
Carbon Dioxide Concentration and Emissions along a Trophic Gradient in Tropical Karst Lakes

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
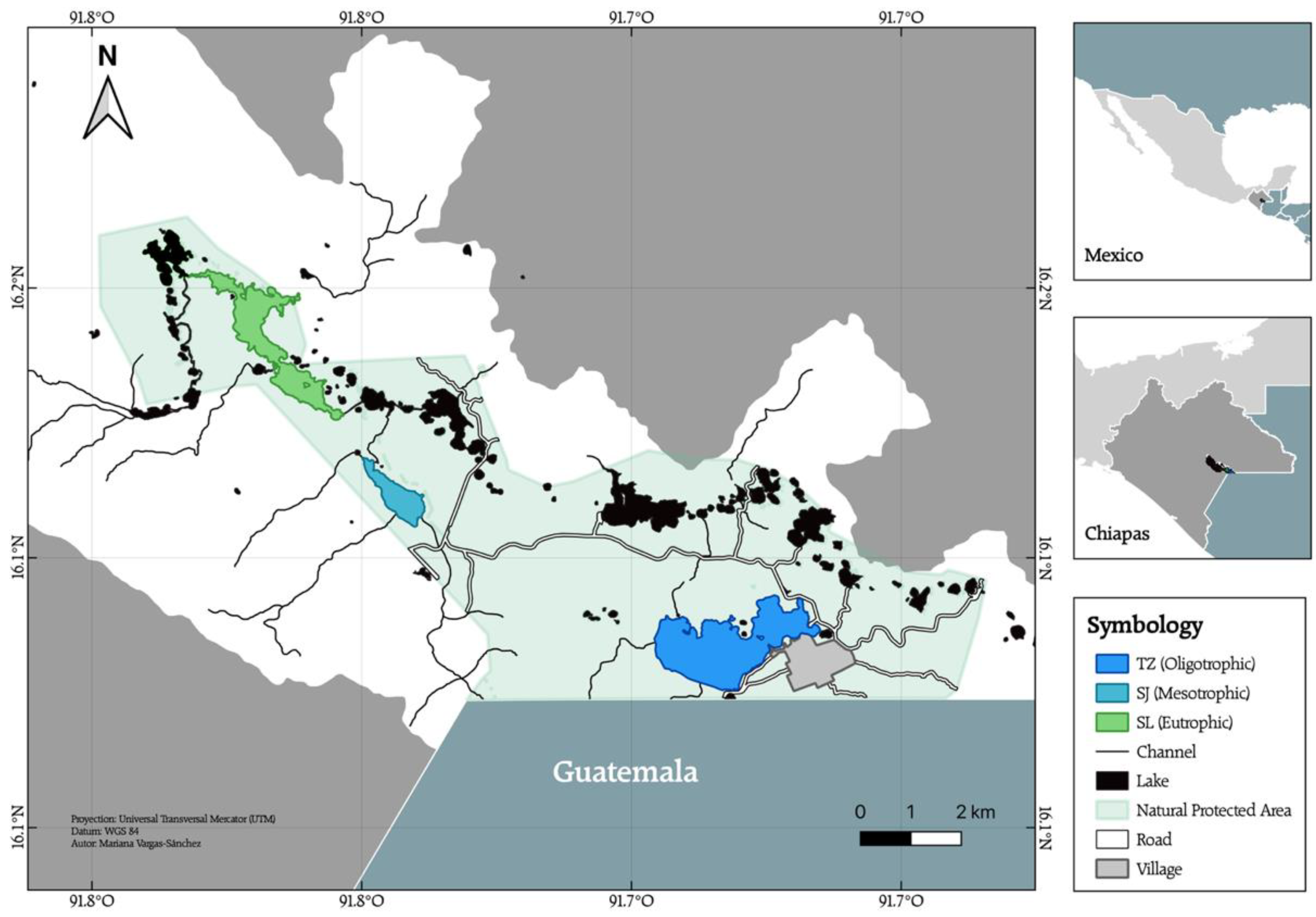
2.1. Study Area
2.2. Physico-Chemical and Biological Characterization
2.3. DIC, Dissolved CO2, and Flux Measurements
2.4. Statistical Analyses
3. Results
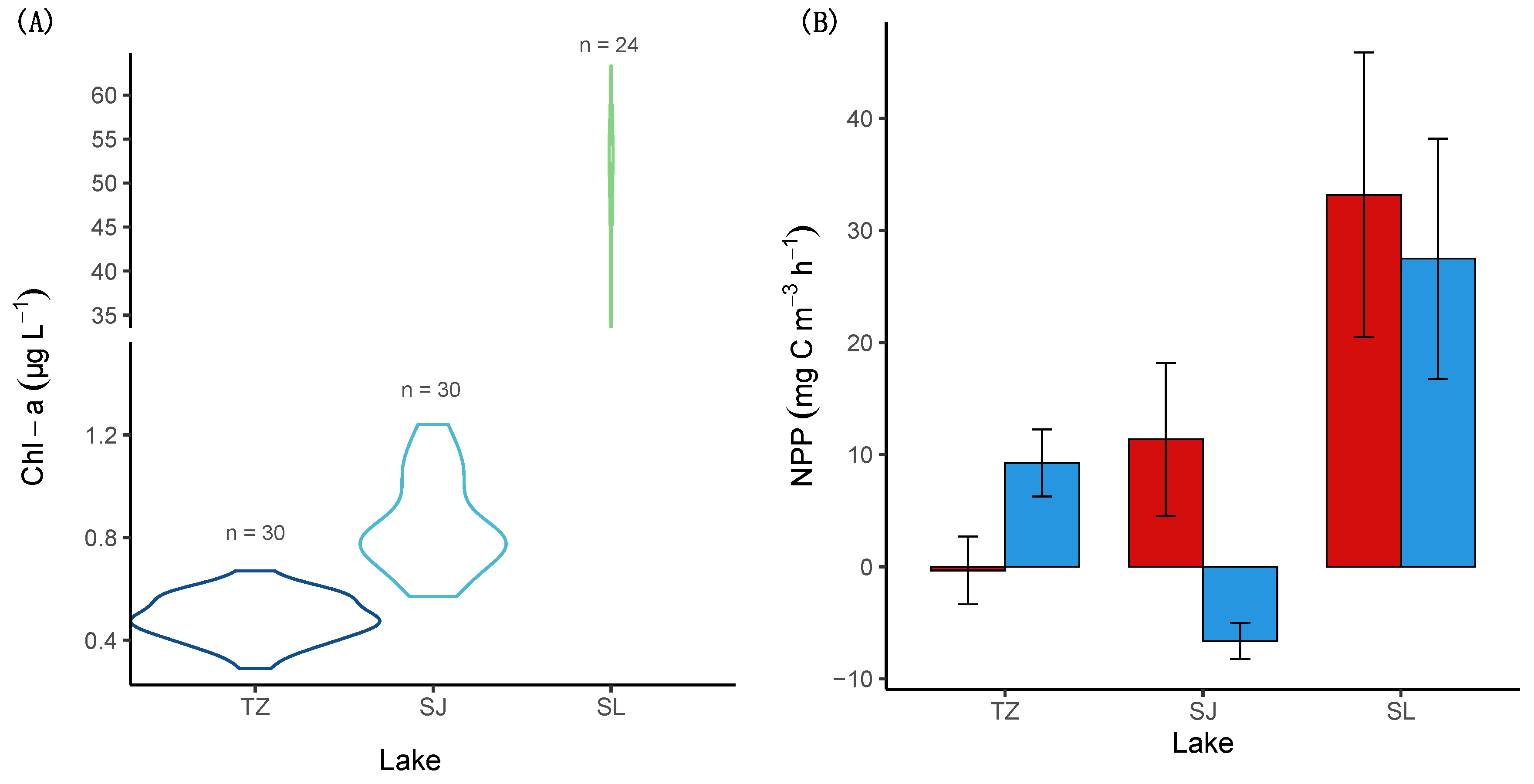
3.1. Physico-Chemical and Biological Characterization
3.2. Dissolved CO2 and DIC
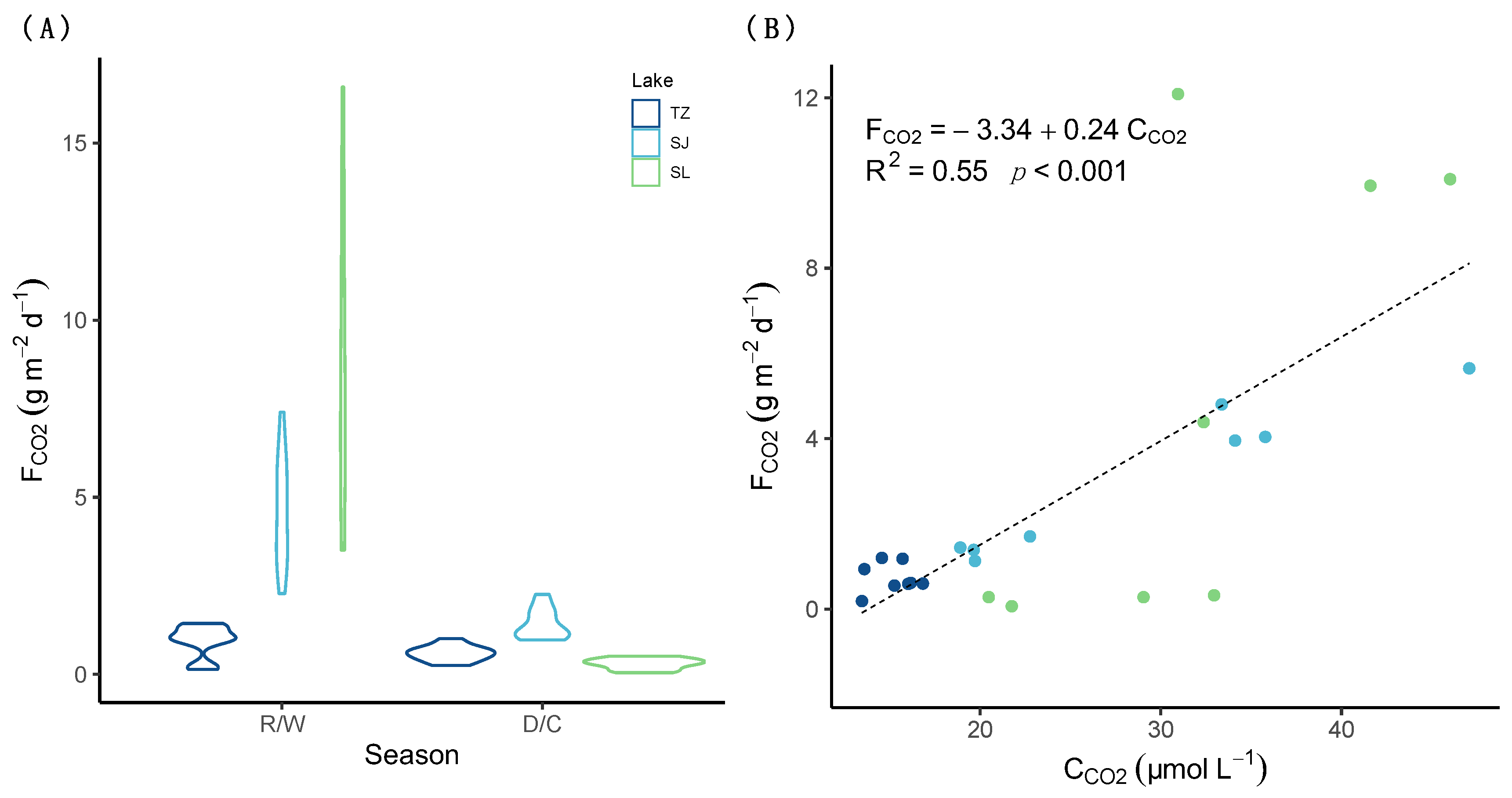
3.3. CO2 Evasion Rates
4. Discussion
4.1. Variation of CCO2 in the Water Column: Seasonality and Main Drivers
4.2. CO2 Evasion from Tropical Karst Lakes: Eutrophication Effects
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, J.J.; Prairie, Y.T.; Caraco, N.F.; McDowell, W.H.; Tranvik, L.J.; Striegl, R.G.; Duarte, C.M.; Kortelainen, P.; Downing, J.A.; Middelburg, J.J.; et al. Plumbing the Global Carbon Cycle: Integrating Inland Waters into the Terrestrial Carbon Budget. Ecosystems 2007, 10, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Battin, T.J.; Kaplan, L.A.; Findlay, S.; Hopkinson, C.S.; Marti, E.; Packman, A.I.; Newbold, J.D.; Sabater, F. Erratum: Biophysical controls on organic carbon fluxes in fluvial networks. Nat. Geosci. 2009, 2, 595. [Google Scholar] [CrossRef]
- Tranvik, L.J.; Downing, J.A.; Cotner, J.B.; Loiselle, S.A.; Striegl, R.G.; Ballatore, T.J.; Dillon, P.; Finlay, K.; Fortino, K.; Knoll, L.B.; et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr. 2009, 54 Pt 2, 2298–2314. [Google Scholar] [CrossRef] [Green Version]
- Butman, D.; Stackpoole, S.; Stets, E.; McDonald, C.P.; Clow, D.W.; Striegl, R.G. Aquatic carbon cycling in the conterminous United States and implications for terrestrial carbon accounting. Proc. Natl. Acad. Sci. USA 2015, 113, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarbuck, E.J.; Lutgens, F.K.; Tasa, D.G. Earth: An Introduction to Physical Geology, 11th ed.; Pearson: London, UK, 2013. [Google Scholar]
- Liu, K.; Atkinson, L.; Quiñones, R.; Talaue-McManus, L.; Mackenzie, F. Carbon and nutrient fluxes in continental margins: A global synthesis. Springer: Berlin/Heidelberg, Germany, 2009. Available online: http://ci.nii.ac.jp/ncid/BB02062305 (accessed on 3 October 2022).
- Prairie, Y.T.; Cole, J.J. The Carbon Cycle in Lakes: A Biogeochemical Perspective. Encycl. Inland Waters 2022, 2, 89–101. [Google Scholar] [CrossRef]
- DelSontro, T.; Beaulieu, J.J.; Downing, J.A. Greenhouse gas emissions from lakes and impoundments: Upscaling in the face of global change. Limnol. Oceanogr. Lett. 2018, 3, 64–75. [Google Scholar] [CrossRef]
- Li, Y.; Shang, J.; Zhang, C.; Zhang, W.; Niu, L.; Wang, L.; Zhang, H. The role of freshwater eutrophication in greenhouse gas emissions: A review. Sci. Total Environ. 2021, 768, 144582. [Google Scholar] [CrossRef] [PubMed]
- Drake, T.W.; Raymond, P.A.; Spencer, R.G.M. Terrestrial carbon inputs to inland waters: A current synthesis of estimates and uncertainty. Limnol. Oceanogr. Lett. 2017, 3, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Pollard, P.C. Globally, Freshwater Ecosystems Emit More CO2 Than the Burning of Fossil Fuels. Front. Environ. Sci. 2022, 10, 904955. [Google Scholar] [CrossRef]
- Sánchez-Carrillo, S.; Alcocer, J.; Vargas-Sánchez, M.; Soria-Reinoso, I.; Rivera-Herrera, E.M.; Cortés-Guzmán, D.; Cuevas-Lara, D.; Guzmán-Arias, A.P.; Merino-Ibarra, M.; Oseguera, L.A. Greenhouse gas emissions from Mexican inland waters: First estimation and uncertainty using an upscaling approach. Inland Waters 2022, 12, 294–310. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; DelSontro, T.; Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nat. Commun. 2019, 10, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatvani, I.G.; Dokulil, M.T.; Clement, A. The role of wetlands in mitigating impacts from diffuse agricultural loads. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 285–299. [Google Scholar]
- Rosseau, D.P.L.; Louage, F.; Wang, Q.; Zhang, R. Constructed wetlands for urban wastewater treatment: An overview. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 272–282. [Google Scholar]
- Deemer, B.R.; Harrison, J.A.; Li, S.; Beaulieu, J.J.; DelSontro, T.; Barros, N.; Bezerra-Neto, J.F.; Powers, S.M.; dos Santos, M.A.; Vonk, J.A. Greenhouse Gas Emissions from Reservoir Water Surfaces: A New Global Synthesis. BioScience 2016, 66, 949–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttunen, J.T.; Alm, J.; Liikanen, A.; Juutinen, S.; Larmola, T.; Hammar, T.; Silvola, J.; Martikainen, P.J. Fluxes of methane, carbon dioxide and nitrous oxide in boreal lakes and potential anthropogenic effects on the aquatic greenhouse gas emissions. Chemosphere 2003, 52, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Beck, B.F. Engineering Issues on Karst. In Karst Management; Springer: Dordrecht, The Netherlands, 2011; pp. 9–45. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, L.; Zhou, Y.; Mao, X.Z. Eutrophication control strategies for highly anthropogenic influenced coastal waters. Sci. Total Environ. 2020, 705, 135760. [Google Scholar] [CrossRef] [PubMed]
- Balmer, M.; Downing, J. Carbon dioxide concentrations in eutrophic lakes: Undersaturation implies atmospheric uptake. Inland Waters 2011, 1, 125–132. [Google Scholar] [CrossRef]
- Gu, B.; Schelske, C.L.; Coveney, M.F. Low carbon dioxide partial pressure in a productive subtropical lake. Aquat. Sci. 2011, 73, 317–330. [Google Scholar] [CrossRef]
- Pacheco, F.; Roland, F.; Downing, J. Eutrophication reverses whole-lake carbon budgets. Inland Waters 2014, 4, 41–48. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Stief, P.; Brandes, J.; Schink, B. Anaerobic methane oxidation coupled to denitrification is the dominant methane sink in a deep lake. Proc. Natl. Acad. Sci. USA 2014, 111, 18273–18278. [Google Scholar] [CrossRef] [Green Version]
- Marcé, R.; Obrador, B.; Morguí, J.A.; Lluís Riera, J.; López, P.; Armengol, J. Carbonate weathering as a driver of CO2 supersaturation in lakes. Nat. Geosci. 2015, 8, 107–111. [Google Scholar] [CrossRef]
- Ford, D.; Williams, P.D. Karst Hydrogeology and Geomorphology (Revised); Wiley: Hoboken, NJ, USA, 2007. [Google Scholar]
- Winter, T.C. Hydrological processes and the water budget of lakes. In Physics and Chemistry of Lakes; Lerman, A., Imboden, D.M., Gat, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 37–62. [Google Scholar]
- Cole, J.J.; Prairie, Y. Dissolved CO2. In Biogeochemistry of Inland Waters; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2010; pp. 343–347. [Google Scholar]
- He, H.; Wang, Y.; Liu, Z.; Bao, Q.; Wei, Y.; Chen, C.; Sun, H. Lake metabolic processes and their effects on the carbonate weathering CO2 sink: Insights from diel variations in the hydrochemistry of a typical karst lake in SW China. Water Res. 2022, 222, 118907. [Google Scholar] [CrossRef]
- Wang, F.; Cao, M.; Wang, B.; Fu, J.; Luo, W.; Ma, J. Seasonal variation of CO2 diffusion flux from a large subtropical reservoir in East China. Atmos. Environ. 2015, 103, 129–137. [Google Scholar] [CrossRef]
- Li, J.; Pu, J.; Zhang, T.; Huang, S.; Yuan, D. Seasonal variations and intricate diel differences in the physio-chemical parameters and CO2 emissions from a typical karst groundwater-fed reservoir in southern China. Environ. Earth Sci. 2019, 78, 484. [Google Scholar] [CrossRef]
- Ni, M.; Ge, Q.; Li, S.; Wang, Z.; Wu, Y. Trophic state index linked to partial pressure of aquatic carbon dioxide in a typical karst plateau lake. Ecol. Indic. 2021, 120, 106912. [Google Scholar] [CrossRef]
- Mora, P.L.; Bonifaz, R.; López-Martínez, R. Unidades geomorfológicas de la cuenca del Río Grande de Comitán, Lagos de Montebello, Chiapas-México. Boletín De La Soc. Geológica Mex. 2016, 68, 377–394. [Google Scholar] [CrossRef]
- Alcocer, J.; Oseguera, L.A.; Sánchez, G.; González, C.G.; Martínez, J.R.; González, R. Bathymetric and morphometric surveys of the Montebello Lakes, Chiapas. J. Limnol. 2016, 75, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Alcocer, J.; Merino-Ibarra, M.; Oseguera, L.A.; Escolero, O. Anthropogenic impacts on tropical karst lakes: “Lagunas de Montebello,” Chiapas. Ecohydrology 2018, 11, e2029. [Google Scholar] [CrossRef]
- Vargas-Sánchez, M.; Alcocer, J.; Oseguera, L.A. Seston and eutrophication on a tropical karst lake district: Lagunas de Montebello, Chiapas, Mexico. Limnetica 2022, 41, 1. [Google Scholar] [CrossRef]
- Comisión Nacional de Áreas Naturales Protegidas (CONANP). Programa de conservación y manejo Parque Nacional Lagunas de Montebello; Secretaría de Medio Ambiente y Recursos Naturales: Mexico City, Mexico, 2007; pp. 9–10. ISBN 978-968-817-848-5. [Google Scholar]
- Comisión Nacional de Áreas Naturales Protegidas (CONANP). Programa de Monitoreo de Calidad del Agua, Estudio para monitorear los parámetros de calidad del agua de las lagunas de Montebello. 2011, pp. 17–29. Available online: https://docplayer.es/5046132-Parque-nacional-lagunas-de-montebello-programa-de-monitoreo-de-calidad-del-agua.html (accessed on 17 July 2022).
- Comisión Nacional del Agua (CONAGUA). Plan de gestión de la cuenca del Río Grande-Lagunas de Montebello, Chiapas, México. 2009, pp. 37–45. Available online: https://transparencia.comitan.gob.mx/ART74/I/DESARROLLO_RURAL/plan_de_gestion_cuencas.pdf (accessed on 17 July 2022).
- García, E. Modificaciones al sistema de clasificación climática de Köppen; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2004. [Google Scholar]
- Hansen, H.P.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis; Grasshoff, K., Kremling, K., Ehrhardt, M., Eds.; Wiley-Verlag: Weinheim, Germany, 1999. [Google Scholar]
- Solórzano, L. Determination of ammonia in natural waters by the phenol hypochlorite method. Limnol. Oceanogr. 1969, 14, 799–801. [Google Scholar] [CrossRef]
- Treguer, P.; Le Corre, P. Manuel D’analyse des Sels Nutritifs Dans L’eau de Mer (Utilisation de L’autoanalyzer ii Technicon r), 2nd ed.; UBO: Brest, Franch, 1974. [Google Scholar]
- Weyhenmeyer, G.A.; Håkanson, L.; Meili, M. A validated model for daily variations in the flux, origin, and distribution of settling particles within lakes. Limnol. Oceanogr. 1997, 42, 1517–1529. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Jellison, R.; Melack, J.M. Nitrogen limitation and particulate elemental ratios of seston in hypersaline Mono Lake, California, USA. Hydrobiologia 2001, 466, 1–12. [Google Scholar] [CrossRef]
- Arar, E.J.; Collins, G. In Vitro Determination of Chlorophyll a and Pheophytin a in Marine and Freshwater Algae by Fluorescence (445.0); US Environmental Protection Agency: Washington, DC, USA, 1997. [Google Scholar]
- Vollenweider, R.A.; Talling, J.F.; Westlake, D.F. A Manual on Methods for Measuring Primary Production in Aquatic Environments; Blackwell Scientific: Oxford, UK, 1974. [Google Scholar]
- American Public Health Association-APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005; pp. 2–30–2–34. [Google Scholar]
- Mora, L.; García, L.A.; Ramos, Y.R.; Bonifaz, R.; Escolero, O. Description of Chemical Changes in a Large Karstic System: Montebello, Mexico. Procedia Earth Planet. Sci. 2017, 17, 829–832. [Google Scholar] [CrossRef]
- Borges, A.V.; Abril, G.; Bouillon, S. Carbon dynamics and CO2 and CH4 outgassing in the Mekong delta. Biogeosciences 2018, 15, 1093–1114. [Google Scholar] [CrossRef] [Green Version]
- Goldenfum, J.A. International Hydropower Association, International Hydropower Association, Unesco, & Greenhouse Gas Status of Freshwater Reservoirs Project. In GHG Measurement Guidelines for Freshwater Reservoirs; International Hydropower Association (IHA): London, UK, 2010. [Google Scholar]
- DelSontro, T.; Boutet, L.; St-Pierre, A.; del Giorgio, P.A.; Prairie, Y.T. Methane ebullition and diffusion from northern ponds and lakes regulated by the interaction between temperature and system productivity. Limnol. Oceanogr. 2016, 61, S62–S77. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 17 July 2022).
- Hebbali, A. olsrr: Tools for Building OLS Regression Models. R Package Version 0.5.3. 2020. Available online: https://CRAN.R-project.org/package=olsrr (accessed on 17 July 2022).
- Durbin, J.; Watson, G.S. Testing for Serial Correlation in Least Squares Regression. II. Biometrika 1951, 38, 159. [Google Scholar] [CrossRef]
- Zeileis, A.; Hothorn, T. Diagnostic Checking in Regression Relationships. R News 2002, 2, 7–10. Available online: https://CRAN.R-project.org/doc/Rnews/ (accessed on 17 July 2022).
- Stefano, S.; Quartagno, M.; Tamburini, M.; Robinson, D. _orcutt: Estimate Procedure in Case of First Order Autocorrelation_. 2018. Available online: https://CRAN.R-project.org/package=orcutt (accessed on 17 July 2022).
- Goldman, C.R.; Elser, J.J.; Richards, R.C.; Reuters, J.E.; Priscu, J.C.; Levin, A.L. Thermal stratification, nutrient dynamics, and phytoplankton productivity during the onset of spring phytoplankton growth in Lake Baikal, Russia. Hydrobiologia 1996, 331, 9–24. [Google Scholar] [CrossRef]
- Cole, J.J.; Pace, M.L.; Carpenter, S.R.; Kitchell, J.F. Persistence of net heterotrophy in lakes during nutrient addition and food web manipulations. Limnol. Oceanogr. 2000, 45, 1718–1730. [Google Scholar] [CrossRef] [Green Version]
- Vachon, D.; Solomon, C.T.; del Giorgio, P.A. Reconstructing the seasonal dynamics and relative contribution of the major processes sustaining CO2 emissions in northern lakes. Limnol. Oceanogr. 2016, 62, 706–722. [Google Scholar] [CrossRef]
- Riera, J.L.; Schindler, J.E.; Kratz, T.K. Seasonal dynamics of carbon dioxide and methane in two clear-water lakes and two bog lakes in northern Wisconsin, USA. Can. J. Fish. Aquat. Sci. 1999, 56, 265–274. [Google Scholar] [CrossRef]
- López Bellido, J.; Tulonen, T.; Kankaala, P.; Ojala, A. CO2 and CH4 fluxes during spring and autumn mixing periods in a boreal lake (Pääjärvi, southern Finland). J. Geophys. Res. 2009, 114. [Google Scholar] [CrossRef]
- Bastviken, D. Methane. Encycl. Inland Waters 2022, 136–154. [Google Scholar] [CrossRef]
- Hanson, P.C.; Carpenter, S.R.; Armstrong, D.E.; Stanley, E.H.; Kratz, T.K. Lake dissolved inorganic carbon and dissolved oxygen: Changing drivers from days to decades. Ecol. Monogr. 2006, 76, 343–363. [Google Scholar] [CrossRef]
- Macklin, P.A.; Suryaputra, I.G.N.A.; Maher, D.T.; Santos, I.R. Carbon dioxide dynamics in a lake and a reservoir on a tropical island (Bali, Indonesia). PLoS ONE 2018, 13, e0198678. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.J.; Caraco, N.F.; Kling, G.W.; Kratz, T.K. Carbon Dioxide Supersaturation in the Surface Waters of Lakes. Science 1994, 265, 1568–1570. [Google Scholar] [CrossRef] [Green Version]
- Glissman, K.; Chin, K.J.; Casper, P.; Conrad, R. Methanogenic Pathway and Archaeal Community Structure in the Sediment of Eutrophic Lake Dagow: Effect of Temperature. Microb. Ecol. 2004, 48, 389–399. [Google Scholar] [CrossRef]
- Aguirrezabala-Campano, T.; Gerardo-Nieto, O.; Gonzalez-Valencia, R.; Souza, V.; Thalasso, F. Methane dynamics in the subsaline ponds of the Chihuahuan Desert: A first assessment. Sci. Total Environ. 2019, 666, 1255–1264. [Google Scholar] [CrossRef]
- Xiao, Q.; Xu, X.; Duan, H.; Qi, T.; Qin, B.; Lee, X.; Hu, Z.; Wang, W.; Xiao, W.; Zhang, M. Eutrophic Lake Taihu as a significant CO2 source during 2000–2015. Water Res. 2020, 170, 115331. [Google Scholar] [CrossRef]
- Guzmán-Arias, A.P. Evaluación de un lago tropical como fuente de CO2 hacia la atmósfera (lago Alchichica, Puebla, México). Master Thesis, Universidad Nacional Autónoma de México, México, México, 2015. [Google Scholar]
- Rudberg, D.; Duc, N.T.; Schenk, J.; Sieczko, A.K.; Pajala, G.; Sawakuchi, H.O.; Verheijen, H.A.; Melack, J.M.; MacIntyre, S.; Karlsson, J.; et al. Diel Variability of CO2 Emissions from Northern Lakes. J. Geophys. Res. Biogeosciences 2021, 126, e2021JG006246. [Google Scholar] [CrossRef]
- López, P.; Marcé, R.; Armengol, J. Net heterotrophy and CO2 evasion from a productive calcareous reservoir: Adding complexity to the metabolism-CO2 evasion issue. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef]
- Morales-Williams, A.M.; Wanamaker, A.D.; Williams, C.J.; Downing, J.A. Eutrophication Drives Extreme Seasonal CO2 Flux in Lake Ecosystems. Ecosystems 2020, 24, 434–450. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Lake | Lat | Long | Alt | Volume | Area | ZMAX | ZM |
|---|---|---|---|---|---|---|---|
| (N) | (W) | (m a.s.l.) | (km3) | (ha) | (m) | (m) | |
| TZ | 16.075 | 91.665 | 1490 | 0.09 | 306.6 | 86 | 28.9 |
| SJ | 16.106 | 91.738 | 1454 | 0.006 | 60.6 | 30 | 10.3 |
| SL | 16.126 | 91.753 | 1455 | 0.02 | 181.3 | 67 | 11.2 |
| Variable | n | Parameter | Lake | KWL | |||||
|---|---|---|---|---|---|---|---|---|---|
| TZ (Oligotrophic) | SJ (Mesotrophic) | SL (Eutrophic) | |||||||
| R/W | D/C | R/W | D/C | R/W | D/C | ||||
| T | 253 | X ± SD | 21.8 ± 0.2 | 19.2 ± 0.4 | 20.9 ± 0.3 | 21.4 ± 0.3 | 21.2 ± 0.1 | 20.8 ± 0.7 | X2 = 9.6, p < 0.01 |
| (°C) | KWS | X2 = 126.5, p < 0.01 | X2 = 4.8, p < 0.05 | X2 = 2.5, p = 0.11 | |||||
| DO | 253 | X ± SD | 7.2 ± 0.73 | 5.4 ± 0.4 | 7.5 ± 0.3 | 6.0 ± 0.1 | 4.7 ± 1.1 | 6.1 ± 3.4 | X2 = 7.4, p < 0.05 |
| (mg L−1) | KWS | X2 = 111.36, p < 0.01 | X2 = 34.71, p < 0.01 | X2 = 4.95, p < 0.01 | |||||
| pH | 253 | X ± SD | 8.5 ± 0.1 | 8.5 ± 0.1 | 9.1 ± 0.04 | 7.6 ± 0.04 | 7.7 ± 0.07 | 9.1 ± 0.3 | X2 = 4.5, p = 0.10 |
| KWS | – | – | – | ||||||
| K25 | 253 | X ± SD | 235.4 ± 2. | 241.9 ± 1.6 | 338.4 ± 1.7 | 333.2 ± 0.6 | 518 ± 0.1 | 609.7 ± 7.1 | X2 = 172, p < 0.01 |
| (µS cm−1) | KWS | X2 = 112.7, p < 0.01 | X2 = 36.5, p < 0.01 | X2 = 22.9, p < 0.01 | |||||
| ZMIX (m) | 6 | X | 34 | 86 * | 10 | 30 * | 5 | 8 | |
| ZSD (m) | 6 | X | 7.2 | 7 | 3.5 | 2.3 | 0.6 | 0.4 | X2 = 4.8, p = 0.1 |
| KWS | – | – | – | ||||||
| Chl-a | 84 | X ± SD | 0.54 ± 0.01 | 0.45 ± 0.01 | 1.0 ± 0.2 | 0.8 ± 0.1 | 46.3 ± 10.1 | 50.8 ± 8.5 | X2 = 72.5, p < 0.01 |
| (µg L−1) | KWS | X2 = 7.5, p < 0.01 | X2 = 12.3, p < 0.01 | X2 = 1.6, p = 0.2 | |||||
| TSS | 57 | X ± SD | 1.2 ± 0.3 | 1.6 ± 1.0 | 4.4 ± 3.0 | 2.9 ± 1.3 | 9.4 ± 2.4 | 7.2 ± 1.3 | X2 = 9.8, p < 0.01 |
| (mg L−1) | KWS | X2 = 0.6, p = 0.44 | X2 = 0, p = 1 | X2 = 2.4, p = 0.12 | |||||
| P-SRP | 24 | X ± SD | 0.1 ± 0.01 | 0.08 ± 0.01 | 0.3 ± 0.1 | 0.2 ± 0.03 | 1.0 ± 0.6 | 43.5 ± 15.6 | X2 = 61.9, p < 0.01 |
| (µmol L−1) | KWS | X2 = 1.9, p = 0.2 | X2 = 13.7, p < 0.001 | X2 = 1.8, p = 0.1 | |||||
| N-NO3 | 24 | X ± SD | 0.8 ± 0.13 | 3.4 ± 0.1 | 3.3 ± 1.7 | 5.3 ± 0.9 | 2.6 ± 1.5 | 2.0 ± 1.1 | X2 = 24.3, p < 0.01 |
| (µmol L−1) | KWS | X2 = 1.7, p = 0.1 | X2 = 13.7, p < 0.001 | X2 = 0.9, p = 0.4 | |||||
| N-NO2 | 24 | X ± SD | 0.1 ± 0.01 | 0.2 ± 0.03 | 0.4 ± 0.1 | 0.4 ± 0.3 | 0.4 ± 0.3 | 0.6 ± 0.4 | X2 = 28.8, p < 0.01 |
| (µmol L−1) | KWS | X2 = 4.3, p < 0.05 | X2 = 0.21, p = 0.6 | X2 = 5.4, p < 0.05 | |||||
| N-NH4 | 24 | X ± SD | 0.6 ± 0.4 | 1.9 ± 0.8 | 12.4 ± 12.2 | 3.7 ± 1.7 | 0.5 ± 0.1 | 0.7 ± 0.1 | X2 = 34.5, p < 0.01 |
| (µmol L−1) | KWS | X2 = 1.3, p = 0.2 | X2 = 3.4, p = 0.06 | X2 = 1.9, p = 0.2 | |||||
| Lake | Season | n | Z | DIC (mmol L−1) | CCO2 (µmol L−1) | ∇CCO2 | %Sat CO2 | ||
|---|---|---|---|---|---|---|---|---|---|
| (m) | X | SD | X | SD | µmol L−1 m−1 | ||||
| TZ | R/W | 12 | 0 | 0.84 | 0.05 | 13.9 | 0.1 | 0.5 | 127.4 |
| 34 | 0.91 | 0.04 | 14.8 | 2.2 | 135.7 | ||||
| 40 | 0.99 | 0.02 | 42.4 | 6.2 | 388.6 | ||||
| D/C | 12 | 0 | 0.77 | 0.02 | 17.4 | 1.6 | −0.1 | 159.3 | |
| 34 | 0.77 | 0.04 | 16.4 | 0.6 | 150.7 | ||||
| 40 | 0.75 | 0.04 | 14.4 | 0.9 | 132.4 | ||||
| SJ | R/W | 12 | 0 | 1.26 | 0.08 | 35.2 | 0.8 | −0.2 | 323.1 |
| 5 | 1.29 | 0 | 39.9 | 6.6 | 366.5 | ||||
| 10 | 1.27 | 0.01 | 33.1 | 1.4 | 303.9 | ||||
| D/C | 12 | 0 | 1.07 | 0.11 | 18.8 | 0.7 | 0.1 | 172.2 | |
| 5 | 1.07 | 0.06 | 22.2 | 2.8 | 204 | ||||
| 10 | 1.1 | 0 | 19.8 | 1.9 | 181.4 | ||||
| SL | R/W | 12 | 0 | 1.11 | 0.04 | 29.2 | 4.3 | 6.4 | 267.6 |
| 5 | 1.06 | 0.08 | 46.3 | 12.9 | 424.8 | ||||
| 15 | 1.37 | 0.33 | 122.4 | 65.3 | 1122.8 | ||||
| D/C | 12 | 0 | 0.95 | 0.08 | 18.5 | 7.8 | 0.4 | 169.9 | |
| 5 | 1.01 | 0.05 | 33.6 | 6.1 | 308.3 | ||||
| 15 | 1 | 0.03 | 27.3 | 13.8 | 250.7 | ||||
| TZ (Oligotrophic) | |||||||
|---|---|---|---|---|---|---|---|
| Regression Summary for the Dependent Variable: CCO2 Adjusted R²= 0.9613, F(5,18) = 89.4, p < 0.0001 Std. Error of Estimate: 2.37 | |||||||
| β | Std. Err. of β | p-Level | Multiple R2 | R2 Change | F-to Enter | p-Level | |
| Intercept | 224.12 | 104.69 | 0.046 | ||||
| pH | −16.43 | 5.25 | 0.006 | 0.8697 | 0.8697 | 145.56 | <0.001 |
| N-NO3 | 3.61 | 0.88 | 0.001 | 0.9194 | 0.0497 | 109.39 | <0.001 |
| DIC | 32.88 | 8.61 | 0.001 | 0.9375 | 0.0181 | 99.918 | <0.001 |
| TSS | −2.05 | 0.7 | 0.009 | 0.9578 | 0.0203 | 107.93 | <0.001 |
| K25 | −0.42 | 0.34 | 0.22 | 0.9613 | 0.0035 | 89.399 | <0.001 |
| SJ (Mesotrophic) | |||||||
| Regression Summary for the Dependent Variable: CCO2 Adjusted R² = 0.7628 F(4,19) = 15.27, p < 0.001 Std. Error of Estimate: 5.21 | |||||||
| β | Std. Err. of β | p-Level | Multiple R2 | R2 Change | F-to Enter | p-Level | |
| Intercept | 209.06 | 50.85 | 0.001 | ||||
| pH | −4.34 | 3.05 | 0.17 | 0.6481 | 0.6481 | 40.51 | <0.001 |
| TSS | 1.65 | 0.62 | 0.015 | 0.6879 | 0.0398 | 23.14 | <0.001 |
| K25 | −0.42 | 0.186 | 0.035 | 0.7315 | 0.0436 | 18.16 | <0.001 |
| N-NO3 | −1.82 | 1.146 | 0.13 | 0.7628 | 0.0313 | 15.27 | <0.001 |
| SL (Eutrophic) | |||||||
| Regression Summary for the Dependent Variable: CCO2 Adjusted R² = 0.9471 F(5,18) = 59.26, p < 0.001 Std. Error of Estimate: 11.16 | |||||||
| β | Std. Err. of β | p-Level | Multiple R2 | R2 Change | F-to Enter | p-Level | |
| Intercept | −18.89 | 59.73 | 0.75 | ||||
| Chl-a | −0.61 | 0.68 | 0.39 | 0.6760 | 0.676 | 45.91 | <0.001 |
| TSS | 3.68 | 0.737 | 0.001 | 0.8116 | 0.1356 | 45.23 | <0.001 |
| DO | −3.4 | 1.019 | 0.004 | 0.8776 | 0.066 | 47.79 | <0.001 |
| N-NO3 | 8.67 | 2.275 | 0.001 | 0.9377 | 0.0601 | 71.509 | <0.001 |
| DIC | 37.98 | 21.82 | 0.1 | 0.9427 | 0.005 | 59.26 | <0.001 |
| N-NH4 | 11.9 | 9.98 | 0.25 | 0.9471 | 0.0044 | 50.77 | <0.001 |
| Lake | Season | n | FCO2 (g CO2 m−2 d−1) | Area | FCO2 (t CO2 d−1) | ||
|---|---|---|---|---|---|---|---|
| X | SD | (m2) | X | SD | |||
| TZ | R/W | 9 | 0.88 | 0.46 | 3.06 × 10−6 | 2.68 | 0.001 |
| D/C | 16 | 0.59 | 0.02 | 3.06 × 10−6 | 1.81 | 4.49 × 10−5 | |
| SJ | R/W | 12 | 4.63 | 0.88 | 6.06 × 10−5 | 2.8 | 0.002 |
| D/C | 16 | 1.42 | 0.18 | 6.06 × 10−5 | 0.86 | 0.0001 | |
| SL | R/W | 10 | 9.13 | 1.24 | 1.81 × 10−6 | 16.54 | 0.02 |
| D/C | 11 | 0.29 | 0.02 | 1.81 × 10−6 | 0.53 | 9.37 × 10−6 | |
| Season | Lake | FCO2 | Trophic | Reference | |
|---|---|---|---|---|---|
| X | SD | Status | |||
| R/W | TZ | 0.9 | 0.5 | O | This study |
| SJ | 4.6 | 0.9 | M | This study | |
| SL | 9.1 | 1.2 | E | This study | |
| Warm | Poza Churince | 1.2 | 1 | O | [70] |
| Los Hundidos | 5.6 | 2.7 | O | ||
| Poza Manantial | 15.3 | 10.2 | O | ||
| Poza Azul | 6.5 | 2.7 | O | ||
| Poza Becerra | 22.9 | 5.8 | O | ||
| Taihu | 3.3 | 1.2 | E | [71] | |
| Stratification | Alchichica | 0.1 | 0.4 | O | [72] |
| Venasjön | 0.2 | 0.8 | E | [73] | |
| D/C | TZ | 0.6 | 0 | O | This study |
| SJ | 1.4 | 0.2 | M | This study | |
| SL | 0.3 | 0 | E | This study | |
| Cold | Los Hundidos | 3.2 | 1.6 | O | [70] |
| Poza Azul | 7.7 | 3.7 | O | ||
| Poza Becerra | 25.6 | 7.2 | O | ||
| Taihu | 0.2 | 0.8 | E | [71] | |
| Mixing | Alchichica | 0.1 | 0.4 | O | [72] |
| Sau | 0.3 | E | [74] | ||
| Venasjön | 3.3 | 1.2 | E | [73] | |
| NM | Ljusvanttentjärn | 0.4 | 0.1 | O | |
| Parsen | 1.1 | 0.5 | M | ||
| Badger | 2.2 | E | [75] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Sánchez, M.; Alcocer, J.; Sánchez-Carrillo, S.; Oseguera, L.A.; Rivera-Herrera, E.M.; Soria-Reinoso, I.; Guzmán-Arias, A.; García-Oliva, F.; Merino-Ibarra, M. Carbon Dioxide Concentration and Emissions along a Trophic Gradient in Tropical Karst Lakes. Water 2023, 15, 13. https://doi.org/10.3390/w15010013
Vargas-Sánchez M, Alcocer J, Sánchez-Carrillo S, Oseguera LA, Rivera-Herrera EM, Soria-Reinoso I, Guzmán-Arias A, García-Oliva F, Merino-Ibarra M. Carbon Dioxide Concentration and Emissions along a Trophic Gradient in Tropical Karst Lakes. Water. 2023; 15(1):13. https://doi.org/10.3390/w15010013
Chicago/Turabian StyleVargas-Sánchez, Mariana, Javier Alcocer, Salvador Sánchez-Carrillo, Luis A. Oseguera, Erika M. Rivera-Herrera, Ismael Soria-Reinoso, Andrea Guzmán-Arias, Felipe García-Oliva, and Martín Merino-Ibarra. 2023. "Carbon Dioxide Concentration and Emissions along a Trophic Gradient in Tropical Karst Lakes" Water 15, no. 1: 13. https://doi.org/10.3390/w15010013
APA StyleVargas-Sánchez, M., Alcocer, J., Sánchez-Carrillo, S., Oseguera, L. A., Rivera-Herrera, E. M., Soria-Reinoso, I., Guzmán-Arias, A., García-Oliva, F., & Merino-Ibarra, M. (2023). Carbon Dioxide Concentration and Emissions along a Trophic Gradient in Tropical Karst Lakes. Water, 15(1), 13. https://doi.org/10.3390/w15010013