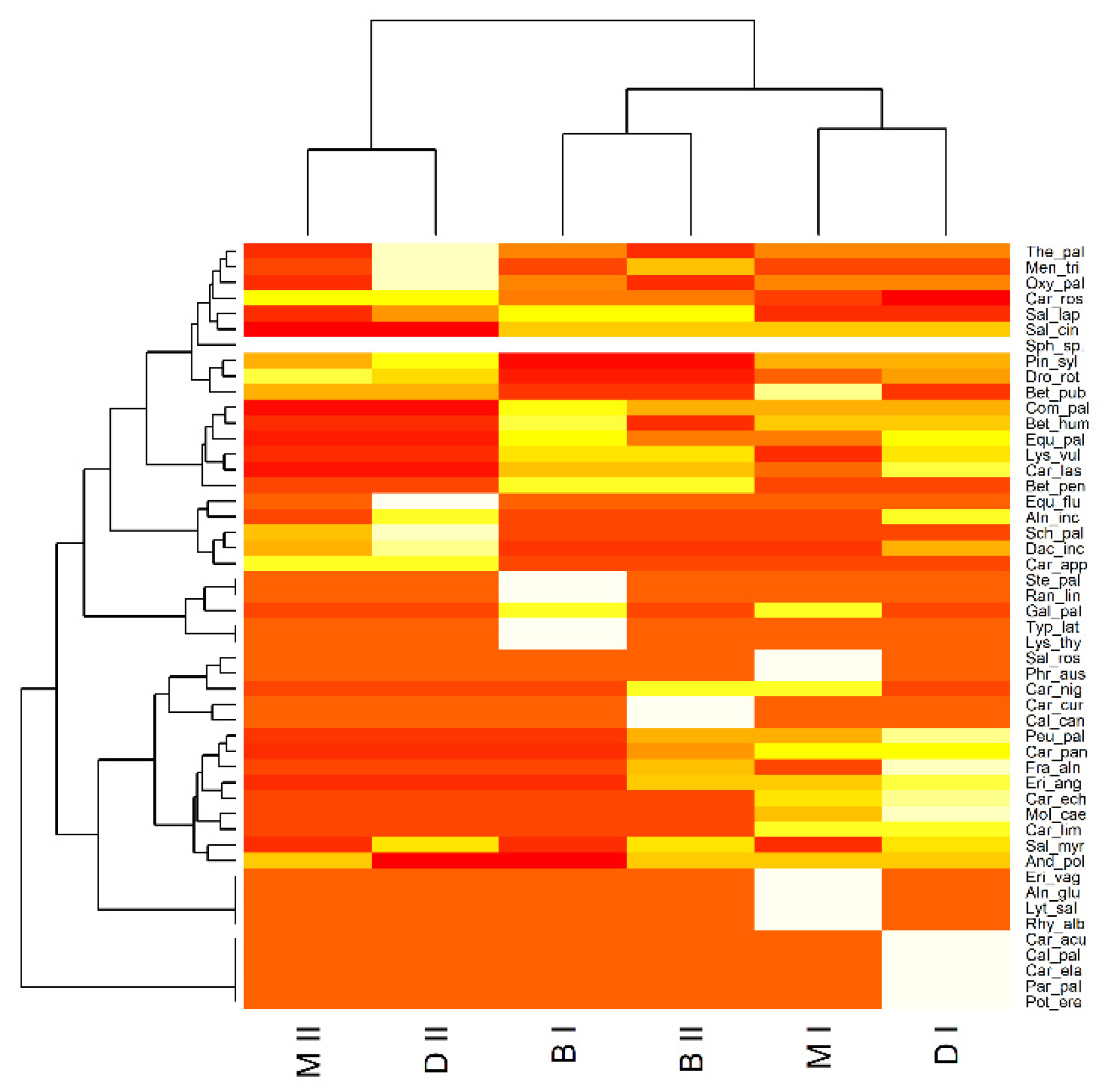
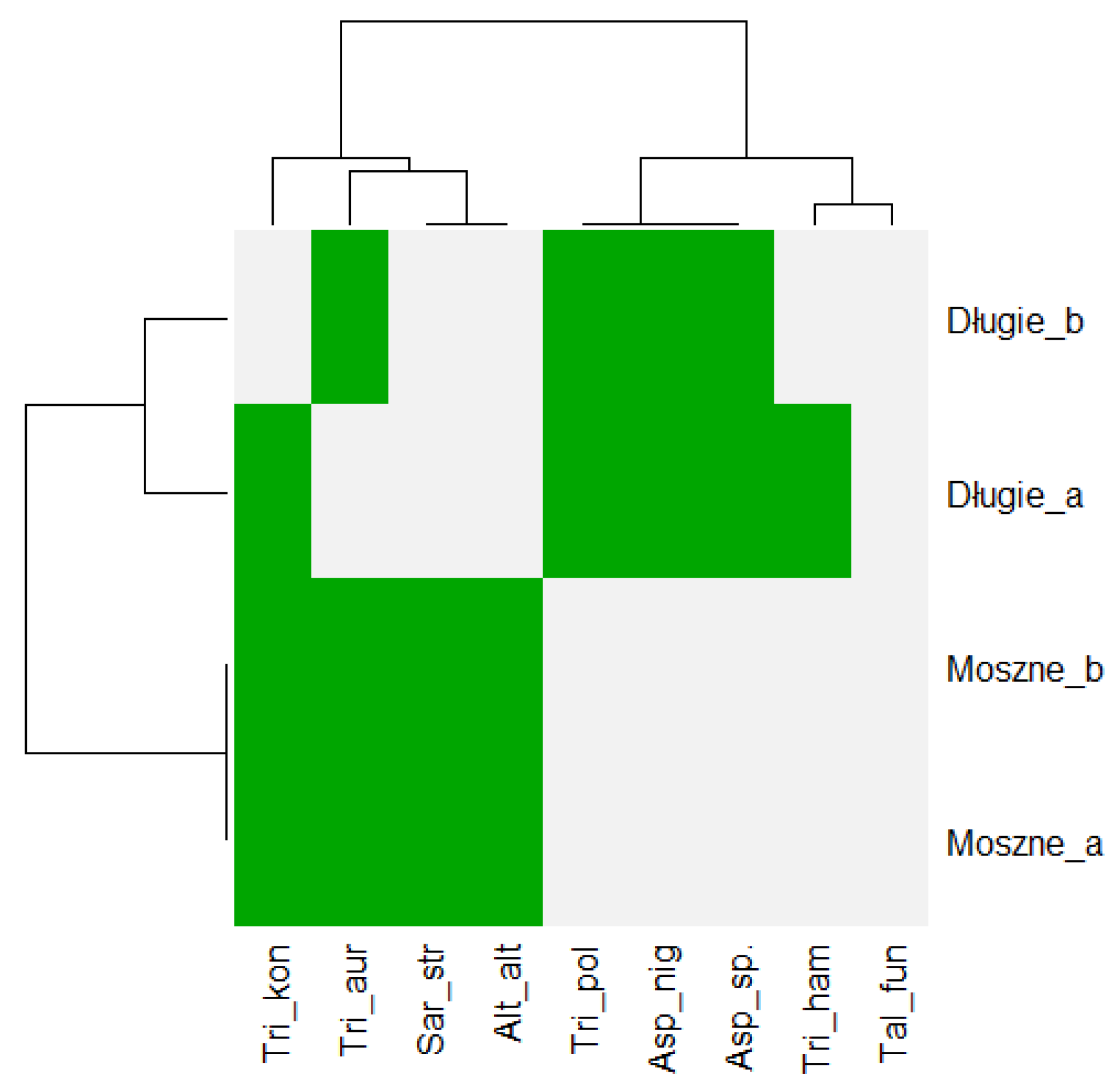
3.2. Biocoenotic Conditions of the Habitats
4. Discussion
4.1. Hydrochemical Conditions of the Habitats
4.2. Biocoenotic Conditions of the Habitats
5. Conclusions
- The hydrochemical characterization of the peatland groundwater at the sites of reintroduction of S. lapponum and changes in the parameters over time does not indicate an intensifying influx of nutrients or other hydrological disturbances caused by human activity.
- Although in 2016–2019 the values of the physical-chemical factors of the groundwater showed variability associated with the internal metabolism of the peatlands, they were within the range of the habitat preferences of the species S. lapponum.
- The minor changes in the abiotic factors of the environment observed during the study should not have a limiting effect on the S. lapponum population.
- The qualitative state of the phytocoenoses and the changes noted over the 4 years of the study at the sites where S. lapponum individuals were planted do not indicate any disturbances caused by human activity. It should be borne in mind, however, that hydrological disturbances in the habitats could accelerate processes of ecological succession associated with changes in the species composition and quantitative structure of the flora.
- Biological activity and the absence of pathogenic fungi in the soil of the sites where plants were reintroduced indicate normal functioning of soil processes, and if this biological activity is not disturbed in the near future, it should be favorable to active conservation of S. lapponum.
- The new S. lapponum populations can be expected to survive and develop at the study sites provided that the current hydrochemical stability and thus the biocoenotic stability of the habitats are maintained.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kłosowski, G.; Kłosowski, S. Flora Polski. Rośliny Wodne i Bagienne; Multico: Warszawa, Poland, 2001. [Google Scholar]
- Tobolski, K. Torfowiska na Przykładzie Ziemi Świeckiej; Towarzystwo Przyjaciół Dolnej Wisły: Toruń, Poland, 2003. [Google Scholar]
- Serafin, A.; Urban, D.; Bronowicka-Mielniczuk, U.; Szczurowska, A. To what degree can the specifics of occurrence of glacial relic Betula humilis Schrank be an indicator of habitat conditions of moderate climate peatlands? Water 2018, 10, 1062. [Google Scholar] [CrossRef] [Green Version]
- Serafin, A.; Pogorzelec, M.; Bronowicka-Mielniczuk, U. Effect of the quality of shallow groundwaters on the occurrence of selected relic plant species of peatlands in the Łęczna-Włodawa Lakeland. J. Water Land Dev. 2020, 45, 133–142. [Google Scholar] [CrossRef]
- Elven, R.; Karlsson, T. Salix L. Flora Nordica; The Bergius Foundation: Stockholm, Sweden, 2000; pp. 117–188. [Google Scholar]
- Medici, G.; Baják, P.; West, L.J.; Chapman, P.J.; Banwart, S.A. DOC and nitrate fluxes from farmland; impact on a dolostone aquifer KCZ. J. Hydrol. 2021, 595, 125658. [Google Scholar] [CrossRef]
- Williams, T.M.; Gresham, C.A. Nitrogen accumulation and changes in nitrate leaching after 4 years of intensive forest culture on marginal agricultural land. NZ For. Sci. 2000, 30, 266–279. [Google Scholar]
- Urban, D.; Wawer, M. Salix lapponum L. i S. myrtilloides L. in the area of Sobibór in the Łęczyńsko-Włodawskie Lake District. Ann. UMCS 2001, 56, 83–93. [Google Scholar]
- Churski, M.; Danielewicz, W. Salix myrtilloides in the north central Poland. Distribution, threats and conservation. Dendrobiology 2008, 60, 3–9. [Google Scholar]
- Pogorzelec, M.; Parzymies, M.; Banach-Albińska, B.; Serafin, A.; Szczurowska, A. Experimental reintroduction of the boreal species Salix lapponum L. to refuges at the southern limit of its range—Short-term results. Boreal Environ. Res. 2020, 25, 161–169. [Google Scholar]
- Pogorzelec, M. The downy willow (Salix lapponum L.) population on the peat bog near Bikcze Lake (Łęczyńsko-Włodawskie Lakeland). Acta Agrobot. 2008, 61, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Pogorzelec, M. Influence of chosen environmental abiotic factors on Salix lapponum L. populations functioning in Polesie Lubelskie Region. Pol. J. Environ. Stud. 2008, 17, 139–144. [Google Scholar]
- Pogorzelec, M. The downy willow (Salix lapponum L.) as a component of different phytocoenosis in Poleski National Park. Acta Agrobot. 2009, 62, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Zhang, Q.; Wang, Z.; Guo, Q.; Wang, J.; Liu, N.; Liang, K. Conservation and possible reintroduction of an endangered plant based on an analysis of community ecology: A case study of Primulina tabacum Hance in China. Plant Species Biol. 2010, 25, 43–50. [Google Scholar] [CrossRef]
- Ardestani, E.G.; Tarkesh, M.; Bassiri, M.; Vahabi, M.R. Potential habitat modeling for reintroduction of three native plant species in central. Iran J. Arid Land 2015, 7, 381–390. [Google Scholar] [CrossRef]
- Serafin, A.; Pogorzelec, M.; Banach, B.; Szczurowska, A.; Mielniczuk, J. Physical--chemical groundwater conditions at Salix lapponum stands in Eastern Poland. Dendrobiology 2015, 73, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Trepińska, J. Fluktuacje termiczne w Europie od małej epoki lodowej do końca XX wieku. Pr. I Studia Geogr. 2001, 29, 73–77. [Google Scholar]
- Zarzycki, K.; Korzeniak, U. (Eds.) Ecological Indicator Values of Vascular Plants of Poland; Instytut Botaniki Polskiej Akademii Nauk: Kraków, Poland, 2002. [Google Scholar]
- Pogorzelec, M.; Bronowicka-Mielniczuk, U.; Banach, B.; Szczurowska, A.; Serafin, A. Relict boreal willows (Salix lapponum and Salix myrtilloides) as an element of phytocoenoses overgrowing the water bodies in Eastern Poland. Appl. Ecol. Environ. Res. 2014, 12, 441–456. [Google Scholar] [CrossRef]
- Roncal, J.; Maschinski, J.; Schaffer, B.; Gutierrez, S.M.; Walters, D. Testing appropriate habitat outside of historic range: The case of Amorpha herbacea var. crenulata (Fabaceae). J. Nat. Conserv. 2012, 20, 109–116. [Google Scholar] [CrossRef]
- Parzymies, M.; Pogorzelec, M.; Głębocka, K.; Śliwińska, E. Genetic stability of the endangered species Salix lapponum L. regenerated in vitro during the reintroduction process. Biology 2020, 9, 378. [Google Scholar] [CrossRef]
- Buisson, E.; Anderson, S.; Holl, K.D.; Corcket, E.; Hayes, G.F.; Peeters, A.; Dutoit, T. Reintroduction of Nassella pulchra to California coastal grasslands: Effects of topsoil removal, plant neighbour removal and grazing. Appl. Veg. Sci. 2008, 11, 195–204. [Google Scholar] [CrossRef]
- Maschinski, J.; Haskins, K.E. (Eds.) Plant Reintroduction in a Changing Climate: Promises and Perils; Island Press: Washington, DC, USA, 2012. [Google Scholar] [CrossRef]
- Questad, E.J.; Kellner, J.R.; Kinney, K.; Cordell, S.; Asner, G.P.; Thaxton, J. Mapping habitat suitability for at-risk plant species and its implications for restoration and reintroduction. Ecol. Appl. 2014, 24, 385–395. [Google Scholar] [CrossRef]
- Fijałkowski, D. Studiem upon distribution and ecology of downy willow (Salix lapponum) in Łęczyńsko-Włodawskie Lakeland. Fragm. Florist. Et Geobot. 1958, 3, 89–103. [Google Scholar]
- Fijałkowski, D. The vegetation cover of lakes in the Łęczna and Włodawa area and of peat bogs adjacent to these lakes. Ann. UMCS Sect. B 1959, 14, 131–206. [Google Scholar]
- Serafin, A.; Pogorzelec, M.; Bronowicka-Mielniczuk, U. Hierarchisation of abiotic environmental properties—Methodical study for research on the ecology of plants conducted in peat bog habitats. Appl. Ecol. Environ. Res. 2019, 17, 4081–4096. [Google Scholar] [CrossRef]
- Wojciechowski, K. Pojezierze Łęczyńsko-Włodawskie; Wojewódzki Ośrodek Informacji Turystycznej: Lublin, Poland, 1983. [Google Scholar]
- Rutkowski, B. Klucz do Oznaczania Roślin Naczyniowych Polski Niżowej; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2001. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; W. Szafer Institute of Botany, Polish Academy of Science: Kraków, Poland, 2002. [Google Scholar]
- Wysocki, C.; Sikorski, P. Fitosocjologia Stosowana; Wyd. SGGW: Warszawa, Poland, 2002. [Google Scholar]
- Martyniuk, S.; Masiak, D.; Stachyra, A.; Myśków, W. Populacje drobnoustrojów strefy korzeniowej różnych traw i ich antagonizm w stosunku do Gaeumannomyces graminis var. tritici. IUNG. Pamiętnik Puławski 1991, 98, 139–144. [Google Scholar]
- Martin, J.P. Use of acid, rose bengal, and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Index Fungorum. 2019. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 10 February 2022).
- Tilaki, G.A.D.; Kamarei, R.; Vafakhah, M. Determining the relation between soil properties and spatial variability of Nitraria schoberi Linn. Using geostatistical analysis: A case study in Meighan Playa in Iran. Pol. J. Ecol. 2013, 61, 93–104. [Google Scholar]
- Serafin, A.; Pogorzelec, M.; Bronowicka-Mielniczuk, U. The specificity of natural habitats of Menyanthes trifoliata L. in peat bogs of the central part of Eastern Poland. Appl. Ecol. Environ. Res. 2017, 15, 849–859. [Google Scholar] [CrossRef]
- Serafin, A.; Sender, J.; Bronowicka-Mielniczuk, U. Potential of shrubs, shore vegetation and macrophytes of a lake to function as a phytogeochemical barrier against biogenic substances of various origin. Water 2019, 11, 290. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.P.; Seddon, P.J. Directions in reintroduction biology. Trends Ecol. Evol. 2008, 23, 20–25. [Google Scholar] [CrossRef]
- Drayton, B.; Primack, R.B. Success rates for reintroductions of eight perennial plant species after 15 years. Restor. Ecol. 2012, 20, 299–303. [Google Scholar] [CrossRef]
- Heywood, V.H. Plant conservation in the Anthropocene—Challenges and future prospects. Plant Divers. 2017, 39, 314–330. [Google Scholar] [CrossRef]
- Maschinski, J.; Albrecht, M.A. Center for plant conservation’s best practice guidelines for the reintroduction of rare plants. Plant Divers. 2017, 39, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Serafin, A.; Pogorzelec, M.; Banach, B.; Mielniczuk, J. Habitat conditions of the endangered species Salix myrtilloides in Eastern Poland. Dendrobiology 2015, 73, 55–64. [Google Scholar] [CrossRef]
- Fijałkowski, D.; Urban, D. Szata roślinna obiektu wodno-torfowiskowego “Uściwierzek” i jej przekształcenia. Ann. UMCS Sect. C 1997, 52, 119–143. [Google Scholar]
- Szafrański, C.Z.; Fiedler, M.; Bykowski, J. Czasowo przestrzenna zmienność chemizmu wód na Pojezierzu Gnieźnieńskim na przykładzie obiektu Mokronosy [Groundwater quality in the agricultural microcatchment on Gniezno Lakeland]. Przegląd Nauk. Wydziału Melior. I Inżynierii Sr. 1998, 16, 262–269. [Google Scholar]
- Marcinek, J.; Komisarek, J.; Kaźmierowski, C. Dynamika składników rozpuszczalnych w wodach gruntowych uprawnych gleb płowych i czarnych ziem. Roczniki AR Poznań. Melior. I Inżynieria Sr. 1994, 268, 69–82. [Google Scholar]
- Grzywna, A. Chemiczne wskaźniki jakości wody w zlewni lasów parczewskich. Inżynieria Ekol. 2014, 36, 120–127. [Google Scholar] [CrossRef]
- Maciak, F. Ocena aktywności biologicznej murszów i torfów na podstawie mineralizacji związków węgla i azotu. Rocz. Glebozn. 1995, 46, 19–28. [Google Scholar]
- Kaszewski, B. Warunki Klimatyczne Lubelszczczyzny; Wyd. UMCS: Lublin, Poland, 2008. [Google Scholar]
- Sapek, B. Nagromadzanie i uwalnianie fosforu w glebach—Źródła, procesy, przyczyny. Woda-Środowisko-Obsz. Wiej. 2014, 14, 77–100. [Google Scholar]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry, an Introduction Emphasizing Chemical Equilibria in Natural Waters, 2nd ed.; Wiley: New York, NY, USA, 1981. [Google Scholar]
- Banaszuk, P.; Kamocki, A.K. Statyczny jakościowy bilans wodny jako podstawa renaturyzacji ekosystemów hydrogenicznych w dolinie górnej Biebrzy. Inżynieria Ekologiczna. Ecol. Eng. 2016, 48, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Brysiewicz, A.; Wesołowski, P.; Rawicki, K. Porównanie stężenia składników chemicznych w wodzie ze śródpolnego oczka wodnego oraz w wodzie gruntowej z przylegających terenów rolniczych. Woda-Środowisko-Obsz. Wiej. 2013, 13, 17–31. [Google Scholar]
- Koerselman, W.; Bakker, S.A.; Blom, M. Nitrogen, phosphorus and potassium budgets for two small fens surrounded by heavily fertilized pastures. J. Ecol. 1990, 78, 428–442. [Google Scholar] [CrossRef]
- Foy, R.H.; Withers, P.J.A. The contribution of Agricultural Phosphorus to Eutrophication. Nawozy I Nawożenie 2002, 3, 20–49. [Google Scholar]
- Igras, J.; Jadczyszyn, T. Zawartość azotanów i fosforanów w płytkich wodach gruntowych w Polsce. Probl. Inżynierii Rol. 2008, 16, 91–102. [Google Scholar]
- Lamers, L.P.M.; Vile, M.A.; Grootjans, A.P.; Acreman, M.C.; Diggelen, R.; Evans, M.G.; Richardson, C.J.; Rochefort, L.; Kooijman, A.M.; Roelofs, J.G.M.; et al. Ecological restoration of rich fens in Europe and North America: From trial and error to an evidence-based approach. Biol. Rev. 2015, 90, 182–203. [Google Scholar] [CrossRef] [Green Version]
- Smolders, A.J.P.; Lamers, L.P.M.; Lucassen, E.C.H.E.T.; van der Velde, G.; Roelofs, J.G.M. Internal eutrophication: How it works and what to do about it—A review. Chem. Ecol. 2006, 22, 93–111. [Google Scholar] [CrossRef]
- Jabłońska, E. Brzoza Niska Betula Humilis Schrank w Polsce—Status Fitocenotyczny, Warunki Siedliskowe, Zagrożenia i Ochrona. Doctoral Dissertation, University of Warsaw, Warsaw, Poland, 2009. Available online: https://www.researchgate.net/publication/270283403 (accessed on 10 February 2022).
- Górniak, A. Substancje Humusowe i Ich Rola w Funkcjonowaniu Ekosystemów Słodkowodnych; Uniwersytet Warszawski, filia w Białymstoku: Białystok, Poland, 1996. [Google Scholar]
- Olaczek, R.; Kucharski, L.; Pisarek, W. Zanikanie obszarów podmokłych i jego skutki środowiskowe na przykładzie województwa piotrkowskiego (zlewni Pilicy i Warty). Studia Ośrodka Dok. Fizjogr. 1990, 18, 141–199. [Google Scholar]
- Herbich, J. Zróżnicowanie i problemy ochrony roślinności torfowisk Pojezierza Kaszubskiego. Acta Bot. Cassubica 2001, 1, 59–69. [Google Scholar]
- Falińska, K. Ekologia Roślin; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2004. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; PWN: Warszawa, Poland, 2005; ISBN 83-01-14439-4. [Google Scholar]
- Brockwell, J.; Bottomley, P.J.; Thies, J.E. Manipulation of rhizobia microflora for improving legume productivity and soil fertility: A critical assessment. Plant Soil 1995, 174, 143–180. [Google Scholar] [CrossRef]
- Hanus-Fajerska, E.; Muszyńska, E.; Giemzik, A. Review on studies of zinc-lead waste heaps microbiota. Arch. Gospod. Odpadami I Ochr. Środowiska 2015, 17, 59–68. [Google Scholar]
- Głowacki, A.; Mocek-Płóciniak, A.; Spychalski, W.; Kayzer, D. Wpływ wieloletniej rekultywacji na właściwości mikrobiologiczne gleb wytworzonych z gruntów pogórniczych. Soil Sci. Annu. 2020, 71, 359–370. [Google Scholar] [CrossRef]
- Zwoliński, J. Oznaczanie udziału grzybów i bakterii w biomasie drobnoustrojów gleb leśnych. Leśne Pr. Badaw. 2005, 4, 7–18. [Google Scholar]
- Galus-Barchan, A.; Paśmionka, I. Występowanie wybranych mikroorganizmów w glebie na obszarze Puszczy Niepołomickiej ze szczególnym uwzględnieniem grzybów pleśniowych. Pol. J. Agron. 2014, 17, 11–17. [Google Scholar]
- Badura, L. Mikroorganizmy w Glebach—Ich rola i znaczenie. Inż. Środ. Półrocznik AGH Kraków 2004, 9, 147–158. [Google Scholar]
- Corredor, A.H.; Van Rees, K.; Vujanovic, V. Changes in root-associated fungal assemblages within newly established clonal biomass plantations of Salix spp. For. Ecol. Manag. 2012, 282, 105–114. [Google Scholar] [CrossRef]
- Zalewska, E.D.; Pogorzelec, M.; Król, E.D.; Serafn, A. Fungi inhabiting the aboveground organs of downy willow (Salix lapponum L.) and swamp willow (Salix myrtilloides L.). Acta Mycol. 2019, 54, 1134. [Google Scholar] [CrossRef]
- Kosicka-Dziechciarek, D.; Tatuśko, N. Wpływ izolatu Trichoderma harzianum oraz słomy pszennej na liczebność grzybów pleśniowych i bakterii w glebie. Woda-Środowisko-Obsz. Wiej. 2017, 17, 111–125. [Google Scholar]
- Zin, A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Study Site | Kruskal-Wallis Chi-Squared | p-Value |
|---|---|---|---|
| Ntot | B | 0.788 | 0.3746 |
| D | 4.339 | 0.0372 | |
| M | 0.054 | 0.8170 | |
| N-NO3 | B | 0.932 | 0.3345 |
| D | 13.735 | 0.0002 | |
| M | 7.319 | 0.0068 | |
| N-NH4 | B | 0.860 | 0.3536 |
| D | 0.013 | 0.9078 | |
| M | 1.526 | 0.2167 | |
| N-NO2 | B | 6.487 | 0.0109 |
| D | 11.031 | 0.0009 | |
| M | 13.208 | 0.0003 | |
| Ptot | B | 1.527 | 0.2165 |
| D | 0.0536 | 0.817 | |
| M | 0.788 | 0.3746 | |
| P-PO4 | B | 0.150 | 0.6984 |
| D | 0.866 | 0.3522 | |
| M | 0.658 | 0.4174 | |
| pH | B | 0.636 | 0.4252 |
| D | 7.409 | 0.0065 | |
| M | 0.303 | 0.5820 | |
| EC | B | 0.002 | 0.9692 |
| D | 10.500 | 0.0012 | |
| M | 4.519 | 0.0335 | |
| DOC | B | 3.434 | 0.0639 |
| D | 8.149 | 0.0043 | |
| M | 6.881 | 0.0087 |
| CLASS | MAGNOLIOPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical Family | Species | |||||||
| Apiaceae | Peucedanum palustre | + | + | + | ||||
| Betulaceae | Alnus glutinosa | + | ||||||
| Alnus incana | + | + | ||||||
| Betula humilis | + | + | + | |||||
| Betula pendula | + | + | ||||||
| Betula pubescens | + | + | + | + | + | + | ||
| Caryophyllaceae | Stellaria palustris | + | ||||||
| Droseraceae | Drosera rotundifolia | + | + | + | + | |||
| Ericaceae | Andromeda polifolia | + | + | + | + | |||
| Oxycoccus palustris | + | + | + | + | + | + | ||
| Lythraceae | Lythrum salicaria | + | ||||||
| Menyanthaceae | Menyanthes trifoliata | + | + | + | + | + | + | |
| Parnassicaceae | Parnassia palustris | + | ||||||
| Primulaceae | Lysimachia thyrsiflora | + | ||||||
| Lysimachia vulgaris | + | + | + | |||||
| Ranunculaceae | Ranunculus lingua | + | ||||||
| Rhamnaceae | Frangula alnus | + | + | |||||
| Rubiaceae | Galium palustre | + | + | |||||
| Rosaceae | Comarum palustre | + | + | + | + | |||
| Potentilla erecta | + | |||||||
| Salicaceae | Salix cinerea | + | + | + | + | + | + | |
| Salix lapponum | + | + | + | + | + | + | ||
| Salix mytrylloides | + | + | + | |||||
| Salix rosmarinifolia | + | |||||||
| Scheuchzeriaceae | Scheuchzeria palustris | + | + | |||||
| CLASS | LILIOPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical family | Species | |||||||
| Araceae | Calla palustris | + | ||||||
| Cyperaceae | Carex acutiformis | + | ||||||
| Carex appropinquata | + | + | ||||||
| Carex curta | + | |||||||
| Carex echinata | + | + | ||||||
| Carex elata | + | |||||||
| Carex lasiocarpa | + | + | + | + | ||||
| Carex limosa | + | + | ||||||
| Carex nigra | + | + | ||||||
| Carex panicea | + | + | + | |||||
| Carex rostrata | + | + | + | + | + | |||
| Eriophorum angustifolium | + | + | + | |||||
| Eriophorum vaginatum | + | |||||||
| Rhynchospora alba | + | |||||||
| Orchidaceae | Dactylorhiza incarnata | + | + | + | ||||
| Poaceae | Calamagrostis canescens | + | ||||||
| Molinia caerulea | + | + | ||||||
| Phragmites australis | + | |||||||
| Typhaceae | Typha latifolia | + | ||||||
| CLASS | EQUISETOPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical family | Species | |||||||
| Equisetaceae | Equisetum fluviatile | + | ||||||
| Equisetum palustre | + | + | + | + | ||||
| CLASS | POLIPODIOPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical family | Species | |||||||
| Thelypteriodaceae | Thelypteris palustris | + | + | + | + | + | + | |
| CLASS | CONIFEROPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical family | Species | |||||||
| Pinaceae | Pinus sylvestris | + | + | + | + | |||
| CLASS | SPHAGNOPSIDA | I | II | |||||
| M | D | B | M | D | B | |||
| Botanical family | Species | |||||||
| Sphagnaceae | Sphagnum sp. | + | + | + | + | + | + | |
| Total | 49 | 29 | 30 | 19 | 14 | 15 | 22 | |
| 44 | 29 | |||||||
| I | II | ||||
|---|---|---|---|---|---|
| M–D | M–B | D–B | M–D | M–B | D–B |
| 0.513 | 0.375 | 0.333 | 0.75 | 0.36 | 0.321 |
| TOTAL | |||||
| M–M | B–B | D–D | |||
| 0.37 | 0.5 | 0.363 | |||
| Parameter/Soil | Research Site | |||
|---|---|---|---|---|
| M | D | |||
| Rhizosphere | Non-RhizoSphere | Rhizosphere | Non-RhizoSphere | |
| Dry weight | 0.7 | 0.11 | 0.53 | 0.44 |
| [g] | ||||
| Bacterial count | 51.43 × 10−6 | 30.08 × 10−6 | 2.13 × 10−6 | 8.54 × 10−6 |
| [CFU × g−1 g DW] | ||||
| Fungal | 225.71 × 10−3 | 186.22 × 10−3 | 254.1 × 10−3 | 226.33 × 10−3 |
| [CFU × g−1 g DW] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serafin, A.; Pogorzelec, M.; Banach-Albińska, B.; Zalewska, E.; Bronowicka-Mielniczuk, U.; Arciszewski, M. The Importance of Groundwater Quality and Other Habitat Parameters for Effective Active Protection of an Endangered Plant Species in Eastern Poland. Water 2022, 14, 1270. https://doi.org/10.3390/w14081270
Serafin A, Pogorzelec M, Banach-Albińska B, Zalewska E, Bronowicka-Mielniczuk U, Arciszewski M. The Importance of Groundwater Quality and Other Habitat Parameters for Effective Active Protection of an Endangered Plant Species in Eastern Poland. Water. 2022; 14(8):1270. https://doi.org/10.3390/w14081270
Chicago/Turabian StyleSerafin, Artur, Magdalena Pogorzelec, Barbara Banach-Albińska, Ewa Zalewska, Urszula Bronowicka-Mielniczuk, and Michał Arciszewski. 2022. "The Importance of Groundwater Quality and Other Habitat Parameters for Effective Active Protection of an Endangered Plant Species in Eastern Poland" Water 14, no. 8: 1270. https://doi.org/10.3390/w14081270
APA StyleSerafin, A., Pogorzelec, M., Banach-Albińska, B., Zalewska, E., Bronowicka-Mielniczuk, U., & Arciszewski, M. (2022). The Importance of Groundwater Quality and Other Habitat Parameters for Effective Active Protection of an Endangered Plant Species in Eastern Poland. Water, 14(8), 1270. https://doi.org/10.3390/w14081270