1. Introduction
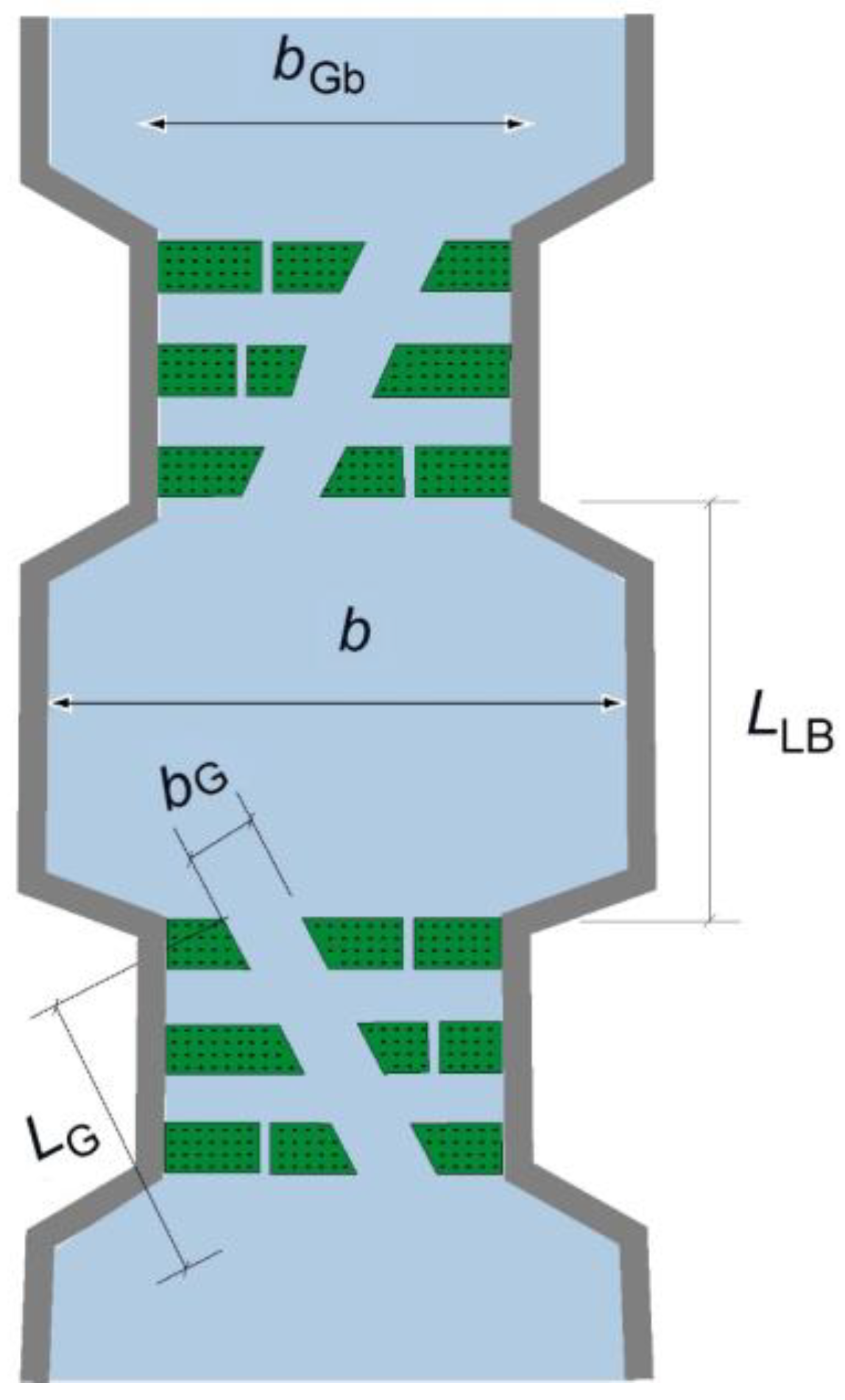
Brush-type fish passes consist of a sloping channel with fixed brush blocks (bristle packs); thus, they provide reduced velocities of less than 1 m/s to ease the upstream movement of fish. This type of upstream fish pass is known to have a variety of structures inside the water body, which provide low velocities and ample resting room for small or weak fish [
1]. Brush blocks are attached on the base of a rectangular channel as roughness elements (
Figure 1). The bristle packs consist of several bristle bundles, each containing five to eight individual elastic bristles. Until 2010, oval plastic bristles with a thickness of 4–6 mm (on average 5 mm) were used, but since 2011, round plastic bristles with a diameter of 6 mm have been in use. The bristle bundles are produced by inserting the bristle bundles into a perforated plastic plate. The carrier plate and, thus, the bristle packs are prepared from a standard base plate where a wide variety of shapes can be cut or welded together. The bristle packs are screwed directly onto the channel bottom or raised individual foundations (e.g., concrete bodies). The outflow in the channel occurs via passages between the bristle packs and through the bristles themselves [
2,
3,
4]. A channel with brush blocks is principally considered a rough channel with individual distributed macro-roughness elements. Brush blocks affect the flow by forming a flow-through obstacle, deflecting flow, and enhancing form drag and skin friction. The flow in the channel, in which hydraulic elements have permeability, is a subcritical and quasi-uniform flow. In a brush fish pass, the energy dissipation is somewhat effective because many flow-induced vibrating bristles initiate strong energy dissipation [
5,
6,
7]. Fish-passage monitoring studies in Germany [
1,
5] revealed that the brush fish pass works well, as designed, and has superior performance compared to other fish-pass types.
The literature provides several studies on the flow and turbulence characteristics of conventional fishways such as vertical-slot passes [
8,
9], pool-weir passes [
10,
11], and natural-like passes [
12,
13]. The common feature of those passes is that the hydraulic energy is mainly dissipated in an energy cascade process. In vertical slots, the turbulent jets plunge into the surrounding pools, and in the natural type, the wake behind the macro-roughness elements dissipates the excess energy. Most fish-pass designs neglect the swimming capacity of smaller and weaker fish. For instance, Chanson and Leng [
14] suggested that a low-velocity zone, which should not exhibit strong recirculation or negative velocity, should be provided for the upstream passage of small-bodied fish. Several test results have shown that fish seek low-velocity zones to pass through the hydraulic structure [
14]. The brush fish pass differs from conventional types of fish passages by providing a nearly optimal migration corridor under uniform and subcritical flow regimes for smaller and weaker fish [
15].
The present study investigates the flow patterns, turbulence, and energy dissipation characteristics of a brush-type fish pass in a pilot hydropower project. Additionally, the effect of the inlet boundary conditions on these parameters, which was ignored in previous studies, is also investigated. Finally, the hydraulic performance of the fish pass is linked to the biological data obtained from the same fishway.
2. Materials and Methods
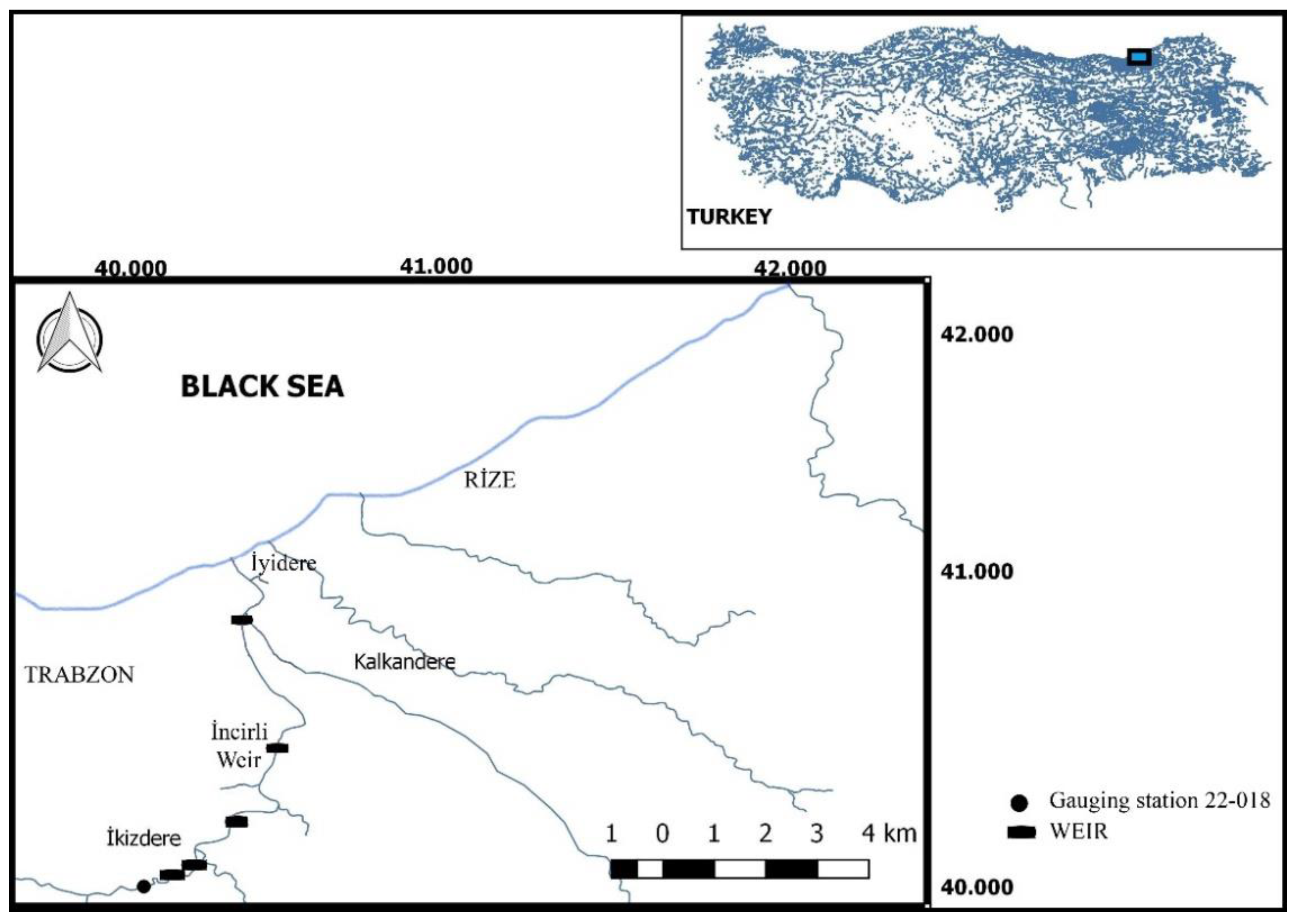
Hydrodynamic characteristics of a 46.2 m long, 1.1 m wide, and 1 m deep brush-type fish pass with a total head of 5 m were monitored at the case study hydropower plant (HPP) Incirli, which is located on the Iyidere River on the East Black Sea Region of Turkey. Iyidere River Basin is rich in biodiversity, with 13 fish species. Its catchment area is 1053 km
2, and the annual average river discharge is 27.8 m
3/s. In the river basin, a cascade-type hydropower system has been developed, and the Incirli HPP is the last HPP downstream of the river in the barbell zone (
Figure 2). Incirli Weir has a catchment area of 895 km
2. Incirli is a diversion-type HPP with a tunnel length of 5.85 km and a gross head of 63 m (net head = 47 m). The HPP is equipped with 3 vertical-axis Francis turbines, with a total output of 25.2 MW at a design discharge of
Qd = 62 m
3/s.
The characteristics of the studied fish species in the studied area are as follows: Alburnoides fasciatus inhabits streams and rivers in foothills with well-oxygenated, fast-flowing water, spawns in small groups, lays eggs deep into gravel with swift currents, and can attain a maximum body length of up to about 13.8 cm. Ponticola rizeensis is found in streams with fresh, running water and with streambeds covered with rounded pebbles, which are presently found only in the Iyidere stream. It is an endangered fish species. Capoeta banarescui inhabits swift-flowing waters with substrates of cobbles and pebbles at the bottom. It is reported to spawn between May and June and can attain a maximum body length of up to about 40 cm. Salmo coruhensis swims in clear and moderately swift-flowing waters with a substrate of sand and pebbles, spawns between October and November, migrates upstream in small groups, can attain a maximum body length of up to about 80 cm, and is a threatened fish species. Barbus tauricus inhabits various habitats, from mountain streams with strong currents to brackish estuaries; spawns on substrates of gravel or rock; can attain a maximum body length of up to about 70 cm; and is a vulnerable fish species.
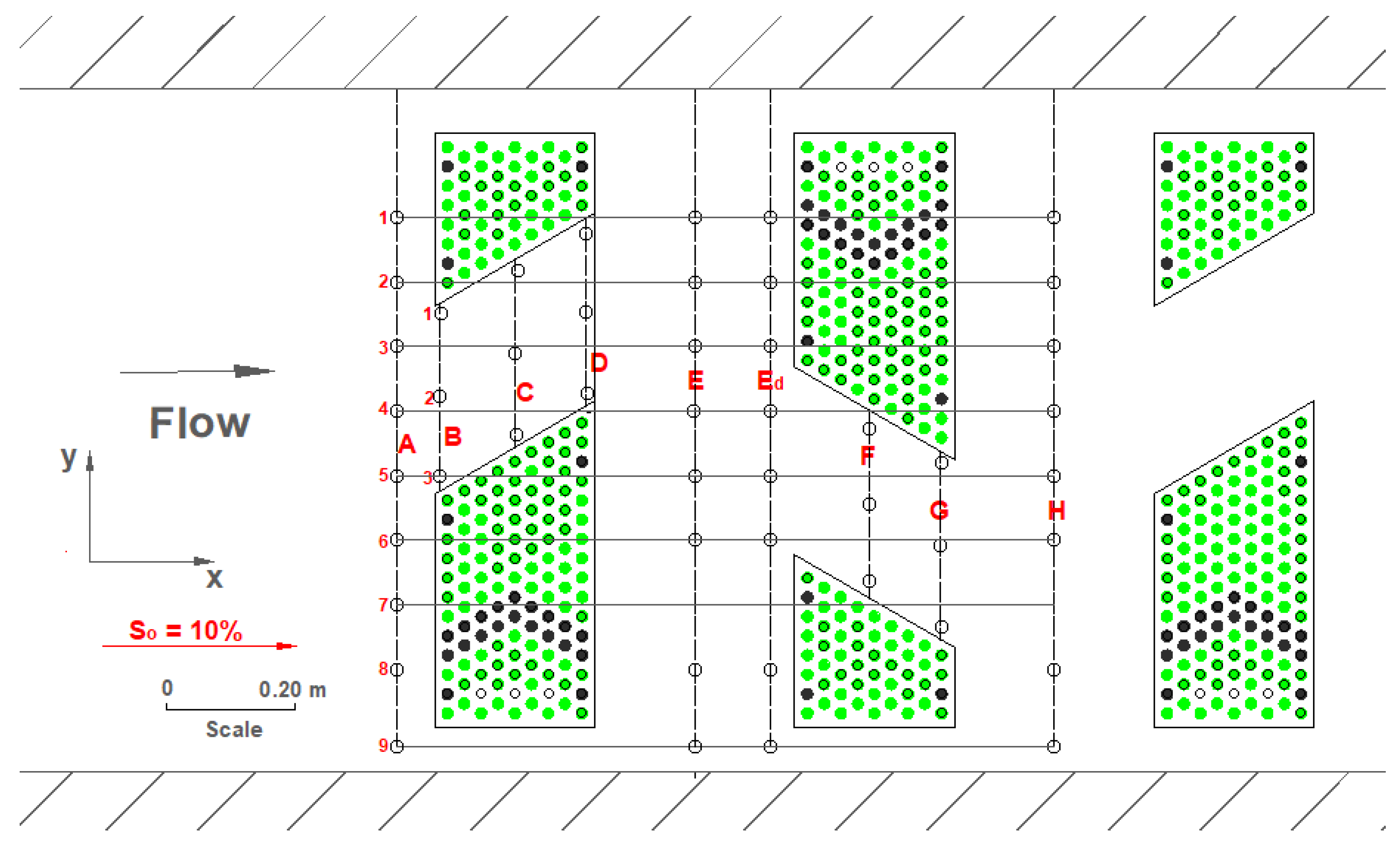
Figure 3 shows the arrangement of the brush blocks in the fish pass constructed in 2017. The grouping of the brush blocks causes pools to form between the groups. Such diagonal arrangement causes flow to cross-exchange constantly and lateral momentum transfer [
16]. Rounded river stones (maximum diameter of 0.16 m) are placed on the channel bed (
Figure 3). This benefits species with low swimming performance. An acoustic Doppler velocimeter (SonTek 16-MHz MicroADV, San Diego, CA, USA) was used to measure three instantaneous velocity components at a densely spaced grid in the fish pass. The employed ADV typically has three receivers and one emitter; its measurement volume is 0.05 m away from the emitter. The sampling frequency and duration were 50 Hz and 30 s, respectively. The employed ADV measures flow velocities from about 1mm/s up to 2.5 m/s with an accuracy of ±1%. Hydraulic conditions tend to have repeating patterns (
Figure 3). Accordingly, a representative basin is selected for the flow and turbulence measurements. The representative basin is located about 6 m downstream from the fish-pass outlet (
Figure 4).
Turbulent flows can be described in terms of their time-averaged value (
) at the point during the sampling period, the fluctuating part of the velocity
, and the instantaneous velocity
u as follows:
The root-mean-square of the instantaneous velocities
in the longitudinal, lateral, and vertical directions, respectively, were calculated at each measurement point, and then the turbulence intensities in three directions were normalized as:
where
U is the cross-sectional average flow velocity. The
x,
y, and
z coordinates represent the streamwise, lateral, and vertical directions, respectively. Then, the Reynolds shear stresses in the vertical direction τ
xz and in the lateral direction τ
xy are computed from
where
ρ is the density of water. The Reynolds shear stresses are induced by velocity fluctuations and affect momentum exchange and mixing processes [
17]. The Reynolds shear stress represents fluid force that involves a velocity gradient and is, therefore, of major importance for fish passage studies. Fish may, therefore, experience shear stress when moving between two layers of different velocities. Reynolds shear stresses have also been shown to strongly affect fish swimming performance and stability [
18,
19]. Silva et al. [
20] identified the Reynolds shear stress in the lateral direction as the key parameter which controls the movement of
Iberian Barbel in the experimental flume. Turbulent kinetic energy per unit mass
k is calculated as follows:
Turbulent kinetic energy, which corresponds to the kinetic energy associated with the fluctuating velocity at a given point, was shown to affect fish swimming performance by increasing energetic costs of swimming [
19]. Gao et al. [
21] identified turbulent kinetic energy as the single most important stimulus for fish trajectories. The average energy dissipation rate per unit mass for uniform flow,
εa (m
2/s
3), is defined as:
Where
So is the channel bed slope,
U is the uniform velocity, and
g is the acceleration due to gravity. Darcy-Weisbach friction factor,
f, is calculated by using Equation (9):
in which
Rh is the hydraulic radius (ratio of the cross-sectional area to the wetted perimeter). Therein, Reynolds Re and Froude number
Fr are defined as:
where
q is the unit discharge,
ν is the kinematic viscosity of water, and
d is the flow depth. Moreover, volumetric energy dissipation ∆
P is computed as follows:
where
Q is the discharge, γ is the specific weight of the water, and
B is the channel width.
Quantifying fish movement is an essential task for evaluating fishway efficiency [
16]. Hence, we installed two underwater GoPro Hero 5 video cameras in the fish pass to investigate fish movements. The cameras’ positions are at the corners of the second brush bar, which is consistent with Schweizer’s [
22] study. Video recordings were carried out in July 2017.
3. Results
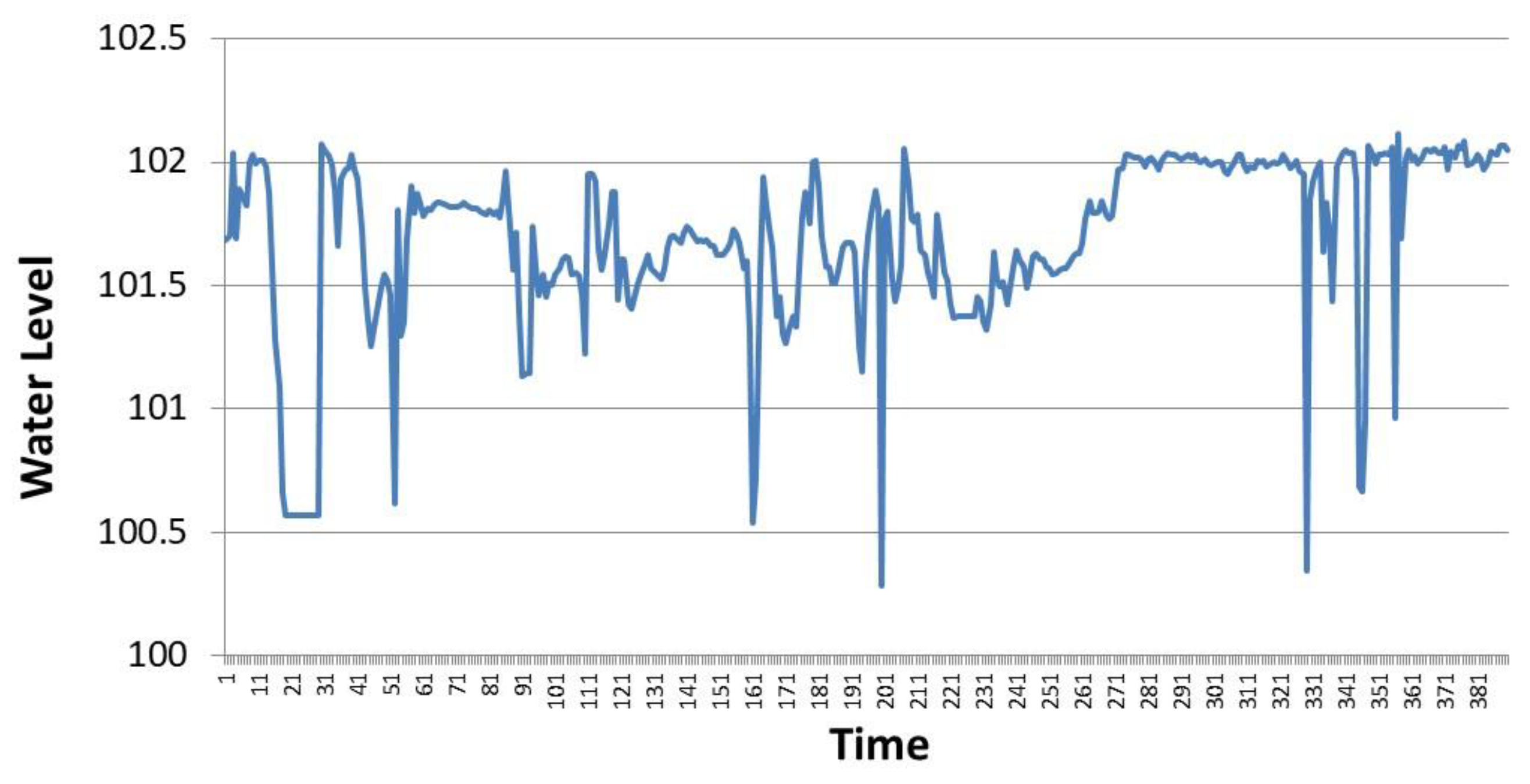
The time series of the water surface elevation of the Incirli HPP reservoir in May 2016 are shown in
Figure 5. The data were taken from the SCADA system of the HPP during operation, with a temporal resolution of 15 min. As seen in
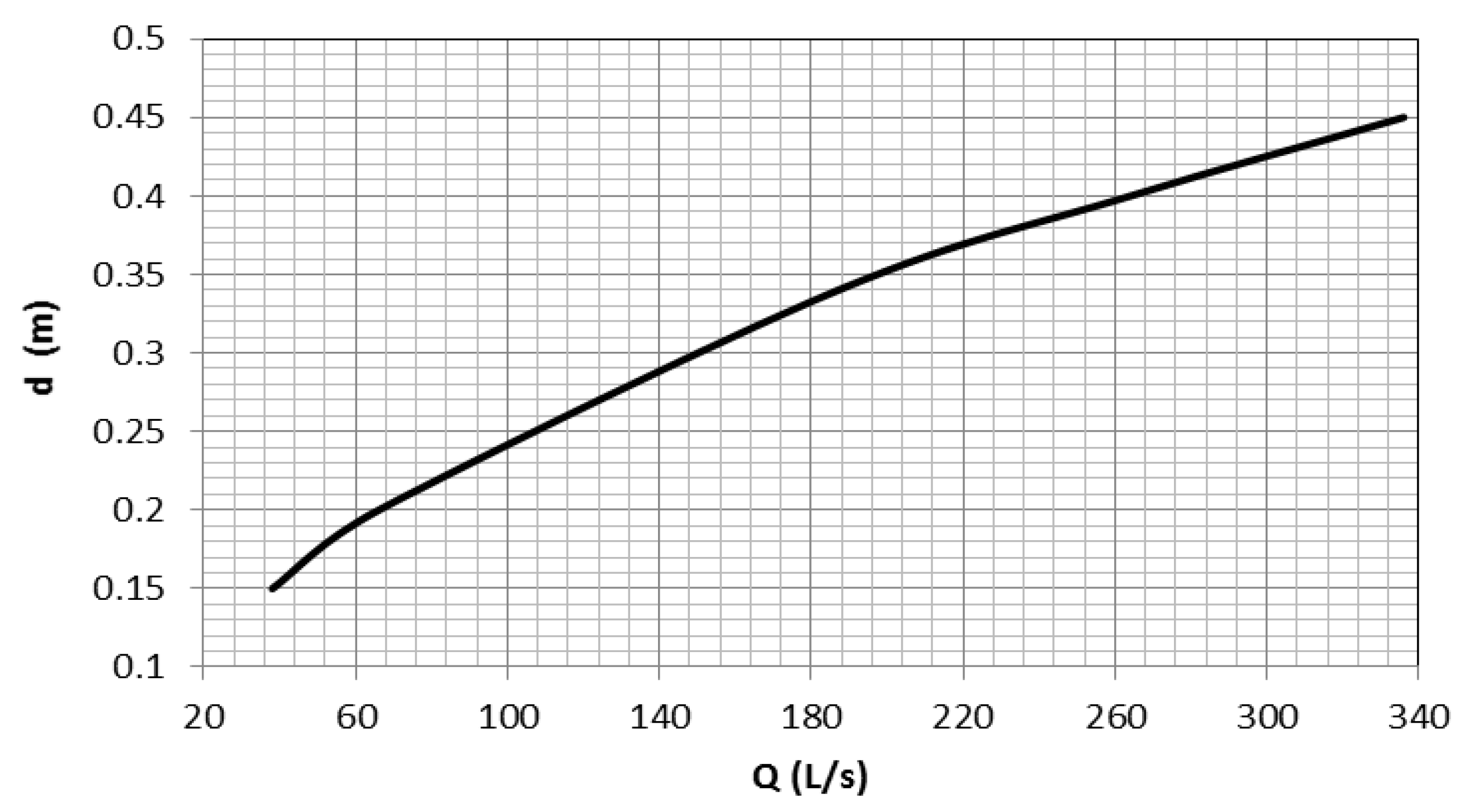
Figure 5, the water surface elevation of the reservoir varies depending on the HPP operating conditions throughout the month. The fish pass receives water when the water surface elevation of the reservoir is higher than 101.20 m. For the fish pass to work effectively, the water surface elevation must be higher than 101.50 m. Overall, the flow rate in the fish pass depends on the reservoir water surface elevation and the geometry of the fish-pass entrance. A flow rating curve shown in
Figure 6 was prepared for the Incirli fish pass based on the measurements made during the field study.
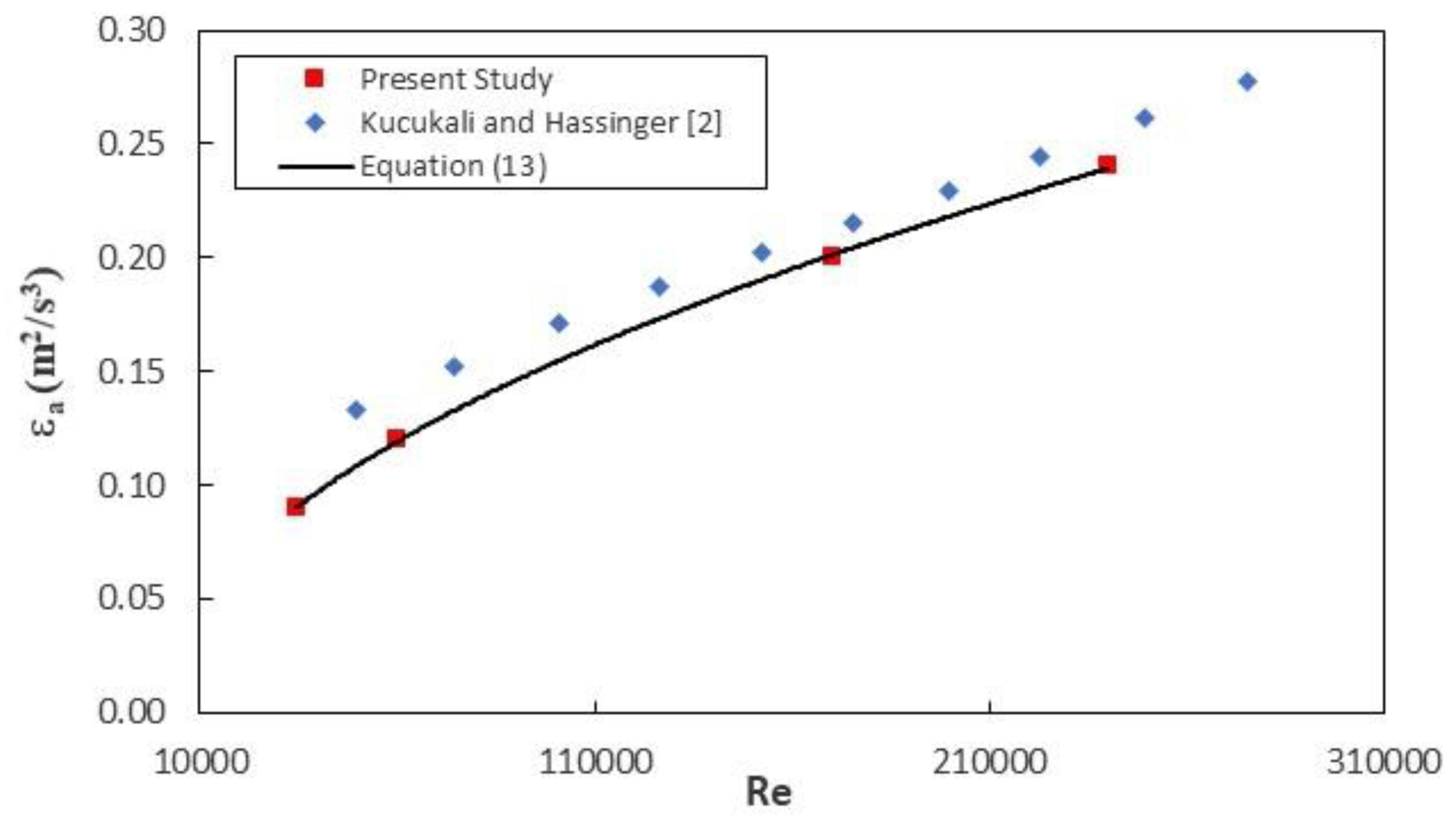
Figure 7 demonstrates the variation of the energy dissipation per unit mass as a function of the Reynolds number, including the data for the baffle-brush composite fish pass (Kucukali and Hassinger [
2]). In both studies, the energy dissipation rises with the rise of the Reynolds number, and the difference between the fishway types is not significant. For Incirli data, the best fit regression equation is found as:
The R
2 value of this equation is 0.99. This result is consistent with the findings of Yagci [
23]’s experimental study of the pool-weir fish-pass type for the range of Re = 1.5 × 10
4–5.4 × 10
4, indicating that despite different fish-pass geometries, the Reynolds number has a significant effect on the amount of energy dissipation per unit mass.
The hydraulic data reveal an inverse relationship between the friction factor and the bristle’s relative submergence,
d/
h (flow depth/bristle height) (
Figure 8). When
d/
h increases from 0.50 to 1.08,
f declines from 6.98 to 2.03. In
Figure 8, the friction factor is plotted as a function of the relative submergence of a bristle with an aerial density of 0.25, and a logarithmic function best fits this trend as:
The R
2 value of this equation is 0.95. Equation (14) demonstrated the dependence of the friction factor on the relative submergence for the proposed fish-pass structure and tested flow conditions (
d/
h = 0.50–1.08). In
Figure 8, the prototype data are compared with the brush-fish-pass physical model data from Kucukali [
24] for three different bed slopes (2–6%).
Figure 8 shows a direct relationship between the flow resistance and bed slope. Furthermore, Kucukali [
24] reported that bed slope is one of the main parameters controlling the magnitudes of energy dissipation and
TKE.
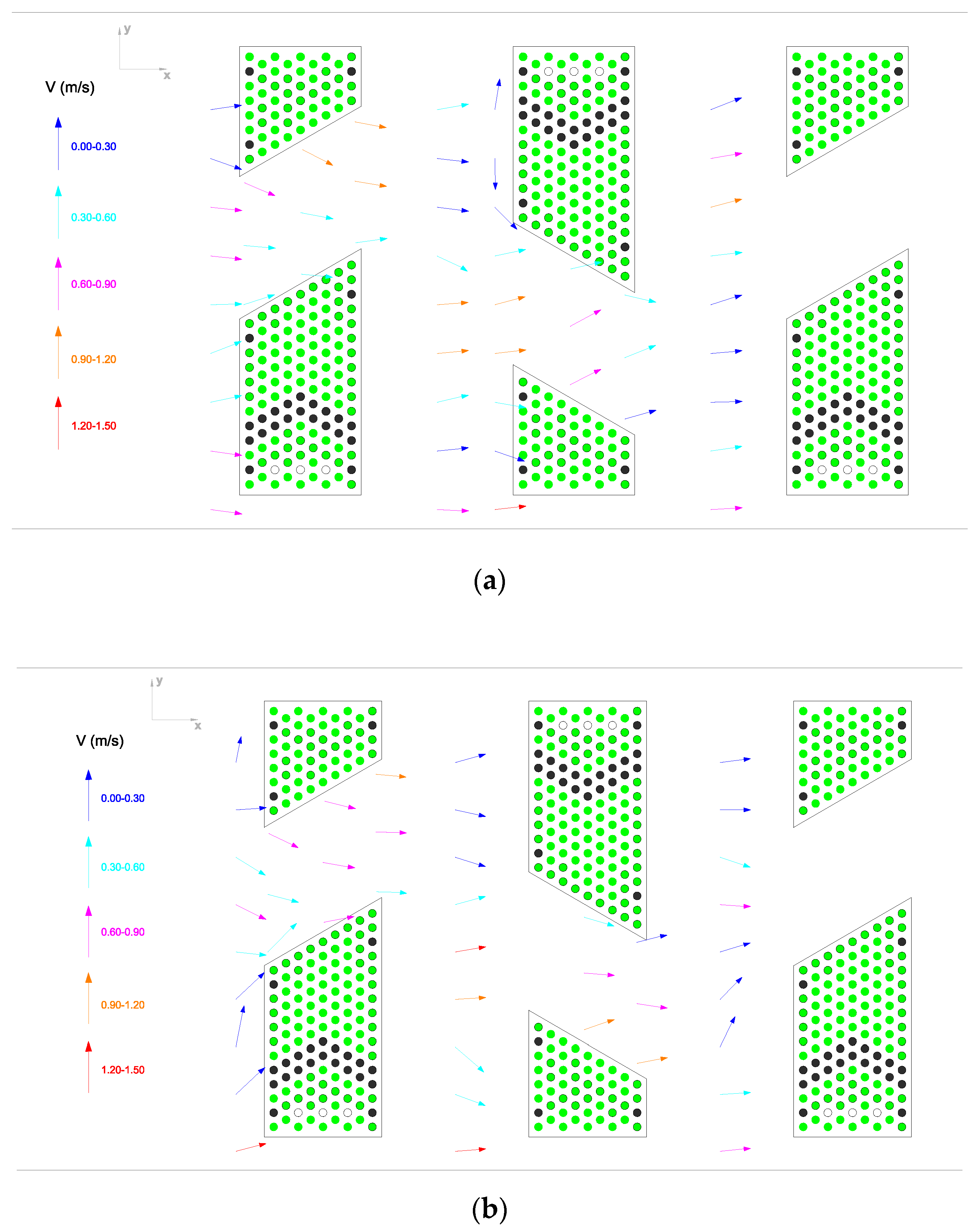
The spatial distribution of the temporally averaged 2D velocity vectors in the brush pass for a discharge of
Q = 0.187 m
3/s is shown in
Figure 9. Flow velocities are high in the S flow (main flow) region. Immediately behind the brush block, the mean flow velocity is reduced by about 70%, but the flow does not separate, and no recirculation zone forms (
Figure 9). However, it can still produce boundary layer separation and form resistance at each bristle scale [
25], which cannot be seen in
Figure 9. The flow resistance within the brush blocks includes the sum of all the bristles’ friction and drag forces.
In all measurements, the highest flow velocities are observed along the axis of number 9 beside the side wall (see points from A to H in
Figure 4), where the free-water jet is formed. The flow velocities observed in the region of the free-water jet are in the order of 1.5 m/s. However, the free jet zone is reserved only for
Salmo coruhensis, which has a high swimming capacity; it is not reserved for all fish species. Additionally, a comparison of the flow field at the bottom and mid-depths (
Figure 9a,b) reveals that there is not a considerable difference between velocity distribution and magnitudes, indicating a two-dimensional flow structure.
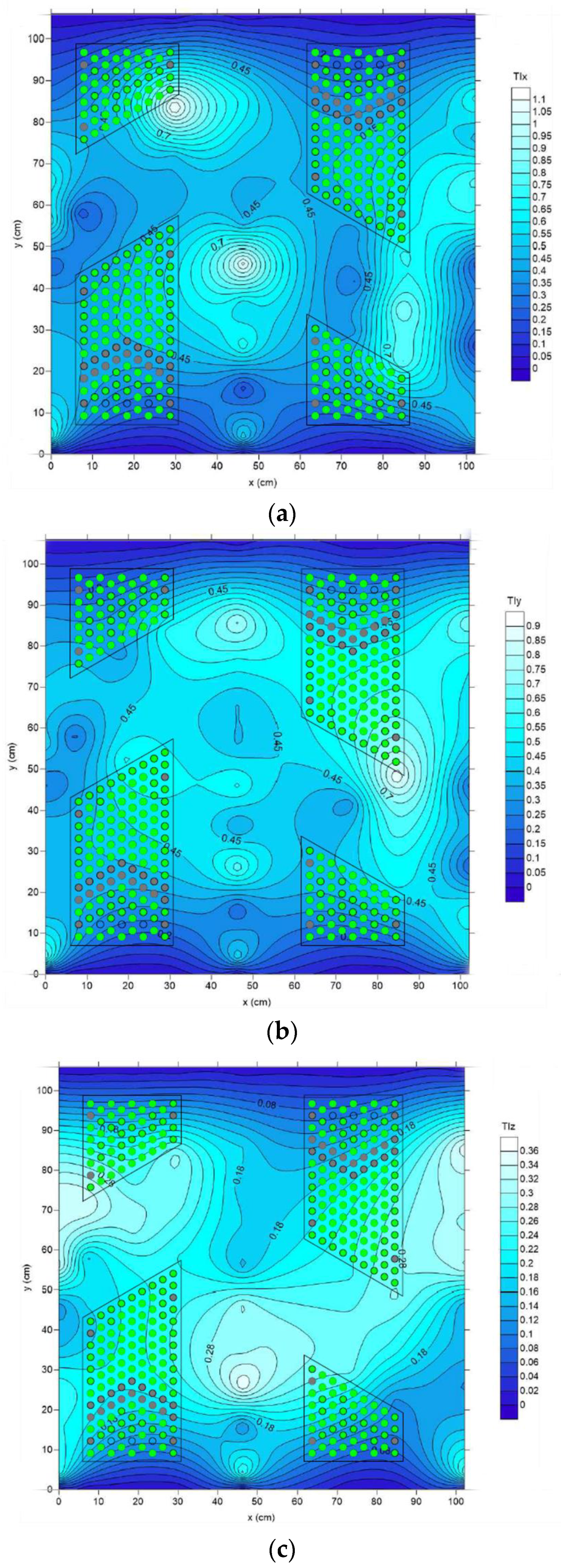
The distribution of streamwise (
TIx), transverse (
TIy), and vertical (
TIz) turbulence intensities in the vicinity of the brush blocks is presented in
Figure 10a, b, and c, respectively.
Figure 10 clearly show that turbulence is not isotropic, and
TIx values are higher than other directions.
TIz distribution is similar to
TIx but with much lower magnitude. Overall,
TIy is responsible for 37% of the turbulent kinetic energy (
TKE). Contributions to
TKE from the streamwise and vertical components are 46% and 17%, respectively. In addition,
Figure 10 highlights that high-velocity zones are associated with high turbulence intensity levels.
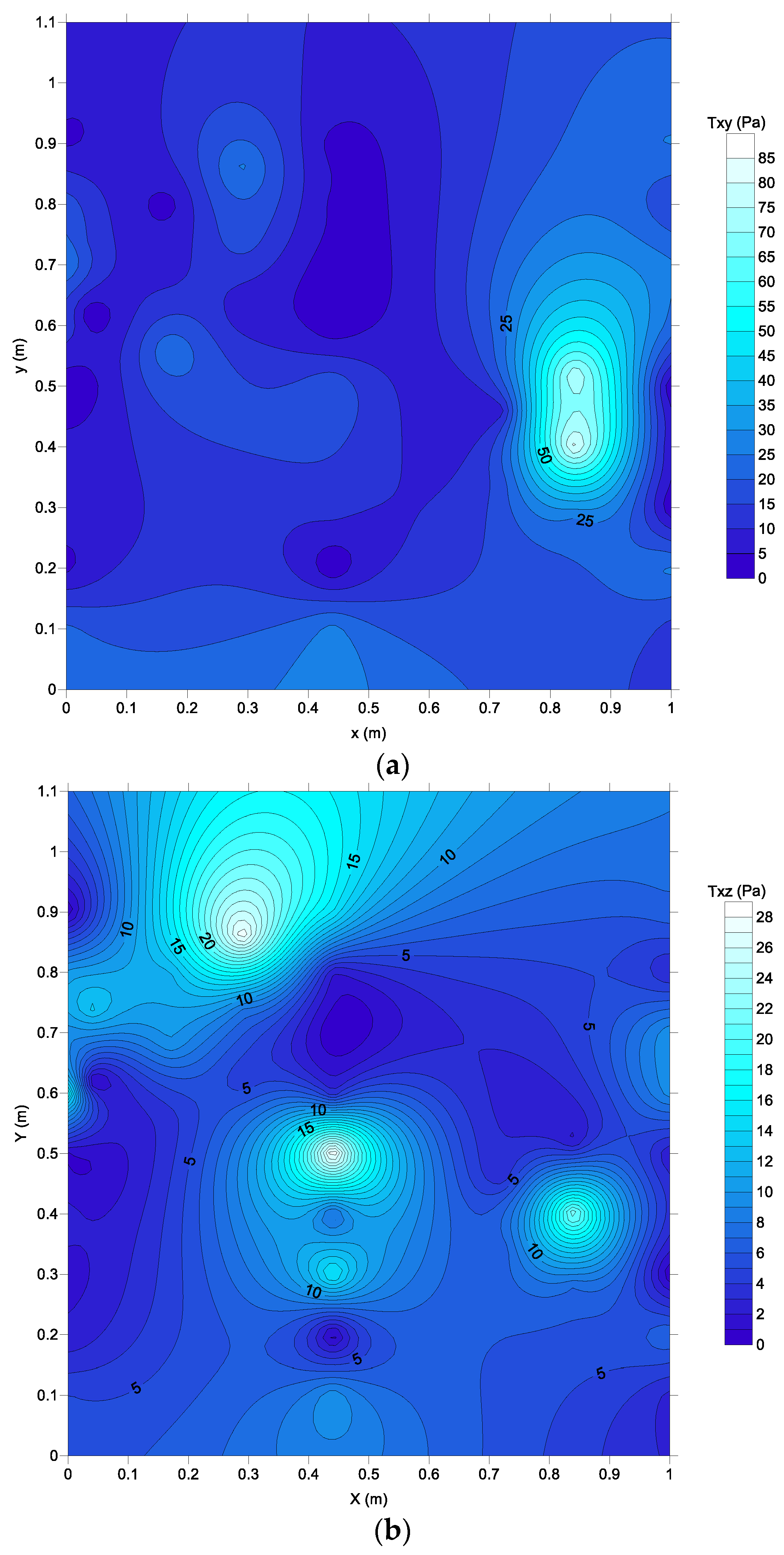
Figure 11 shows the distribution of Reynold shear stresses in vertical and lateral directions. Behind the brush blocks, shear stresses have lower values, while higher stress values are observed in the main flow region. It was found that the spatially averaged lateral component of Reynolds shear stress is higher than the vertical component of Reynolds shear stress by a factor of 2.2. The increase in the lateral component of Reynolds shear stress (
τxy) can be explained due to the horizontal mixing in the vicinity of brush blocks. In brush fish pass, the lateral velocity distribution creates a transverse exchange of momentum. This flow pattern may have important implications for upstream fish migration.
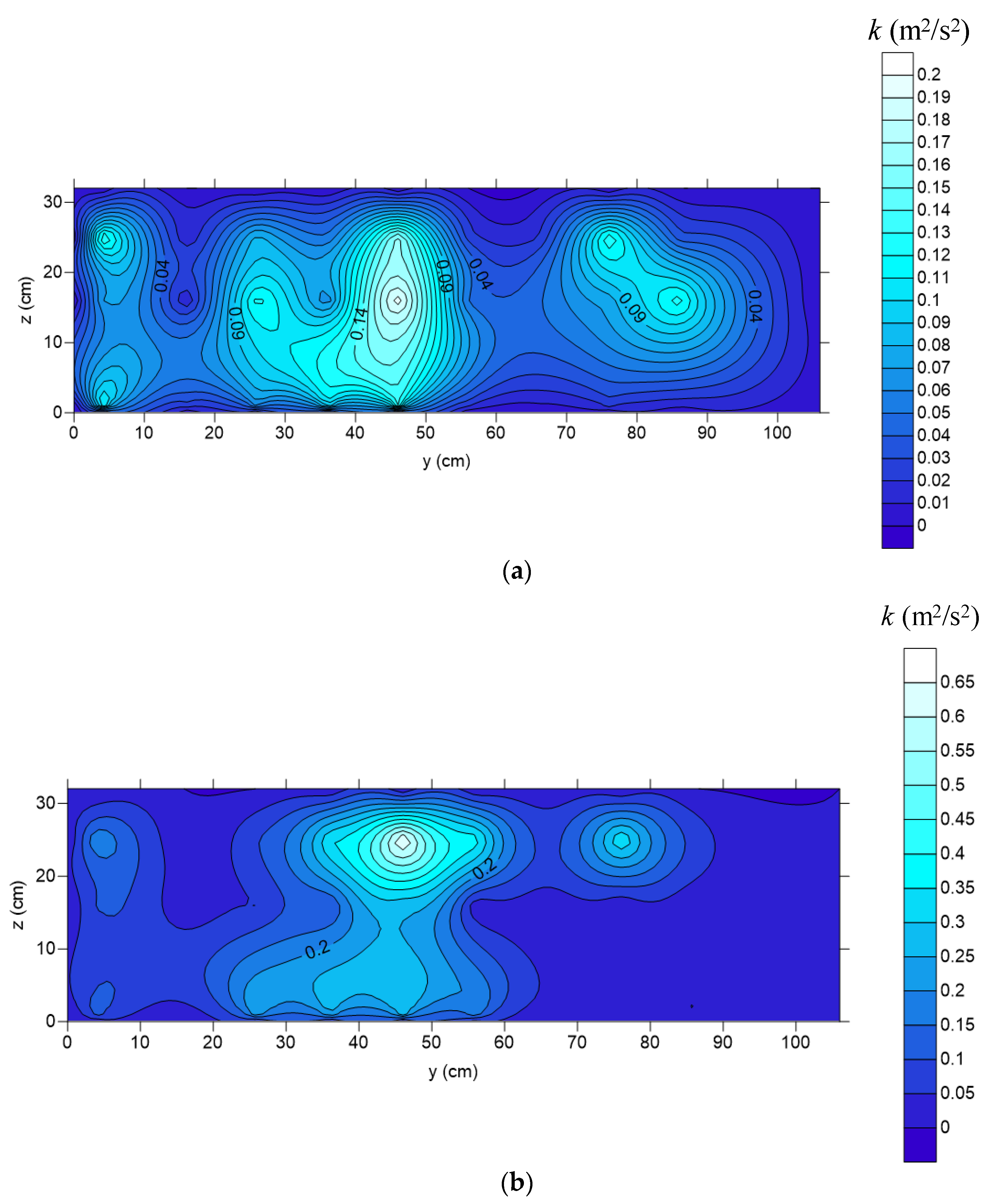
Figure 12 shows the cross-sectional distribution of turbulent kinetic energy at the streamwise location of
E for relative submergences of
d/
h = 0.92 (unsubmerged) for
Q = 0.187 m
3/s and
d/
h = 1.08 (submerged) for
Q = 0.264 m
3/s.
TKE has its maximum value of 0.2 m
2/s
2 in the outer region of the diagonal for
d/
h = 0.92 (
Figure 12a).
TKE values are significantly lower near the wall region than in the S flow region (main flow).
Figure 12 shows that when the bristles are submerged, the maximum
TKE is increased by a factor of three compared to unsubmerged conditions (
Figure 12a).
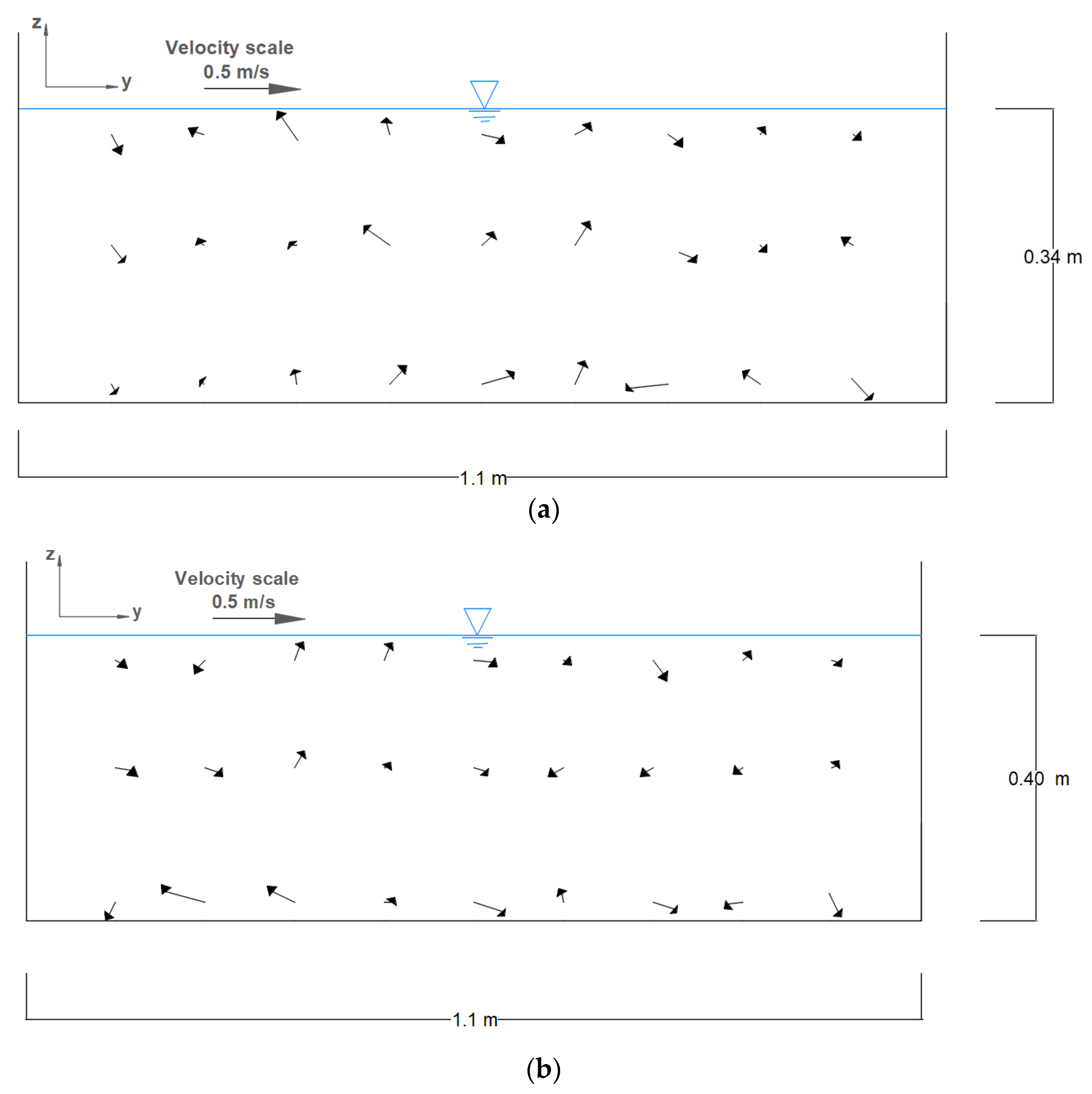
Figure 13 shows the distribution of the temporally averaged velocity vectors (
Vy and
Vz velocities) in the vertical section
E just in front of the second brush bar for two different submergence ratios. Although there is a considerable difference between the order of
TKE magnitudes in submerged and unsubmerged conditions (
Figure 12), velocity magnitudes did not change considerably (
Figure 13).
An analysis of the underwater video recordings revealed that a group of
Alburnoides fasciatus followed the path of the first fish passing the brush block and proceeded with the schooling effect (
Figure 14). The substrates and back of brush blocks create low-velocity zones and shelter for fish. Bottom substrates act as a refugee habitat, especially for fish with a weak swimming capacity (i.e.,
Ponticola rizeensis).
4. Discussion
During the monitoring study, the Reynolds number ranged between 3.45 × 10
4 and 2.4 × 10
5. In the present study, considering the high Reynolds number, the results are not affected by viscous scale effects. Since the formation of vortices and momentum transfer are an important part of the investigations, the Reynolds number is of prime importance. However, this issue, i.e., the effect of Re, was not addressed in some of the previous studies. For instance, Wu et al. [
26] reported that the Reynolds number had no effect on the energy dissipation rate and the dissipated power density was directly proportional to the slope of the vertical-slot fish pass. However, in the present study, the energy dissipation per unit volume tends to increase with the increasing Reynolds number, from 226 to 589 W/m
3. Moreover, the negative correlation of the flow characteristics with the relative submergence agrees with the previous studies [
2,
25,
27].
The monitoring data revealed that a low-velocity zone is ensured for the upstream passage of small-bodied fish [
16]. The maximum measured velocity of 1.5 m/s is well below the recommended value of maximum velocity of 2.0 m/s for fish passes [
6]. Moreover, the present results have been also compared to those from the conventional fish-passage structures. In most of the international standards [
6,
7], it is recommended that Δ
P should not exceed 200 W/m
3. However, the findings reveal that this threshold value is not a relevant criterion for a brush fish pass because a significant amount of energy dissipation takes place in brush blocks with the displacement of bristles.
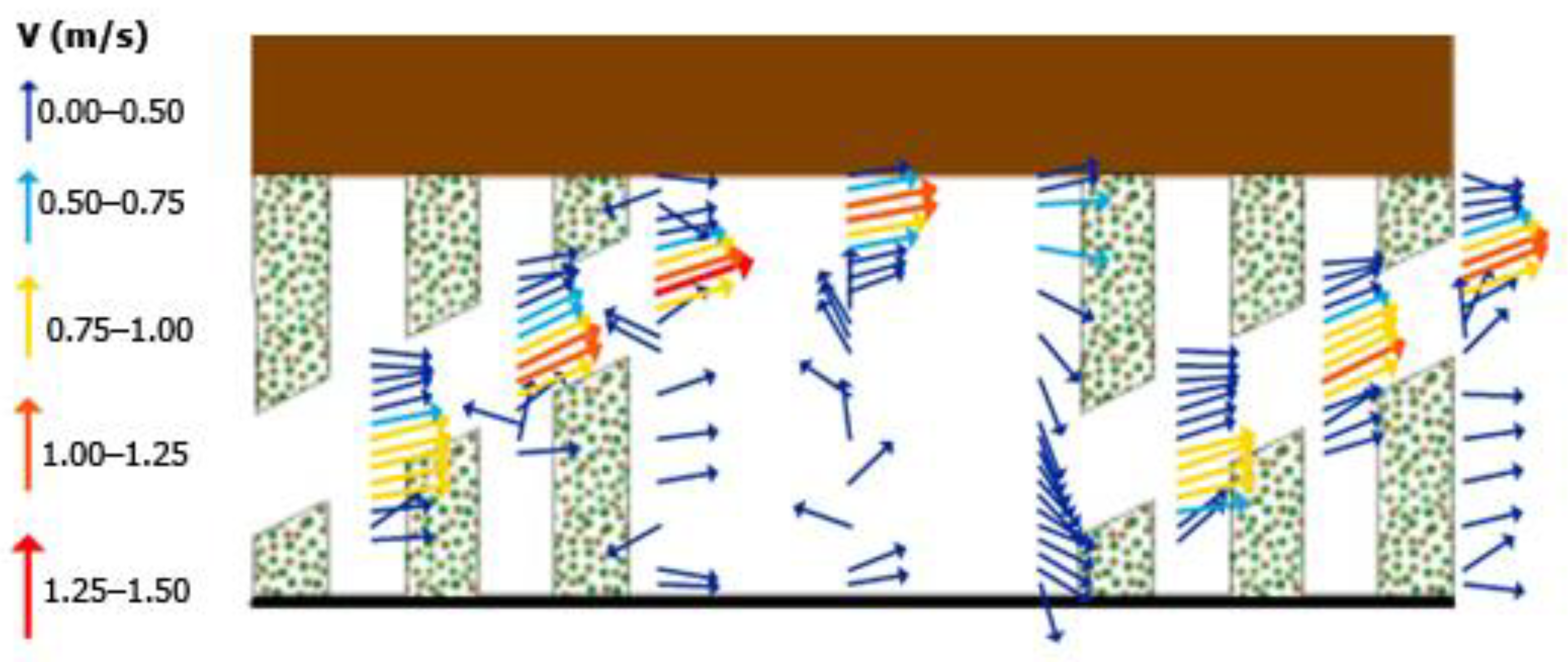
The present maximum measured velocities in the order of 1.5 m/s (
Figure 9) are in good agreement with the findings from the laboratory measurements (
Figure 15) made by Rahn [
3]. Furthermore, the flow pattern with the higher velocities observed in the main flow region (S flow region) in the present study (
Figure 9) is consistent with the physical model results (
Figure 15). Brush blocks have permeability (i.e., it is like a porous media) and, hence, 50% of the flow passes through the brush blocks, which is consistent with the Rahn [
3] laboratory measurements for diagonal brush fish passes. The absence of boundary layer separation behind flexible and permeable hydraulic elements is in line with observations by Nikora et al. [
28] and Mosch [
29]. Mixing and momentum transfer occur due to the velocity gradient in the lateral direction around the diagonal brush blocks. This flow structure prevents both excessive flow acceleration and the presence of high-velocity zones, favoring easy passage for small (i.e.,
Alburnoides fasciatus) and large fish (i.e.,
Salmo coruhensis). Such flow patterns may have important implications for upstream fish migration.
Kucukali et al. [
16] conducted a fish-monitoring study at the fish pass of the present case study HPP and reported that the attraction efficiency of the fish pass was 21.98% for all studied fish species. In total, 55 of the 82 marked fish entering the fish pass ascended -the fish pass and were detected at the upstream antenna. Accordingly, the passage efficiency of the fish pass was calculated as 67.07% for all fish species. On a species basis, the attraction efficiency of the fish pass was the highest for
Alburnoides fasciatus (33.33%) and the lowest for
Ponticola rizeensis (12.31%). Passage efficiency was the highest for
Capoeta banarescui (100%) and the lowest for
Ponticola rizeensis (50.00%). The fact that the passage efficiency of
Capoeta banarescui was the highest could be related to the low number of individuals of that species entering the fish passage. On the other hand, the lowest efficiency of
Ponticola rizeensis, which is endemic to Iyidere, could be due to the weak swimming capacity of the fish. The flow velocities observed in the region of the free-water jet are in the order of 1.5 m/s. However, the free-jet zone is reserved only for
Salmo coruhensis, which has a high swimming capacity, not for all fish species. The reported low attraction efficiency of the brush-type fish pass is more likely due to the habitat destruction of the river downstream and the release of the higher environmental flow below the gates of the weir in the order of 5 m
3/s than at the entrance of the fish pass, indicating the importance of the downstream boundary conditions of the fish passage. The attraction efficiency of the fish pass, therefore, needs to be improved for an efficient upstream fish passage in the study river.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}














