
Composition of Colloidal Organic Matter in Phytoplankton Exudates



Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Methods
2.2. Analyses
DOC Analysis
2.3. NMR Spectroscopy
3. Results and Discussion
3.1. COM Concentration
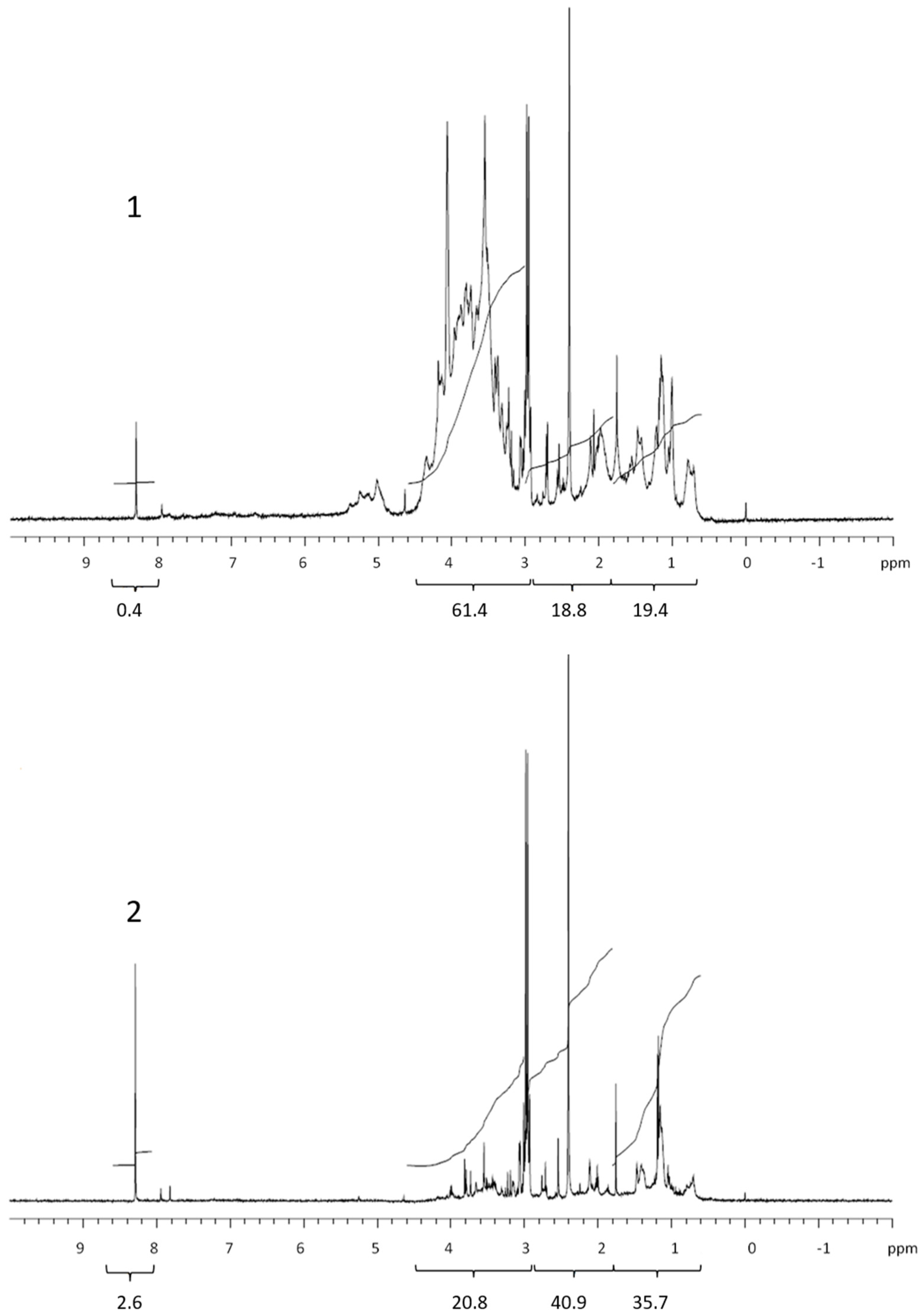
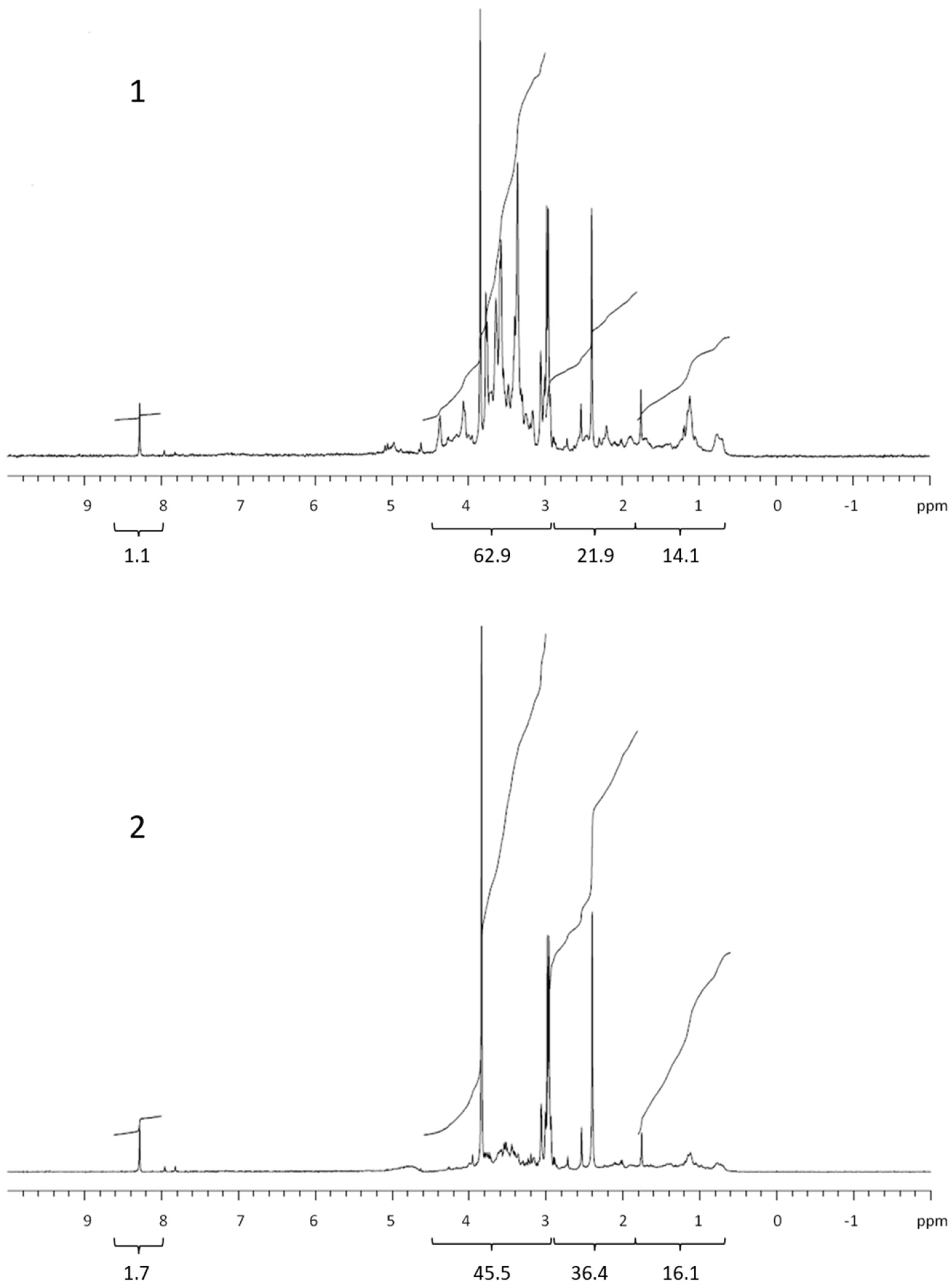
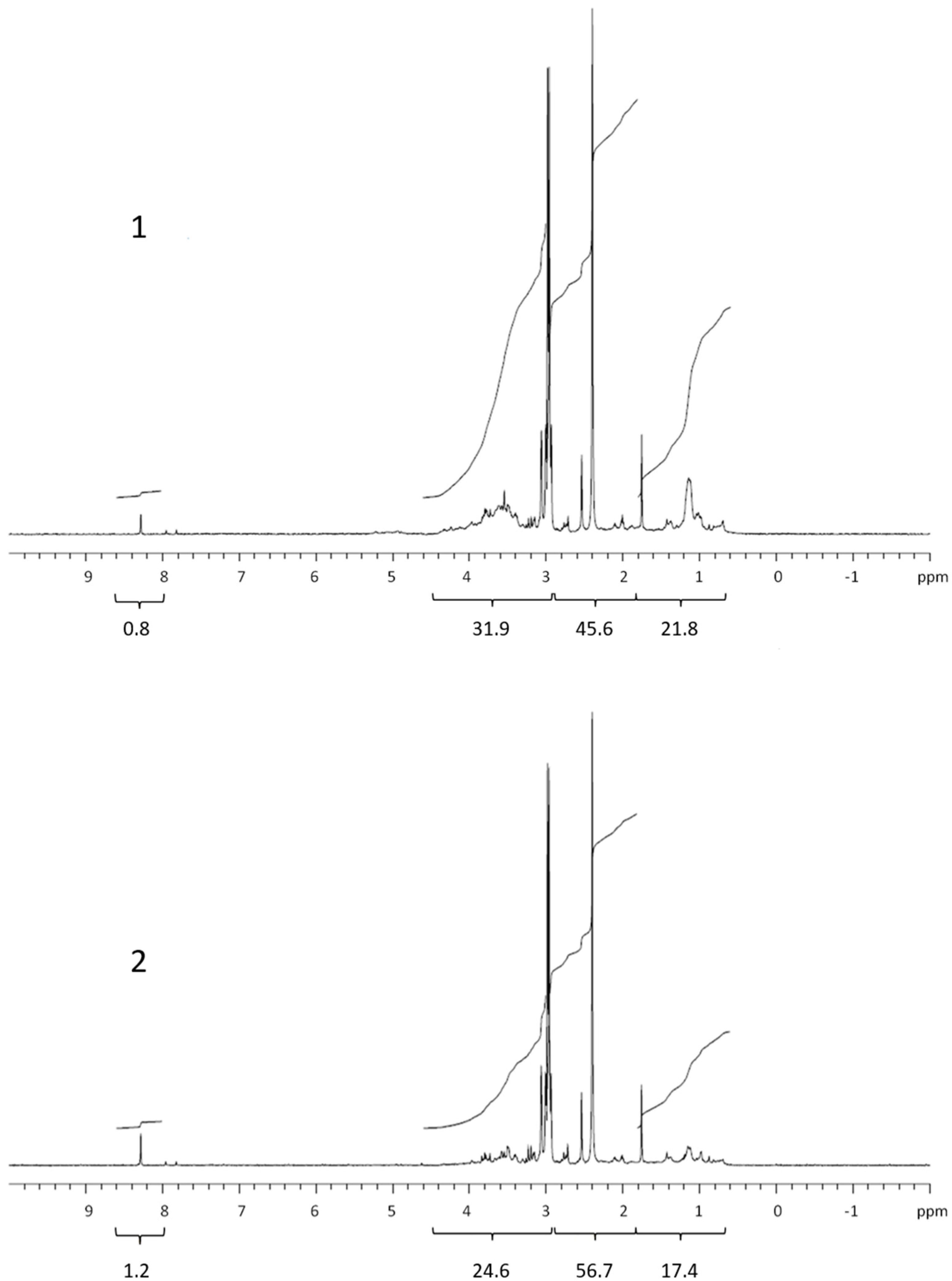
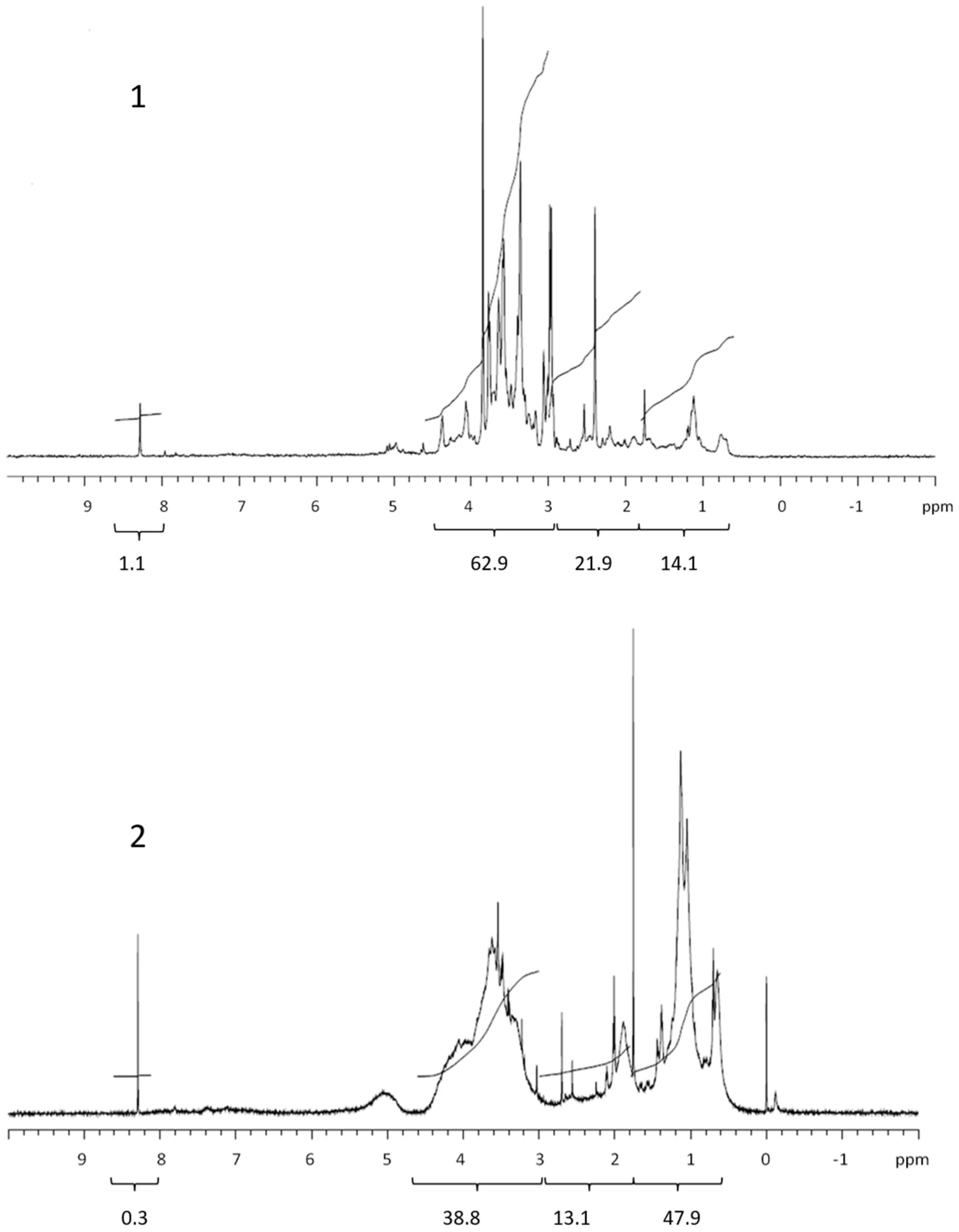
3.2. COM Composition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hedges, J.I. Why dissolved organic matter? In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Academic Press: Boston, MA, USA, 2002; pp. 1–33. [Google Scholar]
- Fillela, M. Environmental colloids and particles (behavior, separation and characterisation). In Environmental Colloids and Particles; Wilkinson, K.J., Lead, J.R., Eds.; John Wiley & Sons: Chichester, UK, 2007; pp. 17–94. [Google Scholar]
- Verdugo, P.; Alldredge, A.L.; Azam, F.; Kirchman, D.L.; Passow, U.; Santschi, P.H. The oceanic gel phase: A bridge in the DOM-POM continuum. Mar. Chem. 2004, 92, 67–85. [Google Scholar] [CrossRef]
- Mopper, K.; Stubbins, A.M.; Ritchie, J.D.; Bialk, H.M.; Hatcher, P.G. Advanced instrumental approaches for characterization of marine dissolved matter: Extraction techniques, mass spectrometry, and nuclear magnetic resonance spectroscopy. Chem. Rev. 2007, 107, 419–442. [Google Scholar]
- Benner, R. Chemical composition and reactivity. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: San Diego, CA, USA, 2002; pp. 59–90. [Google Scholar]
- Moran, M.A.; Kujawinski, E.B.; Schroer, W.F.; Amin, S.A.; Bates, N.R.; Bertrand, E.M.; Braakman, R.; Brown, R.; Covert, C.T.; Doney, S.C.; et al. Microbial metabolites in the marine carbon cycle. Nat. Microbiol. 2022, 7, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Chin, W.C.; Orellana, M.V.; Verdugo, P. Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature 1998, 391, 568–572. [Google Scholar] [CrossRef]
- Verdugo, P.; Santschi, P.H. Polymer dynamics of DOC network and gel formation in seawater. Deep Sea Res. II 2010, 57, 1486–1493. [Google Scholar] [CrossRef]
- Giani, M.; Degobbis, D.; Rinaldi, A. Mucilages in the Adriatic and Tyrrhenian Seas. Sci. Total Environ. 2005, 353, 1–2. [Google Scholar] [CrossRef]
- Lam, B.; Simpson, A.J. Direct 1H NMR spectroscopy of dissolved organic matter in natural waters. Analyst 2008, 133, 263–269. [Google Scholar] [CrossRef]
- Hertkorn, N.; Benner, R.; Frommberger, M.; Schmitt-Kopplin, P.; Witt, M.; Kaiser, K.; Kettrup, A.; Hedges, J.I. Characterizationn of a major refractory component of marine dissolved organic matter. Geochim. Cosmochim. Acta 2006, 70, 2990–3010. [Google Scholar] [CrossRef]
- Hertkorn, N.; Harir, M.; Koch, B.P.; Michalke, B.; Schmitt-Kopplin, P. High-field NMR spectroscopy and FTICR mass spectrometry: Powerful discovery tools for the molecular level characterization of marine dissolved organic matter. Biogeosciences 2013, 10, 1583–1624. [Google Scholar] [CrossRef] [Green Version]
- Klun, K.; Šket, P.; Falnoga, I.; Faganeli, J. Variation of colloidal organic matter composition in coastal waters (Gulf of Trieste, northern Adriatic Sea). Geomicrobiol. J. 2015, 32, 609–615. [Google Scholar] [CrossRef]
- Edison, A.S.; Colonna, M.; Gouvenia, G.J.; Holderman, N.R.; Judge, M.Z.; Shen, X.; Zhang, S. NMR: Unique strengths that enhance modern metabolomics research. Anal. Chem. 2021, 93, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Kovac, N.; Bajt, O.; Faganeli, J.; Sket, B.; Orel, B. Study of macroaggregate composition using FT-IR and 1H-NMR spectroscopy. Mar. Chem. 2002, 78, 205–215. [Google Scholar] [CrossRef]
- Kovac, N.; Faganeli, J.; Bajt, O.; Sket, B.; Orel, B.; Penna, N. Chemical composition of macroaggregates in the northern Adriatic Sea. Org. Geochem. 2004, 35, 1095–1104. [Google Scholar] [CrossRef]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A broad spectrum artificial seawater medium for coastal and open ocean phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar]
- Engel, A.; Handel, N. A novel protocol for determining the concentration and composition of sugars and particulate and high molecular dissolved organic matter (HMW DOM) in sea water. Mar. Chem. 2011, 127, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, Y.; Suzuki, Y. A high temperature catalytic oxidation method for the determination of non-volatile dissolved organic carbon in seawater by direct injection of liquid sample. Mar. Chem. 1988, 24, 105–131. [Google Scholar] [CrossRef]
- Faganeli, J.; Pezdič, J.; Fanuko, N. Chemical and isotopic composition of some phytoplankton species. Biol. Vestn. 1989, 37, 9–20. [Google Scholar]
- Faganeli, J.; Herndl, G.J. Dissolved organic matter in the waters of the Gulf of Trieste (Northern Adriatic). Thalassia Jugosl. 1991, 23, 51–63. [Google Scholar]
- De Vittor, C.; Paoli, A.; Fonda Umani, S. Dissolved organic carbon variability in a shallow coastal marine system (Gulf of Trieste, northern Adriatic Sea). Estuar. Coast. Shelf Sci. 2008, 78, 280–290. [Google Scholar] [CrossRef]
- Fonda Umani, S.; Del Negro, P.; Larato, C.; De Vittor, C.; Cabrini, M.; Celio, M.; Falconi, C.; Tamberich, F.; Azam, F. Major inter-annual variations in microbial dynamics in the Gulf of Trieste (northern Adriatic Sea) and their ecosystem implications. Aquat. Microb. Ecol. 2007, 46, 163–175. [Google Scholar] [CrossRef]
- Penna, N.; Kovac, N.; Ricci, F.; Penna, A.; Capellacci, S.; Faganeli, J. The role of dissolved carbohydrates in the northern Adriatic macroaggregate formation. Acta Chim. Slov. 2009, 56, 305–314. [Google Scholar]
- Aluwihare, L.; Repeta, D.; Chen, R. A major biopolymeric component to dissolved organic carbon in surface sea water. Nature 1997, 387, 166–169. [Google Scholar] [CrossRef]
- Lam, B.; Baer, A.; Alaee, M.; Lefebvre, B.; Moser, A.; Williams, A.; Simpson, A.J. Major structural components in freshwater dissolved organic matter. Environ. Sci. Technol. 2007, 41, 8240–8246. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, K.P.; Cave, E.R.; Abram, D.N.; Jaramillo, T.F. New insights into the electrochemical reduction of carbon dioxide on metallic copper surfaces. Energy Environ. Sci 2012, 5, 7050–7059. [Google Scholar] [CrossRef]
- Becker, J.W.; Berube, P.M.; Follett, C.L.; Waterbury, J.B.; Chisholm, S.W.; DeLong, E.F.; Repeta, D.J. Closely related phytoplankton species produce similar suites of dissolved organic matter. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Biersmith, A.; Benner, R. Carbohydrates in phytoplankton and freshly produced dissolved organic matter. Mar. Chem. 1998, 63, 131–144. [Google Scholar] [CrossRef]
- Repeta, D.J. Chemical characterisation and cycling of dissolved organic matter. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Academic Press: London, UK, 2015; pp. 21–63. [Google Scholar]
- Mozetič, P.; France, J.; Kogovšek, T.; Talaber, I.; Malej, A. Plankton trends and community changes in a coastal sea (northern Adriatic): Bottom-up vs. top-down control on relation to environmental drivers. Estuar. Coast. Shelf Sci. 2012, 115, 138–148. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipids | Proteins and CRAM | Polysaccharides | Formate | Corg. | % COC * | |
|---|---|---|---|---|---|---|
| HCH2-CH2- | HC-HCOR | HC-OH HC-O-C | HCOO | |||
| δ **/ppm | 0–1.8 | 1.8–3.0 | 3.0–4.6 | 8.0–9.0 | µmol L−1 | % |
| Teratselmis marina | ||||||
| 0.2 µm filtrate | 915.1 | |||||
| Retentate | 19.4 | 18.8 | 61.4 | 0.4 | 364 | 39.8 |
| Permeate | 35.7 | 40.9 | 20.8 | 2.6 | 507 | |
| Chaetoceros socialis | ||||||
| 0.2 µm filtrate | 2285 | |||||
| Retentate | 14.4 | 21.9 | 62.9 | 1.1 | 526 | 23.0 |
| Permeate | 16.4 | 36.4 | 45.5 | 1.7 | 1765 | |
| Prorocentrum minimum | ||||||
| 0.2 µm filtrate | 439.3 | |||||
| Retentate | 21.8 | 45.6 | 31.9 | 0,8 | 154 | 35.1 |
| Permeate | 17.4 | 56.7 | 24.6 | 1.2 | 418 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klun, K.; Šket, P.; Beran, A.; Falnoga, I.; Faganeli, J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water 2023, 15, 111. https://doi.org/10.3390/w15010111
Klun K, Šket P, Beran A, Falnoga I, Faganeli J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water. 2023; 15(1):111. https://doi.org/10.3390/w15010111
Chicago/Turabian StyleKlun, Katja, Primož Šket, Alfred Beran, Ingrid Falnoga, and Jadran Faganeli. 2023. "Composition of Colloidal Organic Matter in Phytoplankton Exudates" Water 15, no. 1: 111. https://doi.org/10.3390/w15010111
APA StyleKlun, K., Šket, P., Beran, A., Falnoga, I., & Faganeli, J. (2023). Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water, 15(1), 111. https://doi.org/10.3390/w15010111