
The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains

Abstract
:1. Introduction
2. Materials and Methods
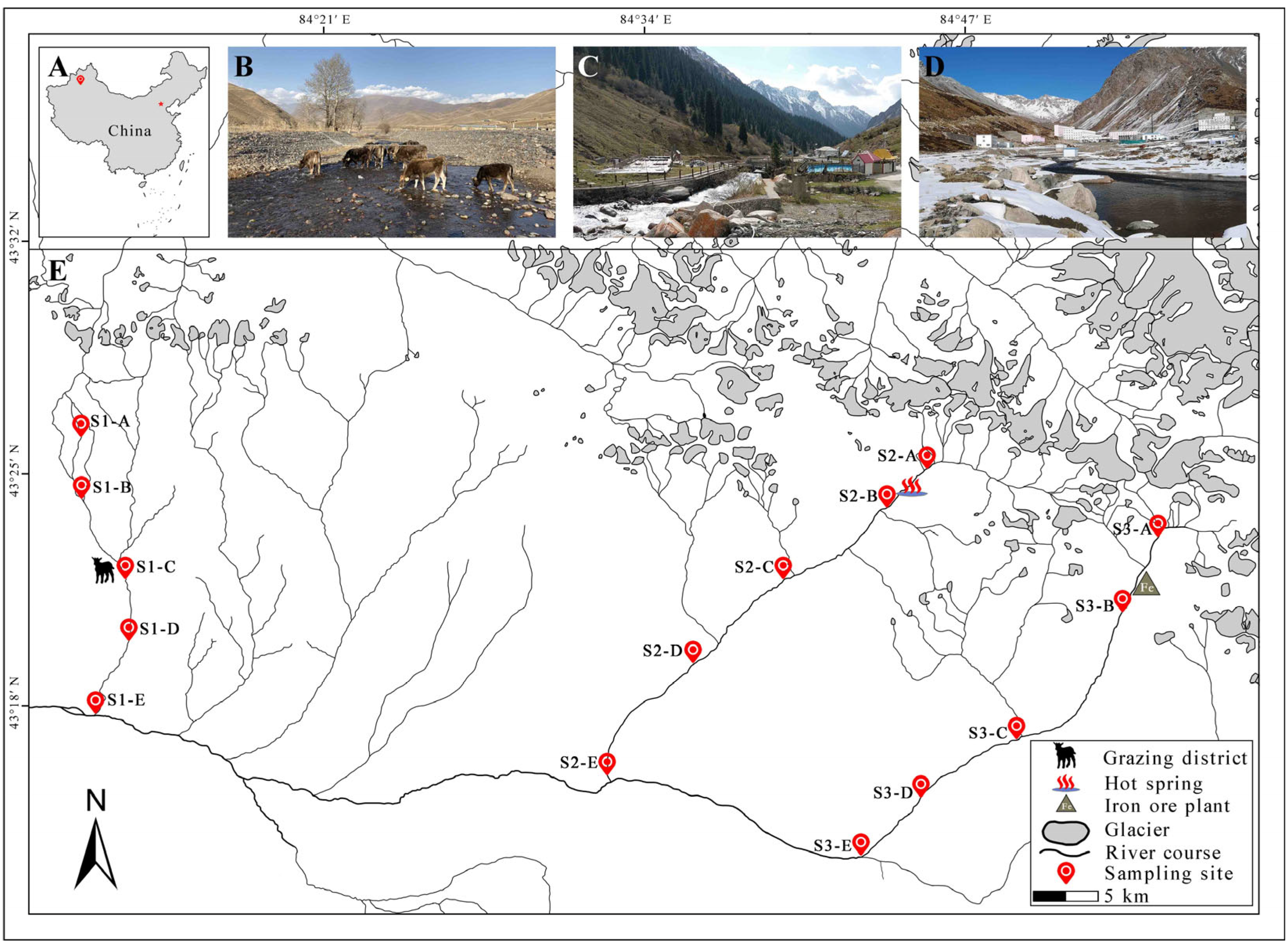
2.1. Study Area
2.2. Macroinvertebrate Collection and Functional Traits
2.3. Measurements of Environmental Factors
2.4. Data Analysis
3. Results
3.1. Environmental Factors
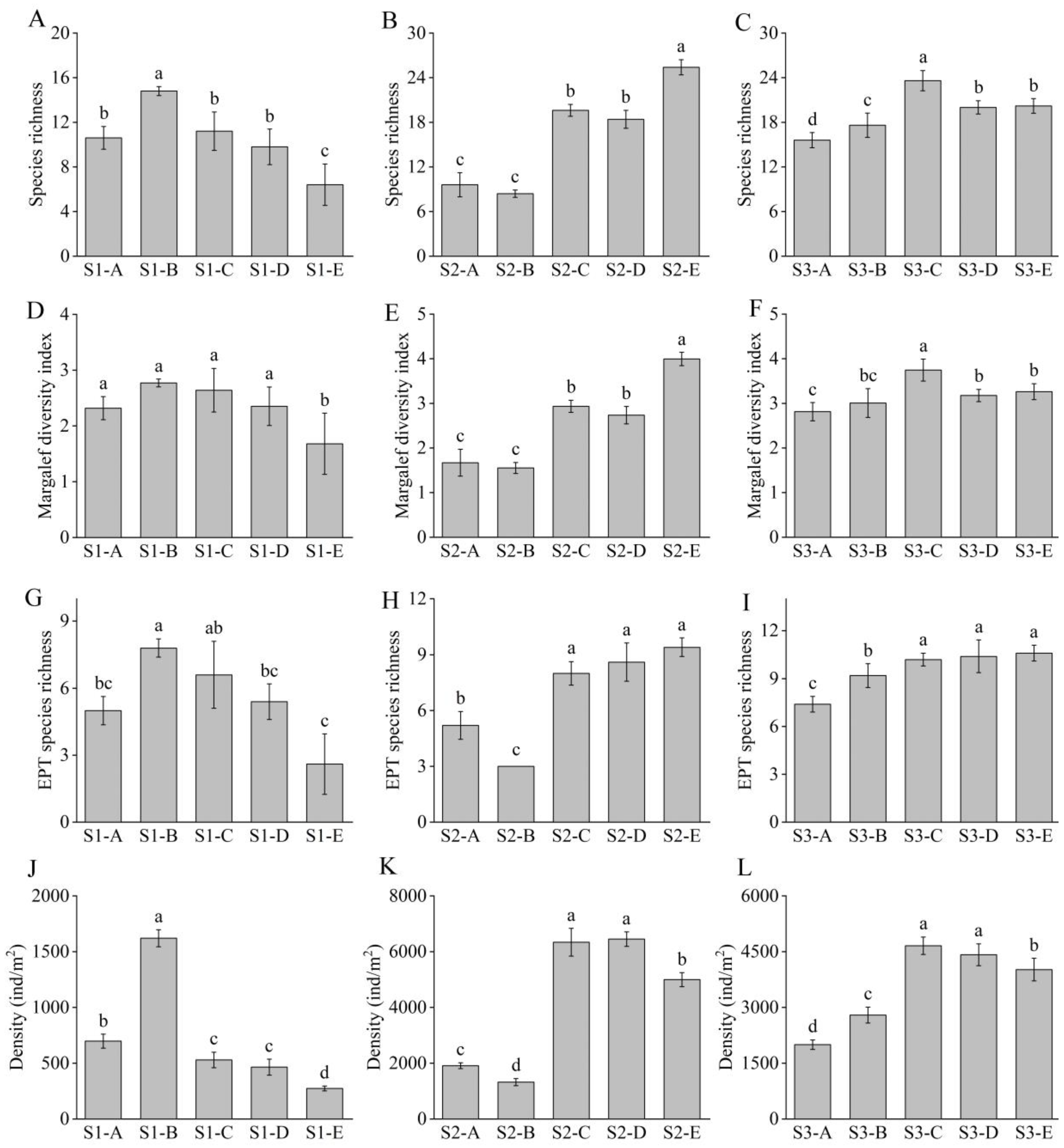
3.2. Macroinvertebrate Community Structure in Three Disturbed Streams
3.3. Functional Traits of Macroinvertebrates
3.4. Relationships between Macroinvertebrate Communities and Functional Traits and Environmental Factors
4. Discussion
4.1. The Effects of Three Disturbances on Stream Environmental Factors
4.2. The Effects of Three Disturbances on Macroinvertebrate Community Structure
4.3. The Effects of Three Disturbances on Macroinvertebrate Functional Traits
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Order | Family | Subfamily/Genus | Grazing Stream | Hot Spring Stream | Mining Stream |
|---|---|---|---|---|---|---|
| Oligochaeta | Haplotaxida | Naididae sp. | + | + | ||
| Gastropoda | Basommatophora | Planorbidae | Anis sp. | + | ||
| Malacostraca | Amphipoda | Gammaridae | Gammarus sp. | + | ||
| Insect | Ephemeroptera | Heptageniidae | Iron sp. | + | + | + |
| Cinygmula sp. | + | + | ||||
| Heptagenia sp. | + | |||||
| Rhithrogena sp. | + | + | + | |||
| Baetidae | Baetis sp. | + | + | + | ||
| Ameletidae | Ameletus sp. | + | + | |||
| Ephemerellidae | Drunella sp. | + | ||||
| Plecoptera | Capniidae | Capnia sp. | + | + | ||
| Taeniopterygidae | Rhabdiopteryx sp. | + | ||||
| Perlodidae | Arcynopteryx sp. | + | ||||
| Isoperla sp. | + | + | + | |||
| Diura sp. | + | |||||
| Perlodes sp. | + | + | ||||
| Nemouridae | Nemoura sp. | + | + | + | ||
| Chloroperlidae | Suwallia sp. | + | + | |||
| Trichoptera | Limnephilidae | Pseudostenophylax sp. | + | + | + | |
| Lepidostomatidae | Theliopsyche sp. | + | + | + | ||
| Lepidostoma sp. | + | + | ||||
| Brachycentridae | Brachycentrus sp. | + | + | + | ||
| Glossosomatidae | Glossosoma sp. | + | + | + | ||
| Hydropsychidae | Hydropsyche sp. | + | ||||
| Diptera | Tipulidae | Dicranota sp. | + | + | + | |
| Cheilotrichia sp. | + | + | ||||
| Cryptolabis sp. | + | |||||
| Antocha sp. | + | + | + | |||
| Tipula (Arctotipula) sp. | + | |||||
| Tipula (Sinotipula) sp. | + | + | ||||
| Hexotoma sp. | + | + | + | |||
| Pedica sp. | + | |||||
| Chironomidae | Orthocladiinae sp.1 | + | + | + | ||
| Orthocladiinae sp.2 | + | + | + | |||
| Orthocladiinae sp.3 | + | + | + | |||
| Orthocladiinae sp.4 | + | + | ||||
| Tanytansini sp. | + | + | + | |||
| Chironomini sp. | + | + | ||||
| Tanypodinae sp. | + | + | ||||
| Diamesinae | Boreoheptagyia sp. | + | + | |||
| Simuliidae | Simulium sp. | + | + | + | ||
| Empididae sp.1 | + | + | ||||
| Empididae sp.2 | + | + | ||||
| Blephariceridae | Blepharicera sp. | + | + | |||
| Ceratopogonidae sp. | + | |||||
| Psychodidae | Pericoma sp. | + | + | + | ||
| Dixdae | Dixa sp. | + | ||||
| Arachnida | Acariformes | Hydrachnellae sp. | + | + |
References
- Crespo-Pérez, V.; Dangles, O.; Ibarra, C.; Espinosa, R.; Andino, P.; Jacobsen, D.; Cauvy-Fraunié, S. Functional structure and diversity of invertebrate communities in a glacierised catchment of the tropical Andes. Freshw. Biol. 2020, 65, 1348–1362. [Google Scholar] [CrossRef]
- Milner, A.M.; Khamis, K.; Battin, T.J.; Brittain, J.E.; Barrand, N.E.; Füreder, L.; Cauvy-Fraunié, S.; Gíslason, G.M.; Jacobsen, D.; Hannah, D.M.; et al. Glacier shrinkage driving global changes in downstream systems. Proc. Natl. Acad. Sci. USA 2017, 114, 9770–9778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotti, A.; Jacobsen, D.; Tappeiner, U.; Bottarin, R. Spatial and temporal variation of benthic macroinvertebrate assemblages during the glacial melt season in an Italian glacier-fed stream. Hydrobiologia 2019, 827, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, D.; Dangles, O.; Andino, P.; Espinosa, R.; Hamerlík, L.; Cadier, E. Longitudinal zonation of macroinvertebrates in an Ecuadorian glacier-fed stream: Do tropical glacial systems fit the temperate model? Freshw. Biol. 2010, 55, 1234–1248. [Google Scholar] [CrossRef]
- Cauvy-Fraunié, S. and Dangles, O. A global synthesis of biodiversity responses to glacier retreat. Nat. Ecol. Evol. 2019, 3, 1675–1685. [Google Scholar] [CrossRef]
- Espinoza-Toledo, A.; Mendoza-Carranza, M.; Castillo, M.M.; Barba-Macías, E.; Capps, K.A. Taxonomic and functional responses of macroinvertebrates to riparian forest conversion in tropical streams. Sci. Total Environ. 2021, 757, 143972. [Google Scholar] [CrossRef]
- Shain, D.H.; Novis, P.M.; Cridge, A.G.; Zawierucha, K.; Geneva, A.J.; Dearden, P.K. Five animal phyla in glacier ice reveal unprecedented biodiversity in New Zealand’s Southern Alps. Sci. Rep.-UK 2021, 11, 3898. [Google Scholar] [CrossRef]
- Docherty, C.L.; Hannah, D.M.; Riis, T.; Lund, M.; Abermann, J.; Milner, A.M. Spatio-temporal dynamics of macroinvertebrate communities in northeast Greenlandic snowmelt streams. Ecohydrology 2018, 11, e1982. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.E.; Khamis, K.; Wilkes, M.; Blaen, P.; Brittain, J.E.; Carrivick, J.L.; Fell, S.; Friberg, N.; Füreder, L.; Gislason, G.M.; et al. Functional diversity and community assembly of river invertebrates show globally consistent responses to decreasing glacier cover. Nat. Ecol. Evol. 2018, 2, 325–333. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin III, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.A.; Dirzo, R.; Huber- Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.X.; Liu, S.L.; Liu, Y.X.; Dong, Y.H.; Li, M.Q.; An, Y.; Shi, F.N.; Beazley, R. Effects of the interaction among climate, terrain and human activities on biodiversity on the Qinghai-Tibet Plateau. Sci. Total Environ. 2021, 794, 148497. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.C.; Loewen, C.J.G.; Vinebrooke, R.D.; Chimimba, C.T. Net effects of multiple stressors in freshwater ecosystems: A meta-analysis. Glob. Chang. Biol. 2016, 22, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Masese, F.O.; Otieno, A. Macroinvertebrate taxa display increased fidelity to preferred biotopes among disturbed sites in a hydrologically variable tropical river. Hydrobiologia 2021, 848, 321–343. [Google Scholar] [CrossRef]
- Martins, I.; Castro, D.M.P.; Macedo, D.R.; Hughes, R.M.; Callisto, M. Anthropogenic impacts influence the functional traits of Chironomidae (Diptera) assemblages in a neotropical savanna river basin. Aquat. Ecol. 2021, 55, 1081–1095. [Google Scholar] [CrossRef]
- Hughes, R.M.; Vadas, R.L., Jr. Agricultural Effects on Streams and Rivers: A Western USA Focus. Water 2021, 13, 1901. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef]
- Covich, A.P.; Palmer, M.A.; Crowl, T.A. The role of benthic invertebrate species in freshwater ecosystems. Bioscience 1999, 49, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Chessman, B.C.; Royal, M.J. Bioassessment without reference sites: Use of environmental filters to predict natural assemblages of river macroinvertebrates. J. N. Am. Benthol. Soc. 2003, 23, 599–615. [Google Scholar] [CrossRef]
- Resh, V.H.; Rosenberg, D.M. Recent trends in life-history research on benthic macroinvertebrates. J. N. Am. Benthol. Soc. 2010, 29, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Liu, J.; Zhou, J.; Xia, Y.; Liu, L. Composition of macrozoobenthos functional feeding groups and assessment of ecological quality using the feeding evenness index calculated from biomass data. Biodivers. Sci. 2016, 24, 1045–1055. [Google Scholar] [CrossRef]
- Firmiano, K.R.; Castro, D.M.P.; Linares, M.S.; Callisto, M. Functional responses of aquatic invertebrates to anthropogenic stressors in riparian zones of neotropical savanna streams. Sci. Total Environ. 2021, 753, 141865. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.X.; Cai, Q.H.; Tang, T.; Qu, X.D. The functional feeding group ecology of macroinvertebrate in Xiangxi River system. Acta Ecol. Sin. 2009, 29, 5207–5218. [Google Scholar] [CrossRef]
- Ferreira, W.R.; Ligeiro, R.; Macedo, D.R.; Hughes, R.M.; Kaufmann, P.R.; Oliveira, L.G.; Callisto, M. Is the diet of a typical shredder related to the physical habitat of headwater streams in the Brazilian Cerrado? Ann. Limnol.-Int. J. Limnol. 2015, 51, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet Frédéric Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Zhu, P.; Pan, B.; Li, Z.; Yuan, J.; Jiang, X.; Jiang, W.; Lu, Y. Responses of macroinvertebrate functional traits to riverbed structure of typical debris flow gullies in the upper reaches of the Yangtze River, China. Sci. Total Environ. 2020, 738, 138906. [Google Scholar] [CrossRef]
- Chessman, B.C. Relationships between lotic macroinvertebrate traits and responses to extreme drought. Freshw. Biol. 2015, 60, 50–63. [Google Scholar] [CrossRef]
- Murphy, P.N.C.; Mellander, P.E.; Melland, A.R.; Buckley, C.; Shore, M.; Shortle, G.; Wall, D.P.; Treacy, M.; Shine, O.; Mechan, S.; et al. Variable response to phosphorus mitigation measures across the nutrient transfer continuum in a dairy grassland catchment. Agric. Ecosyst. Environ. 2015, 207, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Fulgoni, J.N.; Whiles, M.R.; Dodds, W.K.; Larson, D.M.; Jackson, K.E.; Grudzinski, B.P. Responses and resilience of tallgrass prairie streams to patch-burn grazing. J. Appl. Ecol. 2020, 57, 1030–1313. [Google Scholar] [CrossRef]
- Cavaillé, P.; Dumont, B.; Van Looy, K.; Floury, M.; Tabacchi, E.; Evette, A. Influence of riverbank stabilization techniques on taxonomic and functional microbenthic communities. Hydrobiologia 2018, 807, 19–35. [Google Scholar] [CrossRef]
- González-Trujillo, J.D.; Petsch, D.K.; Córdoba-Ariza, G.; Rincón-Palau, K.; Donato-Rondon, J.C.; Castro-Rebolledo, M.I.; Sabater, S. Upstream refugia and dispersal ability may override benthic-community responses to high-Andean streams deforestation. Biodivers. Conserv. 2019, 28, 1513–1531. [Google Scholar] [CrossRef]
- Cantonati, M.; Gerecke, R.; Bertuzzi, E. Springs of the Alps-sensitive ecosystems to environmental change: From biodiversity assessments to long-term studies. Hydrobiologia 2006, 562, 59–96. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Resh, V.H. Distribution of benthic algae and macroinvertebrates along a thermal stream gradient. Hydrobiologia 1985, 128, 13–21. [Google Scholar] [CrossRef]
- Vilane, B.R.T.; Dlamini, J. An assessment of the impacts of hot spring usage on water quality in Swaziland. J. Agric. Sci. Technol. 2016, 2, 40–45. [Google Scholar]
- Holt, R.F. Special elements of biodiversity in British Columbia. British Colombia, 2007, 1–30. Available online: http://www.biodiversitybc.org/assets/Default/BBC%20Special%20Elements.pdf (accessed on 10 February 2022).
- Byrne, P.; Wood, P.J.; Reid, I. The impairment of river systems by metal mine contamination: A review including remediation options. Crit. Rev. Environ. Sci. Technol. 2017, 42, 2017–2077. [Google Scholar] [CrossRef]
- Wagener, S.M.; LaPerriere, J.D. Effects of placer mining on the invertebrate communities of interior Alaska, USA, streams. Freshw. Sci. 1985, 4, 208–214. [Google Scholar] [CrossRef]
- Stubblefield, A.; Chandra, S.; Eagan, D.; Tuvshinjargal, G.; Davaadorzh, D.; Gilroy, J.; Sampson, J.; Thorne, B.; Allen, Z.; Hogan, Z. Impacts of gold mining and land use alterations on the water quality of central Mongolian rivers. Integr. Environ. Assess.Manag. Int. J. 2005, 4, 365–373. [Google Scholar] [CrossRef]
- Inam, E.; Khantotong, S.; Kim, K.; Tumendemberel, B.; Erdenetsetseg, S.; Puntsag, T. Geochemical distribution of trace element concentrations in the vicinity of Boroo Gold Mine, Mongolia. Environ. Geochem. Health 2011, 33, 57–59. [Google Scholar] [CrossRef]
- Wood, P.J.; Armitage, P.D. Biological effects of fine sediment in the lotic environment. Environ. Manag. 1997, 21, 203–217. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2010, 81, 117–142. [Google Scholar] [CrossRef]
- Kimmel, W.G.; Argent, D.G. Impacts of point-source Net Alkaline Mine Drainage (NAMD) on stream macroinvertebrate communities. J. Environ. Manag. 2019, 250, 109484. [Google Scholar] [CrossRef] [PubMed]
- Musonge, P.S.L.; Boets, P.; Lock, K.; Goethals, P.L.M. Drivers of Benthic Macroinvertebrate Assemblages in Equatorial Alpine Rivers of the Rwenzoris (Uganda). Water 2020, 12, 1668. [Google Scholar] [CrossRef]
- Araújo-Flores, J.M.; Garate-Quispe, J.; García Molinos, J.; Pillaca-Ortiz, J.M.; Caballero-Espejo, J.; Ascorra, C.; Silman, M.; Fernandez, L.E. Seasonality and aquatic metacommunity assemblage in three abandoned gold mining ponds in the southwestern Amazon, Madre de Dios (Peru). Ecol. Indic. 2021, 125, 107455. [Google Scholar] [CrossRef]
- Kotalik, C.J.; Cadmus, P.; Clements, W.H. Before-After Control-Impact field surveys and novel experimental approaches provide valuable insights for characterizing stream recovery from acid mine drainage. Sci. Total Environ. 2021, 771, 145419. [Google Scholar] [CrossRef]
- Liu, S.Y.; Guo, W.Q.; Xu, J.L. The Second Glacier Inventory Dataset of China (Version 1.0) (2006–2011); National Tibetan Plateau Data Center: Beijing, China, 2012. [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- Moretti, M.; Dias, A.T.C.; Bello, D.F.; Altermatt, F.; Chown, S.L.; Azcárate, F.M.; Bell, J.R.; Fournier, B.; Hedde, M.; Hortal, J.; et al. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Funct. Ecol. 2017, 31, 558–567. [Google Scholar] [CrossRef]
- Cummins, K.W. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic waters. Am. Midl. Nat. 1962, 67, 477–504. [Google Scholar] [CrossRef]
- Jones, C.S.; Drake, C.W.; Hruby, C.E.; Schilling, K.E.; Wolter, C.F. Livestock manure driving stream nitrate. Ambio 2019, 48, 1143–1153. [Google Scholar] [CrossRef]
- Wang, L.; Gao, Y.Z.; Han, B.P.; Fan, H.J.; Yang, H.J. The impacts of agriculture on macroinvertebrate communities: From structural changes to functional changes in Asia’s cold region streams. Sci. Total Environ. 2019, 676, 155–164. [Google Scholar] [CrossRef]
- Rios, S.L.; Bailey, R.C. Relationship between riparian vegetation and stream benthic communities at three spatial scales. Hydrobiologia 2006, 553, 153–160. [Google Scholar] [CrossRef]
- Bottazzi, E.; Bruno, E.M.C.; Pieri, V.; Di Sabatino, A.; Silveri, L.; Carolli, M.; Rosetti, G. Spatial and seasonal distribution of invertebrates in Northern Apennine rheocrene springs. J. Limnol. 2011, 70, 77–92. [Google Scholar] [CrossRef]
- Gathmann, F.O.; Manne, L.L.; Williams, D.D. Spatial patterns in insect community composition in coldwater springs. Aquat. Ecol. 2009, 43, 501–512. [Google Scholar] [CrossRef]
- Jong, G.D.D.; Canton, S.P.; Chadwick, J.W. Macroinvertebrates Occurring in Sunbeam Hot Springs, an Absolutely Hot Spring in Idaho, USA. J. Freshw. Ecol. 2005, 20, 611–613. [Google Scholar] [CrossRef]
- Bekele, T.G.; Ambelu, A.; Chegen, R.G.; Bekele, A.G.; Mekonen, S.; Endale, H. Relevance of macroinvertebrate communities as a water quality monitoring tool in ecosystems under harsh environmental conditions in the Rift Valley region. Environ. Monit. Assess. 2021, 193, 138. [Google Scholar] [CrossRef]
- Wigger, F.W.; Schmidlin, L.; Nagel, P.; Fumetti, S.V. Macroinvertebrate assemblages of natural springs along an altitudinal gradient in the Bernese Alps, Switzerland. Ann. Limnol.-Int. J. Limnol. 2015, 51, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Dethier, E.; Sartain, S.L.; Lutz, D.A. Heightened levels and seasonal inversion of riverine suspended sediment in a tropical biodiversity hot spot due to artisanal gold mining. Proc. Natl. Acad. Sci. USA 2019, 116, 23936–23941. [Google Scholar] [CrossRef]
- Drover, D.R.; Schoenholtz, S.H.; Soucek, D.J.; Zipper, C.E. Multiple stressors influence benthic macroinvertebrate communities in central Appalachian coalfield streams. Hydrobiologia 2020, 847, 191–205. [Google Scholar] [CrossRef]
- Encalada, A.C.; Calles, J.; Ferreira, V.; Canhoto, C.M.; Graça, M.A.S.; Graca, M.A.S. Riparian land use and the relationship between the benthos and litter decomposition in tropical montane streams. Freshw. Biol. 2010, 55, 1719–1733. [Google Scholar] [CrossRef]
- Lorion, C.M.; Kennedy, B.P. Relationships between deforestation, riparian forest buffers and benthic macroinvertebrates in neotropical headwater streams. Freshw. Biol. 2009, 54, 165–180. [Google Scholar] [CrossRef]
- Espinosa, R.; Andino, P.; Cauvy-Fraunié, S.; Dangles, O.; Jacobsen, D.; Crespo-Pérez, V. Diversity patterns of aquatic macroinvertebrates in a tropical high-Andean catchment. Rev. Biol. Trop. 2020, 68, S29–S53. [Google Scholar] [CrossRef]
- Duan, X.; Wang, Z.; Xu, M. Benthic Macroinvertebrate and Application in the Assessment of Stream Ecology; Tsinghua University Publishing House: Beijing, China, 2010; pp. 30–31. [Google Scholar]
- Torres-Olvera, M.J.; Durán-Rodríguez, O.Y.; Torres-García, U.; Pineda-López, R.; Ramírez- Herrejón, J.P. Validation of an index of biological integrity based on aquatic macroinvertebrates assemblages in two subtropical basins of Central Mexico. Lat. Am. J. Aquat. Res. 2018, 46, 945–960. [Google Scholar] [CrossRef]
- Weir, S.M.; Salice, C.J. High tolerance to abiotic stressors and invasion success of the slow growing freshwater snail, Melanoides tuberculatus. Biol. Invasions 2012, 14, 385–394. [Google Scholar] [CrossRef]
- Egler, M.; Buss, D.F.; Moreira, J.C.; Baptista, D.F. Influence of agricultural land-use and pesticides on benthic macroinvertebrate assemblages in an agricultural river basin in southeast Brazil. Braz. J. Biol. 2012, 72, 437–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.E.; Hannah, D.M.; Milner, A.M. Vulnerability of alpine stream biodiversity to shrinking glaciers and snowpacks. Glob. Chang. Biol. 2007, 13, 958–966. [Google Scholar] [CrossRef]
- Jacobsen, D. Altitudinal changes in diversity of macroinvertebrates from small streams in the Ecuadorian Andes. Arch. Hydrobiol. 2003, 158, 145–167. [Google Scholar] [CrossRef]
- Derso, S.; Beyene, A.; Melaku, G.; Ambelu, A. Ecological status of hot springs in Eastern Amhara Region: Macroinvertebrates diversity. Am. Acad. Sci. Res. J. Eng.Technol. Sci. 2015, 14, 1–22. [Google Scholar]
- Gerecke, R.; Di Sabatino, A. Water mites (Acari, Hydrachnellae) and spring typolog in Sicily. Crunoecia 1996, 5, 35–42. [Google Scholar]
- Heishman, A.; Mcluky, R.G. Use of Conductivity to Define Compliance with State Narrative Water Quality Standard; Energy & Mineral. Law Institute: Amelia Island, FL, USA, 2011. [Google Scholar]
- Arkle, R.S.; Pilliod, D.S. Stream Restoration Is Influenced by Details of Engineered Habitats at a Headwater Mine Site. Diversity 2021, 13, 48. [Google Scholar] [CrossRef]
- Herbst, D.B.; Medhurst, R.B.; Black, N.J.P. Long-term effects and recovery of streams from acid mine drainage and evaluation of toxic metal threshold ranges for macroinvertebrate community reassembly. Environ. Toxicol. Chem. 2018, 37, 2575–2592. [Google Scholar] [CrossRef]
- Lake, P.S.; Bond, N.; Reich, P. Linking ecological theory with stream restoration. Freshw. Biol. 2007, 52, 597–615. [Google Scholar] [CrossRef]
- Winking, C.; Lorenz, A.W.; Sures, B.; Hering, D. Recolonization patterns of benthic invertebrates: A field investigation of restored former sewage channels. Freshw. Biol. 2014, 59, 1932–1944. [Google Scholar] [CrossRef]
- Damme, P.A.V.; Hamel, C.; Ayala, A.; Bervoets, L. Macroinvertebrate community response to acid mine drainage in rivers of the High Andes (Bolivia). Environ. Pollut. 2008, 156, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.; Baird, D.J.; Soares, A. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Saito, V.S.; Soininen, J.; Fonseca-Gessner, A.A.; Siqueira, T. Dispersal traits drive the phylogenetic distance decay of similarity in Neotropical stream metacommunities. J. Biogeogr. 2015, 42, 2101–2111. [Google Scholar] [CrossRef]
- Brederveld, R.J.; Jähnig, S.C.; Lorenz, A.W.; Brunzel, S.; Soons, M.B. Dispersal as a limiting factor in the colonization of restored mountain streams by plants and macroinvertebrates. J. Appl. Ecol. 2011, 48, 1241–1250. [Google Scholar] [CrossRef]
- Füreder, L. Life at the edge: Habitat condition and bottom fauna of alpine running waters. Int. Rev. Hydrobiol. 2007, 92, 491–513. [Google Scholar] [CrossRef]
- Cauvy-Fraunié, S.; Andino, P.; Espinosa, R.; Calvez, R.; Anthelme, F.; Jacobsen, D.; Dangles, O. Glacial flood pulse effects on benthic fauna in equatorial high-Andean streams. Hydrol. Processes 2013, 28, 3008–3017. [Google Scholar] [CrossRef]
- Bêche, L.A.; Mcelravy, E.P.; Resh, V.H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, U.S.A. Freshw. Biol. 2006, 51, 56–75. [Google Scholar] [CrossRef]
- Milner, A.M.; Brittain, J.E.; Castella, E.; Petts, G.E. Trends of macroinvertebrate community structure in glacier-fed rivers in relation to environmental conditions: A synthesis. Freshw. Biol. 2001, 46, 1833–1847. [Google Scholar] [CrossRef]




| Functional Traits | Trait State | Code |
|---|---|---|
| Life history | ||
| Voltinism | Semivoltine | Volt 1 |
| Univoltine | Volt 2 | |
| Bi- or- multivoltine | Volt 3 | |
| Mobility | ||
| Occurrence in drift | Rare | Drft 1 |
| Common | Drft 2 | |
| Abundant | Drft 3 | |
| Morphology | ||
| Shape | Streamlined | Shpe 1 |
| Not streamlined | Shpe 2 | |
| Size at maturity | Small (<9 mm) | Size 1 |
| Medium (9–16 mm) | Size 2 | |
| Large (>16 mm) | Size 3 | |
| Ecology | ||
| Habit | Burrower | Habi 1 |
| Climber | Habi 2 | |
| Sprawler | Habi 3 | |
| Clinger | Habi 4 | |
| Swimmer | Habi 5 | |
| Trophic habit | Collector-gatherer | Trop 1 |
| Collector-filterer | Trop 2 | |
| Herbivore | Trop 3 | |
| Predator | Trop 4 | |
| Shredder | Trop 5 |
| Environment Variables | S1 | S2 | S3 |
|---|---|---|---|
| Altitude (m) | 1788.59 ± 234.31 a | 2417.73 ± 141.75 b | 2439.45 ± 292.71 b |
| Velocity (m·s−1) | 0.56 ± 0.05 | 0.60 ± 0.16 | 0.52 ± 0.17 |
| Depth (m) | 0.32 ± 0.06 | 0.36 ± 0.09 | 0.35 ± 0.09 |
| Water temperature (°C) | 7.29 ± 0.70 a | 4.58 ± 0.34 b | 5.10 ± 0.76 b |
| Dissolved oxygen (%) | 93.18 ± 1.64 | 95.38 ± 8.30 | 92.44 ± 6.89 |
| Electrical conductivity (μS·cm−1) | 242.28 ± 46.66 a | 191.08 ± 40.52 b | 243.52 ± 80.92 c |
| pH | 8.22 ± 0.41 | 8.10 ± 0.26 | 8.11 ± 0.41 |
| Turbidity (FNU) | 39.99 ± 5.11 a | 30.52 ± 1.54 b | 38.67 ± 0.74 a |
| Ammonium nitrogen (mg·L−1) | 0.34 ± 0.04 a | 0.25 ± 0.01 b | 0.19 ± 0.01 c |
| Total nitrogen (mg·L−1) | 0.93 ± 0.05 a | 0.81 ± 0.01 b | 0.72 ± 0.01 c |
| Total phosphorus (mg·L−1) | 0.82 ± 0.08 a | 0.64 ± 0.02 b | 0.59 ± 0.01 b |
| Substrate: | |||
| Sands (<2 mm) | 30.0% ± 10.85 a | 17.9% ± 3.64 b | 21.6% ± 5.65 c |
| Gravels (2–64 mm) | 33.1% ± 3.23 | 35.6% ± 3.99 | 36.5% ± 7.01 |
| Cobbles (64–256 mm) | 30.6% ± 5.94 | 37.7% ± 3.9 | 37.5% ± 7.38 |
| Boulders (>256 mm) | 7.4% ± 6.70 | 8.8% ± 5.13 | 4.4% ± 3.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Tian, Y.; Gao, Y.; Cui, D.; Zhang, W.; Jiao, Z.; Yao, F.; Zhang, Z.; Yang, H. The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains. Water 2022, 14, 1298. https://doi.org/10.3390/w14081298
Liu Y, Tian Y, Gao Y, Cui D, Zhang W, Jiao Z, Yao F, Zhang Z, Yang H. The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains. Water. 2022; 14(8):1298. https://doi.org/10.3390/w14081298
Chicago/Turabian StyleLiu, Yuwei, Yilin Tian, Yingzhi Gao, Dong Cui, Wei Zhang, Ziwei Jiao, Fulong Yao, Zhenxing Zhang, and Haijun Yang. 2022. "The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains" Water 14, no. 8: 1298. https://doi.org/10.3390/w14081298
APA StyleLiu, Y., Tian, Y., Gao, Y., Cui, D., Zhang, W., Jiao, Z., Yao, F., Zhang, Z., & Yang, H. (2022). The Impacts of Different Anthropogenic Disturbances on Macroinvertebrate Community Structure and Functional Traits of Glacier-Fed Streams in the Tianshan Mountains. Water, 14(8), 1298. https://doi.org/10.3390/w14081298