


Twenty-Eight Years of Plant Community Development and Dynamics in the Balize Mississippi River Delta, Louisiana, USA


Abstract
:1. Introduction

2. Materials and Methods
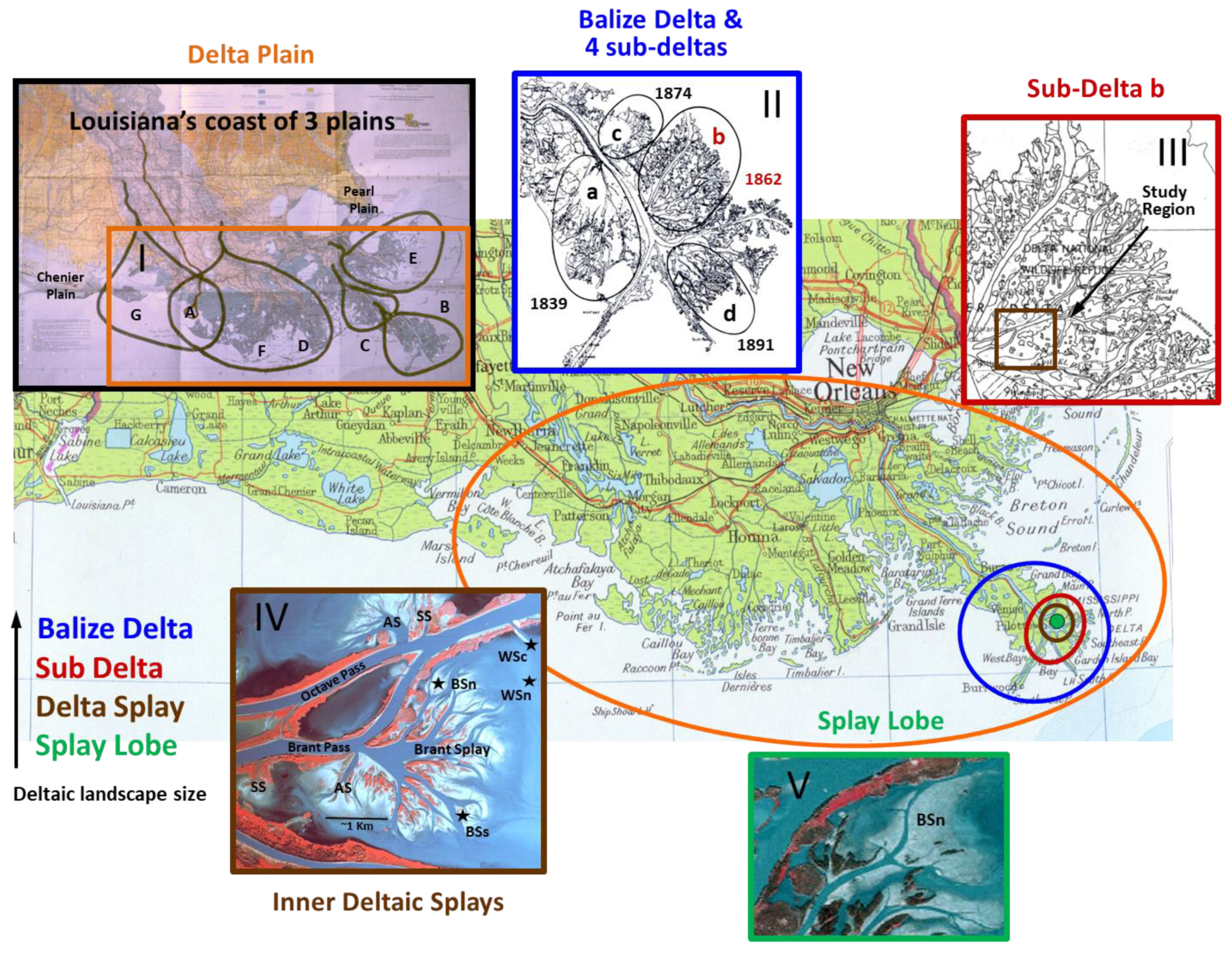
2.1. Study Region and Sites
2.2. Study-Wide Harvested Vegetation
2.3. Analyses of Splay Lobe Phytosociological Dynamics
3. Results
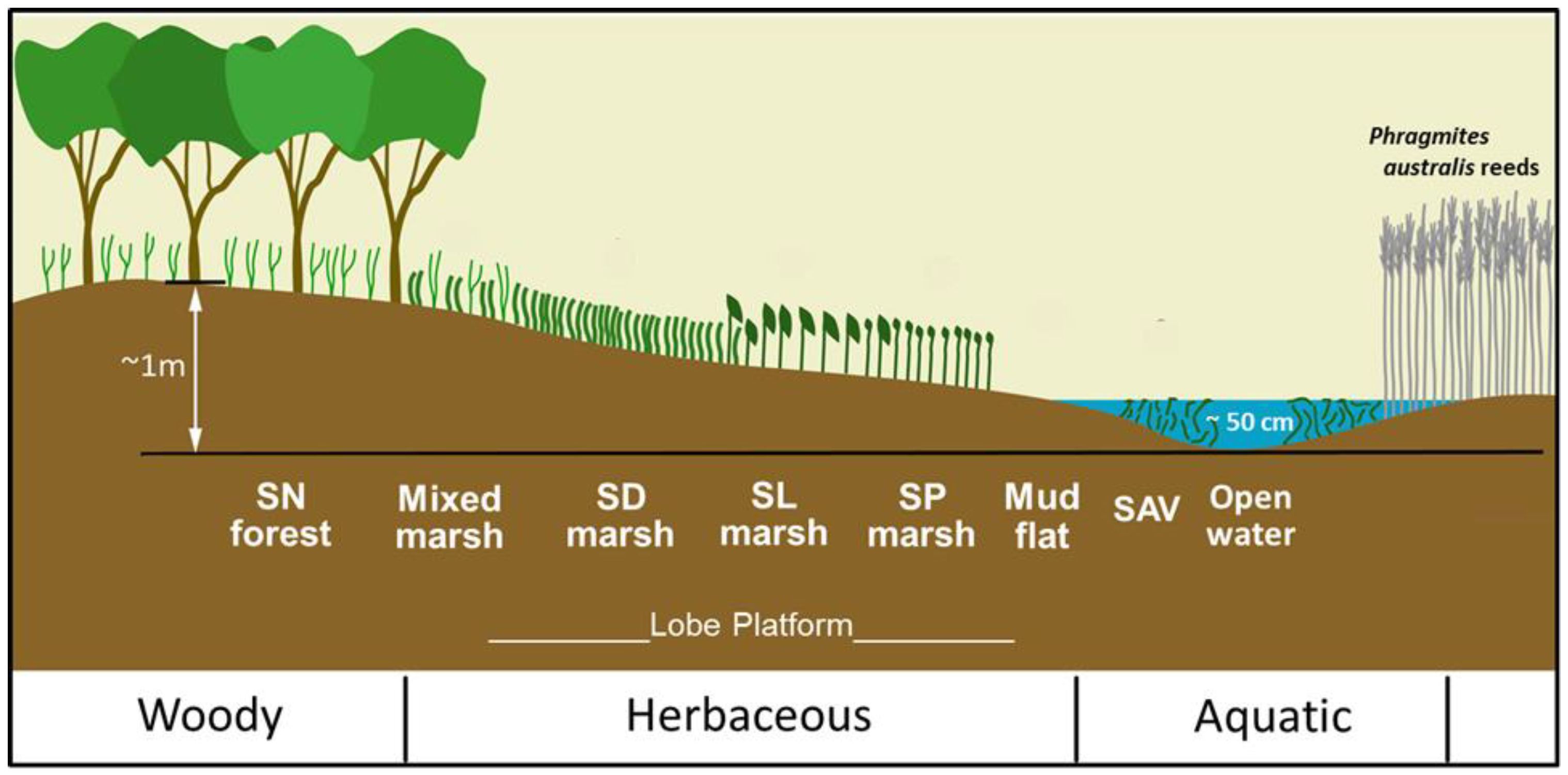
3.1. Splay Lobe General Vegetation
3.2. Phytosociology on Deltaic Lobes
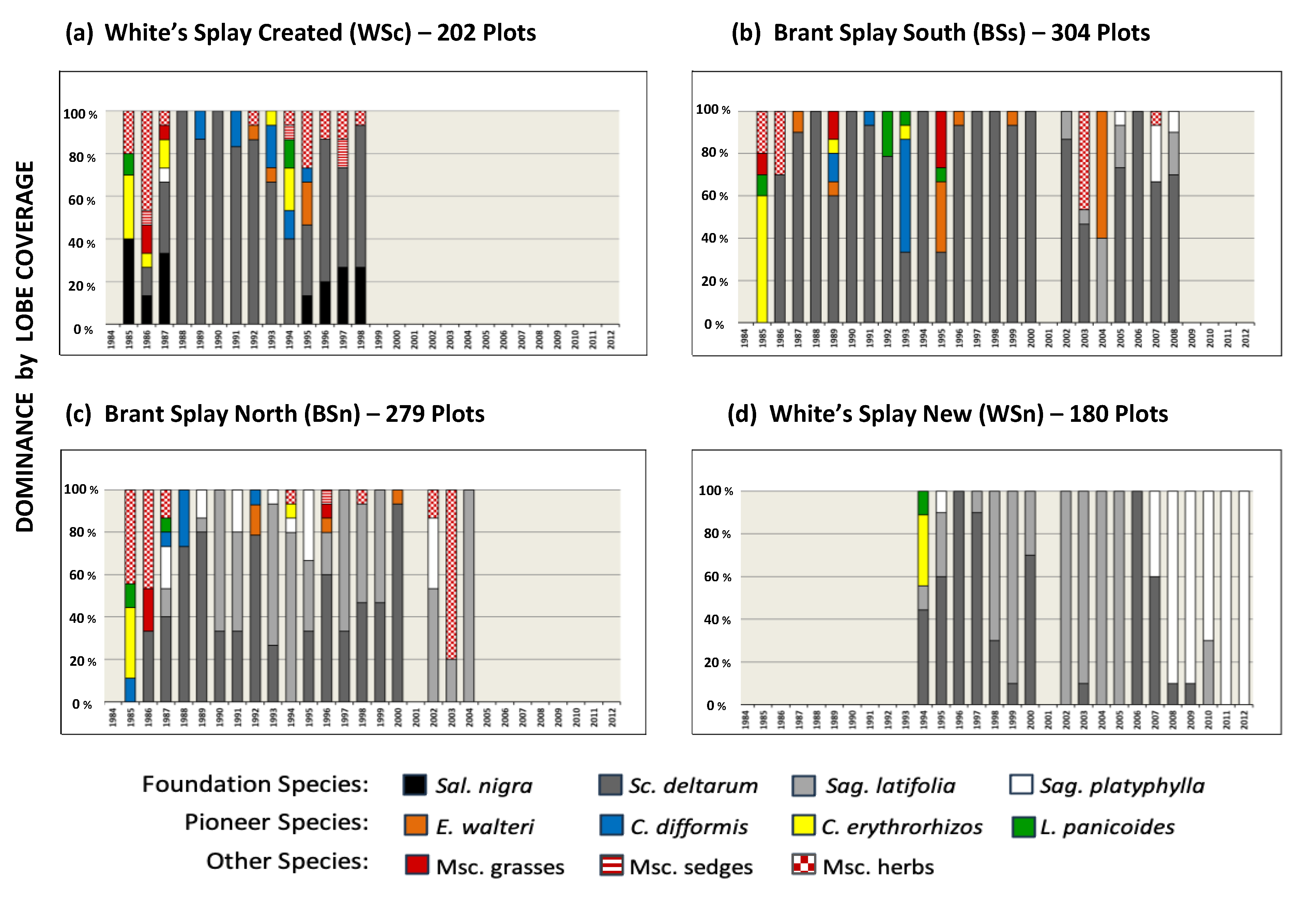
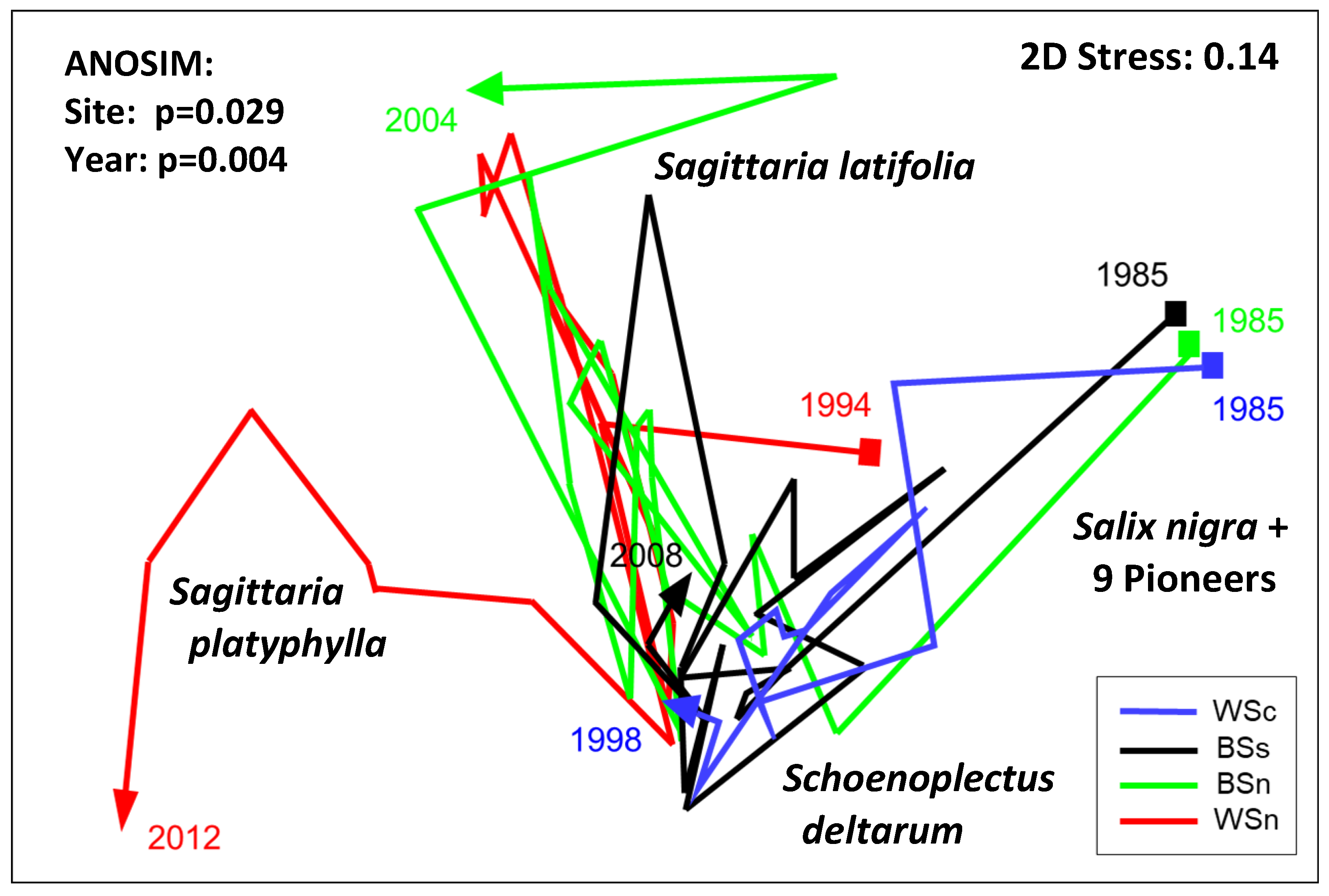
3.2.1. Plot Taxa Dominance over Time
3.2.2. Plot Integration over Time
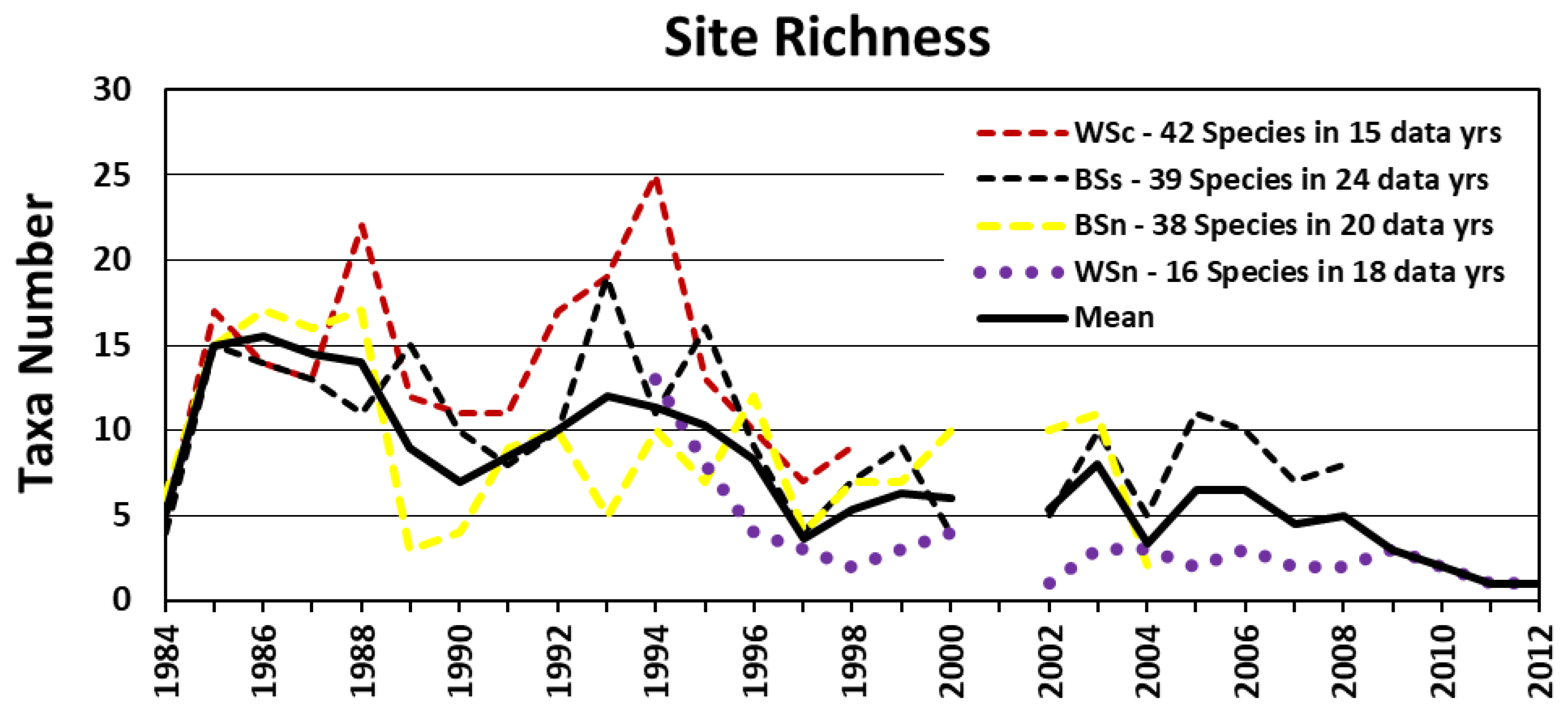
3.2.3. Lobe-Wide Richness Patterns
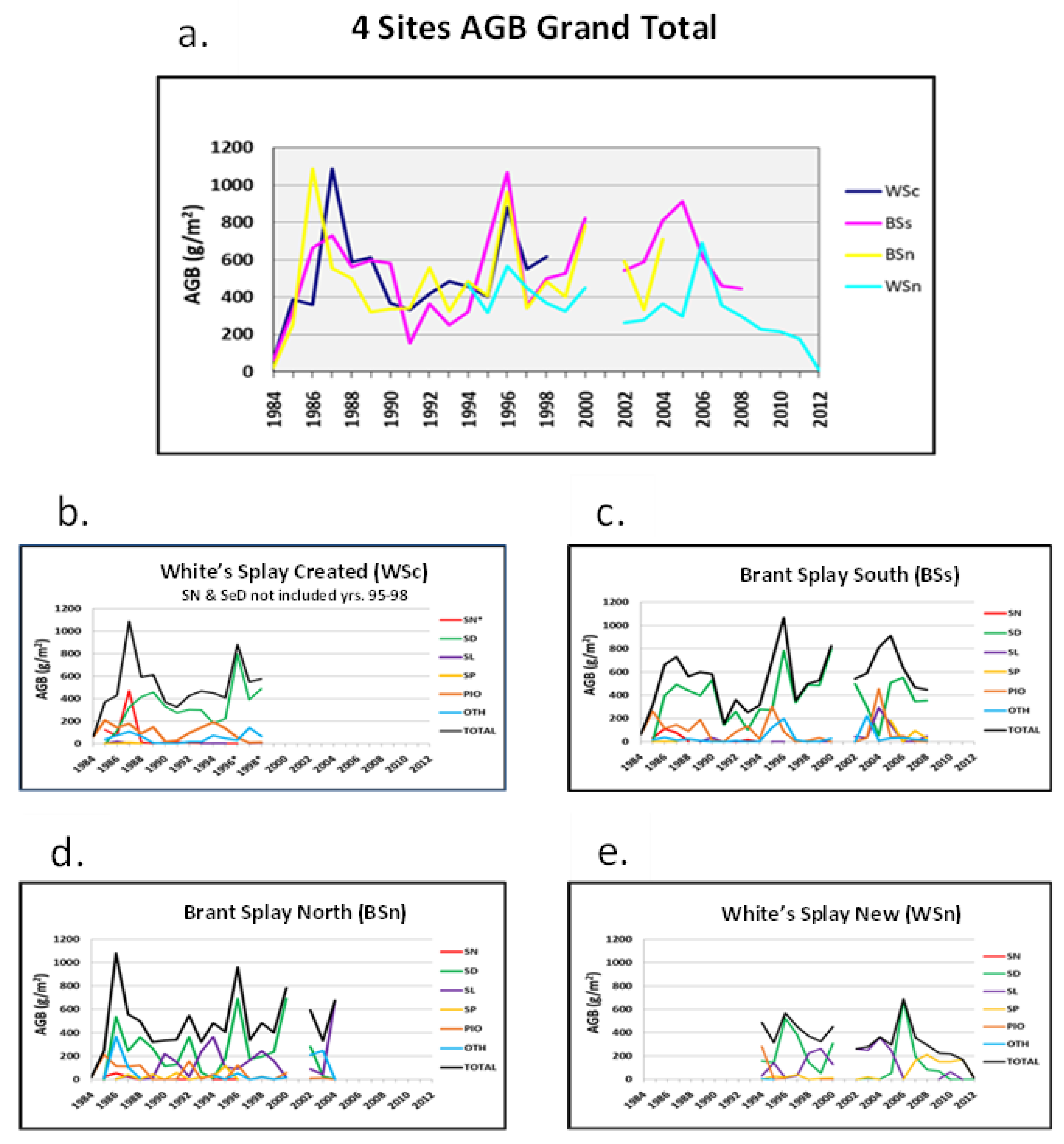
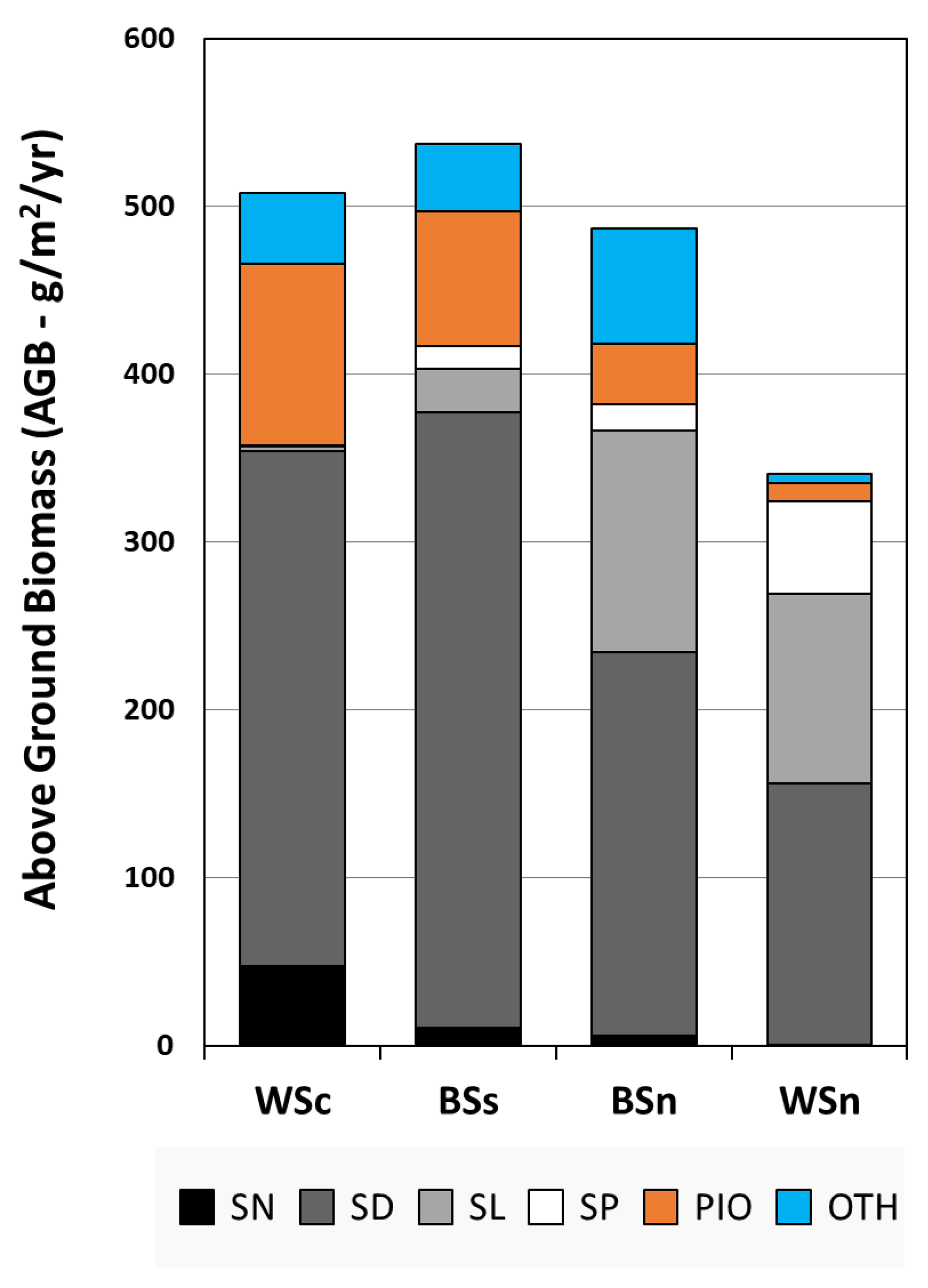
3.2.4. Lobe Total Biomass Patterns
3.2.5. Lobe Decadal Vegetation Patterns
- A highly significant correlation among the three most elevated sites (WSc, BSs, and BSn) when the three common PSYs were included. The one exception was for the 15 common harvest years between WSc and BSn—a period of no significance, although the AGB did show a correlation during the PPSY (i.e., after 1987);
- The lowest mean elevation site (WSn) paired with the other three sites never showed significance across the common years of study (Table 3);
- The most striking pattern of the annual total AGB values occurred in 1996, when all lobe sites experienced a sharp one-year increase in community biomass, particularly for the three most elevated sites (Figure 4a);
- Large single-year increases were common across all sites (in 1986 at BSn, 1987 at WSc, 2005 at BSs, and 2006 at WSn), with matched increases in other years (BSs–BSn in 2000, BSs–BSn in 2004), and likely through 2005 if BSn had been harvested (the channel access was blocked by more than 0.5 km of floating and aquatic vegetation). The yearly trends in the total harvested AGB followed the abundances of Sc. deltarum and Sag. latifolia (Figure 4b–e);
- An intrasite pattern was statistically substantiated. Over the full study period at BSn, an inverse relationship between the presence of Sc. deltarum and Sag. latifolia occurred (CC = −0.534; p < 0.05). At BSn, Sc. deltarum and the combined members of OTH species showed positive relationships over the study years (CC = 0.551; p < 0.05). An inverse significance was observed among FS at the WSn site: between Sc. deltarum and Sag. latifolia (CC = −0.545; p < 0.05) and at the same site between Sag. latifolia and Sag. platyphylla (CC = −0.584; p < 0.05; Figure 4d). Hints of this pattern were visible at WSn between Sc. deltarum and Sag. latifolia (Figure 4e) but not among herbaceous FS at WSc. Because WSc was the most elevated site, its construction likely had impacts, although Sc. deltarum was common at that site;
- Several FS experienced eruptions in particular years, such as the 1996 increase in Sc. deltarum at all sites and two large increases of the same species at the three sites that were then under study: BSs, BSn, and WSn in 2000 and BSs and WSn in 2006. In 2004, at the same sites, Sag. latifolia exhibited substantial increases. The fourth and woody FS, Sal. Nigra, showed a massive peak in abundance at WSc during the fourth year of study (1987, its first PPSY) and primarily as seedlings during all years of the study. During the PPSYs, Sal. nigra seedlings were harvested in only 15 plots of the 763 other sites and grew into young shading trees the last 4 years at WSc, along with the semi woody Se. drummondii (Figure 4). Figure 4 does not adequately depict the extensive diversity of understory woody to semi woody shrubs and the occasional other tree taxa.
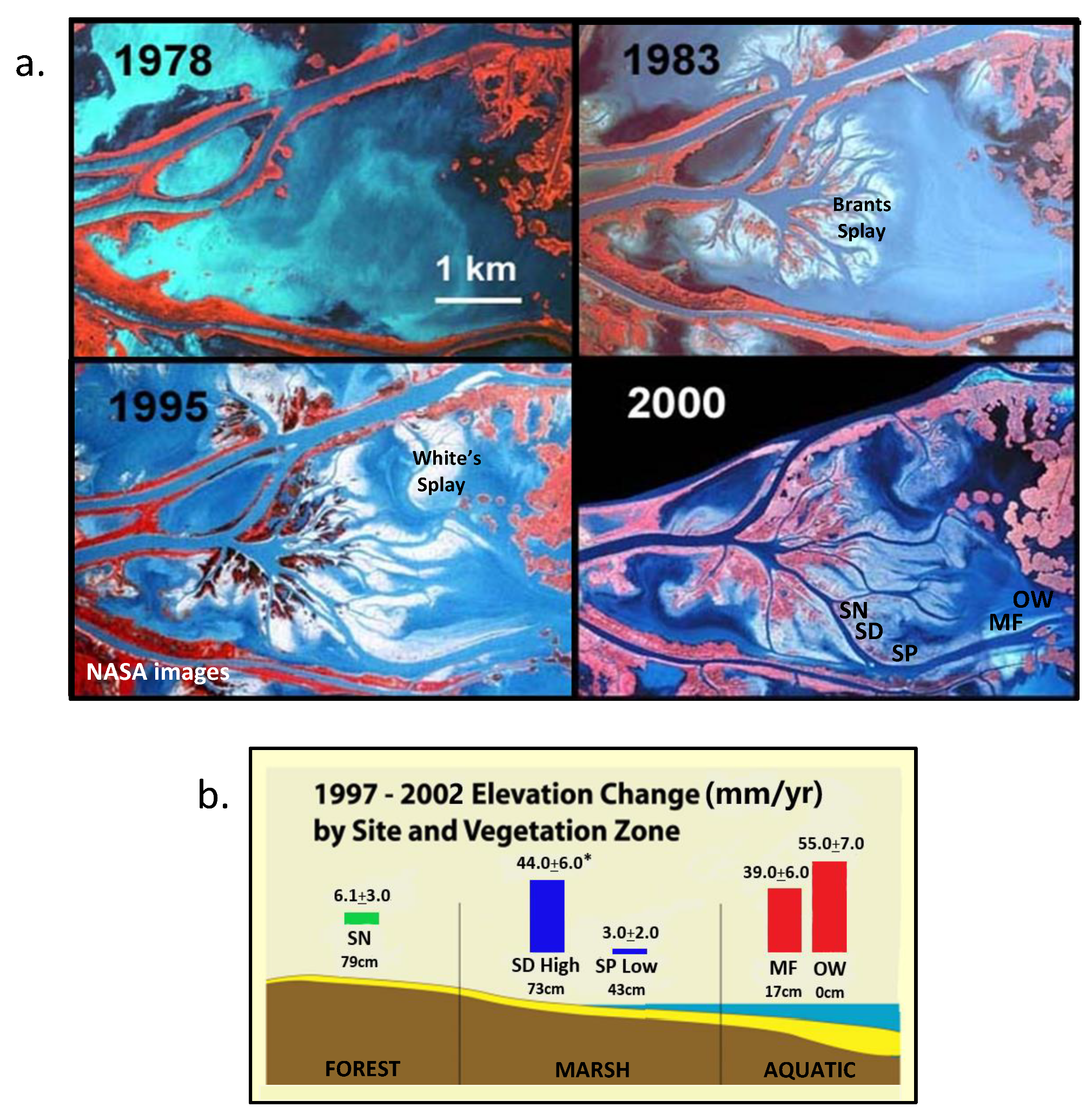
3.2.6. MRBD Splay Wetland Growth
3.2.7. River Quality and Wetland Impacts
4. Discussion
- Sediment pulses, which are instances of spotty deposition or erosion from small sediment pulse events, which create local patches of ±elevation changes, forcing vegetation changes [80]. They fit into the category of regeneration niches through the re-establishment process after a disturbance [81] and contributed considerably to the increase in the lobe-wide alpha diversity reported in this study, furthering lobe plant community dynamism;
- Vegetation sediment impact: Some areas were fashioned in more subtle ways than others in this study at the landscape scale by autogenic processes, that is, within the lobe ecosystem itself by the taxa forcing changes in sedimentation [82,83,84]. The clearest example of this did not appear in the sampled plots but in localized lobe areas where particular vegetation (especially tall, introduced haplotypes, mostly of Typha sp., but at times Ph. australis; D.W., pers. obs.) created pockets of substrate elevation differences, often resulting in areas available for secondary establishment and further impacting the wetland’s alpha diversity.
Author Contributions
Funding
Data Availability Statement
Acknowledgments

Conflicts of Interest
References
- Edmonds, D.A.; Caldwell, R.L.; Brondizio, E.S.; Siani, S.M.O. Coastal flooding will disproportionately impact people on river deltas. Nat. Commun. 2020, 11, 4741. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; Van Nostrand Reinhold: New York, NY, USA, 1986. [Google Scholar]
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, A.; Moora, M.; Pärtel, M.; et al. Ecological assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- National Research Council (NRC). Wetlands: Characteristics and Boundaries; National Academy Press: Washington, DC, USA, 1995. [Google Scholar]
- Fontaine, T.D.; Bartell, S.M. (Eds.) Dynamics of Lotic Ecosystems; Ann Arbor Science: Ann Arbor, MI, USA, 1983. [Google Scholar]
- Giosan, L.; Syvitski, J.; Constantinescu, S.; Day, J. 2Climate change: Protect the world’s deltas. Nature 2014, 516, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Tessler, Z.D.; Vörösmarty, C.J.; Grossberg, M.; Gladkova, I.; Aizenman, H.; Syvitski, J.P.M.; Foufoula–Georgiou, E. Profiling risk and sustainability in coastal deltas of the world. Science 2015, 349, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Stone, G.W.; Donley, J. The World Deltas Conference: A Tribute to the Late Professor J. Plummer Morgan: 1919–1995. J. Coastal Res. 1998, 14, 695–697. [Google Scholar]
- Day, J.W.; Boesch, D.F.; Clairain, E.J.; Kemp, G.P.; Laska, S.B.; Mitsch, W.J.; Orth, K.; Mashriqui, H.; Reed, D.J.; Shabman, L.; et al. Restoration of the Mississippi Delta: Lessons from hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar] [CrossRef]
- Nienhuis, J.H.; Ashton, A.D.; Edmonds, D.A.; Hoitink, A.J.F.; Kettner, A.J.; Rowland, J.C.; Törnqvist, T.E. Global–scale human impact on delta morphology has led to net land area gain. Nature 2020, 577, 514–518. [Google Scholar] [CrossRef]
- Tao, B.; Tian, H.; Ren, W.; Yang, J.; Yang, Q.; He, R.; Cai, W.; Lohrenz, S. Increasing Mississippi river discharge throughout the 21st century influenced by changes in climate, land use, and atmospheric CO2. Geophys. Res. Lett. 2014, 41, 4978–4986. [Google Scholar] [CrossRef]
- Coleman, J.M.; Roberts, H.H.; Stone, G.W. Mississippi River delta: An overview. J. Coastal Res. 1998, 14, 699–716. [Google Scholar]
- Batker, D.; de la Torre, I.; Costanza, R.; Day, J.W.; Swedeen, P.; Boumans, R.; Bagstad, K. The threats to the value of ecosystem goods and services of the Mississippi delta. In Perspectives on the Restoration of the Mississippi Delta; Day, J., Kemp, J.G., Freeman, A., Muth, D., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 55–173. [Google Scholar] [CrossRef]
- Day, J.W.; Kemp, J.G.; Freeman, A.; Muth, D. Introduction: Perspectives on the restoration of the Mississippi Delta. In Perspectives on the Restoration of the Mississippi Delta; Day, J., Kemp, J.G., Freeman, A., Muth, D., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 1–7. [Google Scholar] [CrossRef]
- Martin, J.F.; Reyes, E.; Kemp, G.P.; Mashriqui, H.; Day, J.W. Landscape modeling of the Mississippi delta: Using a series of landscape models, we examined the survival and creation of Mississippi Delta marshes and the impact of altered riverine inputs, accelerated sea–level rise, and management proposals on these marshes. BioScience 2002, 52, 357–365. [Google Scholar] [CrossRef]
- Roberts, H.H. Dynamic changes of the Holocene Mississippi River delta plain: The delta cycle. J. Coastal Res. 1997, 13, 605–627. [Google Scholar]
- Sasser, C.E.; Gosselink, J.G.; Holm, G.O.; Visser, J.M. Tidal freshwater wetlands of the Mississippi River deltas. In Tidal Freshwater Wetlands; Barendregt, A., Whigham, D., Baldwin, A., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2009; pp. 167–178. [Google Scholar]
- Blum, M.D.; Roberts, H.H. The Mississippi delta region: Past, present, and future. Annu. Rev. Earth Planet. Sci. 2012, 40, 655–683. [Google Scholar] [CrossRef]
- Cahoon, D.R.; White, D.A.; Lynch, J.C. Sediment infilling and wetland formation dynamics in an active crevasse splay of the Mississippi River delta. Geomorphology 2011, 131, 57–68. [Google Scholar] [CrossRef]
- Barbour, M.G.; Burk, J.H.; Pitts, W.D. Terrestrial Plant Ecology; Benjamin/Cummings: Menlo Park, CA, USA, 1980. [Google Scholar]
- Gosselink, J.G. The Ecology of Delta Marshes of Coastal Louisiana: A Community Profile; National Coastal Ecosystems Team, Division of Biological Services, Research Development, Fish and Wildlife Service, US Department of the Interior: Washington, DC, USA, 1984. [Google Scholar]
- Van Der Valk, A.G. Succession in wetlands: A Gleasonian approach. Ecology 1981, 62, 688–696. [Google Scholar] [CrossRef]
- Gosselink, J.G.; Turner, R.E. The role of hydrology in freshwater wetland ecosystems. In Freshwater Wetlands. Ecological Processes and Management Potential; Good, R.E., Whigham, D.F., Simpson, R.L., Johnson, C.G., Eds.; Academic Press: New York, NY, USA, 1978; pp. 63–78. [Google Scholar]
- Day, J.W.; Pont, D.; Hensel, P.F.; Ibañez, C. Impacts of sea-level rise on deltas in the Gulf of Mexico and the Mediterranean: The importance of pulsing events to sustainability. Estuaries 1995, 18, 636–647. [Google Scholar] [CrossRef]
- Middleton, B.A. Wetland Restoration, Flood Pulsing, and Disturbance Dynamics; J. Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Gould, H.R.; McFarlan, E. Geologic history of the Chenier Plain, southwestern Louisiana. Gulf Coast Assoc. Geol. Soc. Trans. 1959, 9, 261–270. [Google Scholar]
- White, D.A. The geological and natural history of coastal Louisiana from the perspective of a plant landscape ecologist—And a lover of plants. In Proceedings of the Botanical Society of America Annual Meeting, New Orleans, LA, USA, 27 July–1 August 2013. [Google Scholar]
- Frazier, D.E. Recent deltaic deposits of the Mississippi River: Their development and chronology. Gulf Coast Assoc. Geol. Soc. Trans. 1967, 17, 287–315. [Google Scholar]
- Van Heerden, I.L.L.; Roberts, H.H. Atchafalaya Delta—Louisiana’s New Prograding Coast. AAPG Bull. 1980, 64, 1568. [Google Scholar]
- Johnson, W.B.; Sasser, C.E.; Gosselink, J.G. Succession of vegetation in an evolving river delta, Atchafalaya Bay, Louisiana. J. Ecol. 1985, 73, 973–986. [Google Scholar] [CrossRef]
- Wells, J.T.; Coleman, J.M. Wetland loss and the subdelta life cycle. Estuar. Coast. Shelf Sci. 1987, 25, 111–125. [Google Scholar] [CrossRef]
- Rejmánek, M.; Sasser, C.E.; Gosselink, J.G. Modeling of vegetation dynamics in the Mississippi River deltaic plain. Vegetatio 1987, 69, 133–140. [Google Scholar] [CrossRef]
- Roberts, H.H.; Adams, R.D.; Cunningham, R.H.W. Evolution of the sand–dominated subaerial phase, Atchafalaya Delta, Louisiana. AAPG Bull. 1980, 64, 264–279. [Google Scholar] [CrossRef]
- White, D.A. Vascular plant community development on mudflats in the Mississippi River delta, Louisiana, USA. Aquat. Bot. 1993, 45, 171–194. [Google Scholar] [CrossRef]
- Boesch, D.F.; Josselyn, M.N.; Mehta, A.J.; Morris, J.T.; Nuttle, W.K.; Simenstad, C.A.; Swift, D.J.P. Scientific assessment of coastal wetland loss, restoration and management in Louisiana. J. Coastal Res. 1994, SI 20, i-103. [Google Scholar]
- Boyer, M.E.; Harris, J.O.; Turner, R.E. Constructed crevasses and land gain in the Mississippi River Delta. Restor. Ecol. 1997, 5, 85–92. [Google Scholar] [CrossRef]
- Cahoon, D.R.; Marin, P.E.; Black, B.K.; Lynch, J.C. A method for measuring vertical accretion, elevation, and compaction of soft, shallow-water sediments. J. Sediment. Res. 2000, 70, 1250–1253. [Google Scholar] [CrossRef]
- O’Neil, T. The Muskrat in the Louisiana Coastal Marshes; Louisiana Department of Wildlife and Fisheries: New Orleans, LA, USA, 1949. [Google Scholar]
- Angelini, C.; Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. BioScience 2011, 61, 782–789. [Google Scholar] [CrossRef]
- Schöb, C.; Butterfield, B.J.; Pugnaire, F.I. Foundation species influence trait-based community assembly. New Phytol. 2012, 196, 824–834. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, R.K. Permanova for Primer: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Gabler, C.A. Modeling the Growth of a Louisiana Wetland: The Dynamics of the Brant Pass Splay Area. Bachelor’s Honors Thesis, Loyola University, New Orleans, LA, USA, 2004. [Google Scholar]
- Visser, J.M.; Sasser, C.E.; Cade, B.S. The effect of multiple stressors on salt marsh end–of–season biomass. Estuaries Coasts 2006, 29, 328–339. [Google Scholar] [CrossRef]
- Prentice, I.C.; Cramer, W.; Harrison, S.P.; Leemans, R.; Monserud, R.A.; Solomon, A.M. Special paper: A global biome model based on plant physiology and dominance, soil properties and climate. J. Biogeogr. 1992, 19, 117–134. [Google Scholar] [CrossRef]
- White, D.A.; Visser, J.M. Water quality change in the Mississippi River, including a warming river, explains decades of wetland plant biomass change within its Balize delta. Aquat. Bot. 2016, 132, 5–11. [Google Scholar] [CrossRef]
- Bevington, A.E.; Twilley, R.R. Island edge morphodynamics along a chronosequence in a prograding deltaic floodplain wetland. J. Coastal Res. 2018, 34, 806–817. [Google Scholar] [CrossRef]
- Nyman, J.A. Integrating successional ecology and the delta lobe cycle in wetland research and restoration. Estuaries Coasts 2014, 37, 1490–1505. [Google Scholar] [CrossRef]
- Wright, L.D. River Deltas. In Coastal Sedimentary Environments; Davis, R.A., Ed.; Springer: New York, NY, USA, 1978; pp. 5–68. [Google Scholar]
- Olliver, E.A.; Edmonds, D.A. Defining the ecogeomorphic succession of land building for freshwater, intertidal wetlands in Wax Lake Delta, Louisiana. Estuar. Coast. Shelf Sci. 2017, 196, 45–57. [Google Scholar] [CrossRef]
- Olliver, E.A.; Edmonds, D.A.; Shaw, J.B. Influence of floods, tides, and vegetation on sediment retention in Wax Lake Delta, Louisiana, USA. J. Geophys. Res. Earth Surface 2020, 125, e2019JF005316. [Google Scholar] [CrossRef]
- Twilley, R.R.; Day, J.W.; Bevington, A.E.; Castaneda-Moya, E.; Christensen, A.; Holm, G.; Heffner, L.R.; Lane, R.; McCall, A.; Aarons, A.; et al. Ecomorphology of coastal deltaic floodplains and estuaries in an active delta: Insights form the Atchafalaya Coastal Basin. Est. Coast. Shelf Sci. 2019, 227, 106341. [Google Scholar] [CrossRef]
- Kulp, M.; Penland, S.; Williams, S.J.; Jenkins, C.; Flocks, J.; Kindinger, J. Geologic framework, evolution, and sediment resources for restoration of the Louisiana coastal zone. J. Coastal Res. 2005, 44, 56–71. [Google Scholar]
- Shaffer, G.P.; Sasser, C.E.; Gosselink, J.G.; Rejmánek, M. Vegetation dynamics in the emerging Atchafalaya Delta, Louisiana, USA. J. Ecol. 1992, 80, 677–687. [Google Scholar] [CrossRef]
- Morton, R.A.; Barras, J.A. Hurricane impacts on coastal wetlands: A half–century record of storm–generated features from southern Louisiana. J. Coastal Res. 2011, 27, 27–43. [Google Scholar] [CrossRef]
- Cahoon, D.R. A review of major storm impacts on coastal wetland elevations. Estuaries Coasts 2006, 29, 889–898. [Google Scholar] [CrossRef]
- Jankowski, K.L.; Törnqvist, T.E.; Fernandes, A.M. Vulnerability of Louisiana’s coastal wetlands to present–day rates of relative sea–level rise. Nat. Commun. 2017, 8, 14792. [Google Scholar] [CrossRef]
- Evers, D.E.; Sasser, C.E.; Gosselink, J.G.; Fuller, D.A.; Visser, J.M. The impact of vertebrate herbivores on wetland vegetation in Atchafalaya Bay, Louisiana. Estuaries 1998, 21, 1–13. [Google Scholar] [CrossRef]
- Bevington, A.E.; Twilley, R.R.; Sasser, C.E. Deltaic floodplain wetland vegetation dynamics along the sediment surface elevation gradient and in response to disturbance from river flooding and hurricanes in Wax Lake Delta, Louisiana, USA. Geomorphology 2022, 398, 108011. [Google Scholar] [CrossRef]
- Day, J.; Lane, R.; Moerschbaecher, M.; DeLaune, R.; Mendelssohn, I.; Baustian, J.; Twilley, R. Vegetation and soil dynamics of a Louisiana estuary receiving pulsed Mississippi River water following Hurricane Katrina. Estuaries Coasts 2013, 36, 665–682. [Google Scholar] [CrossRef]
- Day, J.W.; Christian, R.R.; Boesch, D.M.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L. Consequences of climate change on the ecogeomorphology of coastal wetlands. Estuaries Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Carle, M.V.; Sasser, C.E.; Roberts, H.H. Accretion and vegetation community change in the Wax Lake Delta following the historic 2011 Mississippi River flood. J. Coastal Res. 2015, 31, 569–587. [Google Scholar] [CrossRef]
- Brinson, M.M.; Christian, R.R.; Blum, L.K. Multiple states in the sea-level induced transition from terrestrial forest to estuary. Estuaries 1995, 18, 648–659. [Google Scholar] [CrossRef]
- Jentsch, A.; White, P. A theory of pulse dynamics and disturbance in ecology. Ecology 2019, 100, e02734. [Google Scholar] [CrossRef]
- White, D.A.; Hauber, D.P.; Hood, C.S. Clonal differences in Phragmites australis from the Mississippi River delta. Southeast. Nat. 2004, 3, 531–544. [Google Scholar] [CrossRef]
- Morton, R.A.; Bernier, J.C. Recent subsidence–rate reductions in the Mississippi Delta and their geological implications. J. Coastal Res. 2010, 26, 555–561. [Google Scholar] [CrossRef]
- Montz, G.N. Vegetational characteristics of the Atchafalaya River delta [Louisiana]. Proc. Louisiana Acad. Sci. 1978, 41, 71–84. [Google Scholar]
- Weaver, J.E.; Clements, F.E. Plant Ecology; McGraw Hill: New York, NY, USA, 1983. [Google Scholar]
- Jensen, D.; Cavanaugh, K.C.; Simard, M.; Christensen, A.; Rovai, A.; Twilley, R. Aboveground biomass distributions and vegetation composition changes in Louisiana’s Wax Lake Delta. Estuar. Coast. Shelf Sci. 2021, 250, 107139. [Google Scholar] [CrossRef]
- Gough, L.; Grace, J.B. Effects of flooding, salinity and herbivory on coastal plant communities, Louisiana, United States. Oecologia 1998, 117, 527–535. [Google Scholar] [CrossRef]
- Howard, R.J.; Mendelssohn, I.A. Salinity as a constraint on growth of oligohaline marsh macrophytes. I. Species variation in stress tolerance. Am. J. Bot. 1998, 86, 785–794. [Google Scholar] [CrossRef]
- Holm, G.O.; Sasser, C.E. Differential salinity response between two Mississippi River subdeltas: Implications for changes in plant composition. Estuaries 2001, 24, 78–89. [Google Scholar] [CrossRef]
- Steyer, G.D.; Perez, B.C.; Piazza, S.; Suir, G. Potential consequences of saltwater intrusion associated with Hurricanes Katrina and Rita. In Science and the Storms: The USGS Response to the Hurricanes of 2005; United States Geological Survey: Reston, VA, USA, 2007; pp. 137–146. [Google Scholar] [CrossRef]
- Fuller, D.A.; Sasser, C.E.; Johnson, W.B.; Gosselink, J.G. The effects of herbivory on vegetation on islands in Atchafalaya Bay, Louisiana. Wetlands 1984, 4, 105–114. [Google Scholar] [CrossRef]
- Taylor, K.L.; Grace, J.B. The effects of vertebrate herbivory on plant community structure in the coastal marshes of the Pearl River, Louisiana, USA. Wetlands 1995, 15, 68–73. [Google Scholar] [CrossRef]
- Geho, E.M.; Campbell, D.; Keddy, P.A. Quantifying ecological filters: The relative impact of herbivory, neighbors, and sediment on an oligohaline marsh. Oikos 2007, 116, 1006–1016. [Google Scholar] [CrossRef]
- White, P.S.; Pickett, S.T.A. Natural disturbances and patch dynamics: An introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 3–13. [Google Scholar]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1997, 52, 107–145. [Google Scholar] [CrossRef]
- Van Dijk, W.M.; Teske, R.; Van de Lageweg, W.I.; Kleinhans, M.G. Effects of vegetation distribution on experimental river channel dynamics. Water Resour. Res. 2013, 49, 7558–7574. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Nyman, J.A.; Walters, R.J.; Delaune, R.D.; Patrick, W.H., Jr. Marsh vertical accretion via vegetative growth. Estuar. Coast. Shelf Sci. 2006, 69, 370–380. [Google Scholar] [CrossRef]
- Piliouras, A.; Kim, W. Upstream and downstream boundary conditions control the physical and biological development of river deltas. Geophys. Res. Lett. 2019, 46, 11188–11196. [Google Scholar] [CrossRef]
- Piliouras, A.; Kim, W. Delta size and plant patchiness as controls on channel network organization in experimental deltas. Earth Surf. Process. Landf. 2019, 44, 259–272. [Google Scholar] [CrossRef]
- Pollock, M.M.; Naiman, R.J.; Hanley, T.A. Plant species richness in riparian wetlands—A test of biodiversity theory. Ecology 1998, 79, 94–105. [Google Scholar] [CrossRef]
- Guan, B.; Chen, M.; Elsey-Quirk, T.; Yang, S.; Shang, W.; Li, Y.; Tian, X.; Han, G. Soil seed bank and vegetation differences following channel diversion in the Yellow River Delta. Sci. Total Environ. 2019, 693, 133600. [Google Scholar] [CrossRef]
- Carle, M.V.; Sasser, C.E. Productivity and resilience: Long-term trends and storm-driven fluctuations in the plant community of the accreting Wax Lake Delta. Estuaries Coasts 2016, 39, 406–422. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1971; Volume 3. [Google Scholar]
- Shew, D.M.; Linthurst, R.A.; Seneca, E.D. Comparison of production computation methods in a southeastern North Carolina Spartina alterniflora salt marsh. Estuaries 1981, 4, 97–109. [Google Scholar] [CrossRef]
- Good, R.E.; Whigham, D.F.; Simpson, R.L.; Johnson, C.G. Freshwater Wetlands. Ecological Processes and Management Potential; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Allen, Y.C.; Couvillion, B.R.; Barras, J.A. Using multitemporal remote sensing imagery and inundation measures to improve land change estimates in coastal wetlands. Estuaries Coasts 2012, 35, 190–200. [Google Scholar] [CrossRef]
- Neill, C.; Deegan, L.A. The effect of Mississippi River delta lobe development on the habitat composition and diversity of Louisiana coastal wetlands. Am. Midl. Nat. 1986, 116, 296–303. [Google Scholar] [CrossRef]
- Morris, J.T.; Barber, D.C.; Callaway, J.C.; Chambers, R.; Hagen, S.C.; Hopkinson, C.S.; Johnson, B.J.; Megonigal, P.; Neubauer, S.S.; Troxler, T.; et al. Contributions of organic and inorganic matter to sediment volume and accretion in tidal wetlands at steady state. Earth’s Future 2016, 4, 110–121. [Google Scholar] [CrossRef]
- Osland, M.J.; Gabler, C.A.; Grace, J.B.; Day, R.H.; McCoy, M.L.; McLeod, J.L.; From, A.S.; Enwright, N.N.; Feher, L.C.; Stagg, C.L.; et al. Climate and plant controls on soil organic matter in coastal wetlands. Global Change Biol. 2018, 24, 5361–5379. [Google Scholar] [CrossRef]
- Törnqvist, T.E.; Cahoon, D.R.; Morris, J.T.; Day, J.W. Coastal wetland resilience, accelerated sea-level rise, and the importance of timescale. AGU Adv. 2021, 2, e2020AV000334. [Google Scholar] [CrossRef]
- Corenblit, D.; Baas, A.C.W.; Bornette, G.; Darrozes, J.; Delmotte, S.; Francis, R.A.; Gurnell, A.M.; Julien, F.; Naiman, R.J.; Steiger, J. Feedbacks between geomorphology and biota controlling earth surface processes and landforms: A review of foundation concepts and current understandings. Earth-Sci. Rev. 2011, 106, 307–331. [Google Scholar] [CrossRef]
- Marchetti, Z.Y.; Villalba, A.B.; Ramonell, C.; Brumnich, F.; Pereira, M.S. Biogeomorphic succession in a fluvial–lacustrine delta of the Middle Paraná River (Argentina): Feedbacks between vegetation and morphodynamics. Sci. Total Environ. 2020, 739, 139799. [Google Scholar] [CrossRef]
- Coleman, J.M. Dynamic changes and processes in the Mississippi River delta. Geol. Soc. Am. Bull. 1988, 100, 999–1015. [Google Scholar] [CrossRef]
- Rogers, K.; Zawadzki, A.; Mogensen, L.A.; Saintilan, N. Coastal wetland surface elevation change is dynamically related to accommodation space and influenced by sedimentation and sea–level rise over decadal timescales. Front. Mar. Sci. 2022, 9, 807588. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Allison, M.A. Large-river delta-front estuaries as natural recorders of global environmental change. PNAS Sci. 2009, 106, 8085–8092. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of diversity. Nature. 1949, 163, 688. [Google Scholar] [CrossRef]
- Berger, W.H.; Parker, F.L. Diversity of planktonic foraminifera in deep-sea sediments. Science. 1970, 168, 1345–1347. [Google Scholar] [CrossRef]
- Frans, C.; Istanbulluoglu, E.; Mishra, V.; Munoz-Arriola, F.; Lettenmaier, D.P. Are climatic or land cover changes the dominant cause of runoff trends in the Upper Mississippi River Basin? Geophys. Res. Lett. 2013, 40, 1104–1110. [Google Scholar] [CrossRef]
- Syvitski, J.P.M.; Kettner, A. Sediment flux and the Anthropocene. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 957–975. [Google Scholar] [CrossRef]
- Glick, P.; Clough, J.; Polaczyk, A.; Couvillion, B.; Nunley, B. Potential effects of sea-level rise on coastal wetlands in southeastern Louisiana. J. Coast. Res. 2013, 63, 211–233. [Google Scholar] [CrossRef]
- Gudmundsson, L.; Boulange, J.; Do, H.X.; Gosling, S.N.; Grillakis, M.G.; Koutroulis, A.G.; Leonard, M.; Liu, J.; Müller Schmeid, H.; Papadimitriou, L.; et al. Globally observed trends in mean and extreme river flow attributed to climate change. Science 2021, 371, 1159–1162. [Google Scholar] [CrossRef]
- Collins, S.L. Climate Change and Long-Term Ecological Research. BioScience. 2022, 72, 803. [Google Scholar] [CrossRef]
- Hughes, B.B.; Beas–Luna, R.; Barner, A.K.; Brewitt, A.; Brumbaugh, D.R.; Cerny-Chipman, E.B.; Close, S.L.; Coblentz, K.E.; de Nesnera, K.L.; Drobnitch, S.T.; et al. Long–term studies contribute disproportionately to ecology and policy. BioScience 2017, 67, 271–281. [Google Scholar] [CrossRef]
- Köhler, J. Long–term ecological research in freshwater ecosystems. In Long–Term Ecological Research: Between Theory and Application; Müller, F., Baessler, C., Schubert, H., Klötz, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 179–187. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Species | TAGB | Total | PTB | Total |
|---|---|---|---|---|---|
| Foundation Species (4 taxa) | Schoenoplectus deltarum (Sch.) Soják | 65,808 | 56.4% | ||
| Sagittaria latifolia Willd. | 18,334 | 15.7% | |||
| Sagittaria platyphylla (Engelm.) J.G. Sm. | 5191 | 4.4% | |||
| Salix nigra Marsh. | 2673 | 2.3% | |||
| 92,006 | 78.9% | ||||
| Pioneer Species (9 taxa) | Echinochloa walteri (Pursh) A. Heller * | 5566 | 4.8% | ||
| Cyperus difformis L. * | 3233 | 2.8% | |||
| Cyperus erythrorhizos Muhl. * | 2572 | 2.2% | |||
| Leptochloa panicoides (Presl) Hitchc. * | 1687 | 1.4% | |||
| Ammannia coccinea Rottb. | 806 | ||||
| Sphenoclea zeylanica Gaertn. | 752 | ||||
| Bacopa monnieri (L.) Pennell | 598 | ||||
| Panicum dichotomiflorum Michx. | 341 | ||||
| Eclipta prostrata (L.) L. | 301 | ||||
| 15,856 | 13.6% | ||||
| Other Species (40 taxa) | |||||
| Msc. Grasses (8 taxa, 32.8% of OTH) | Panicum spp. | 1272 | |||
| Msc. Poaceae ** | 1228 | ||||
| Panicum capillare L. | 136 | ||||
| Eragrostis hypnoides (Lam.) Britt., Sterns & Poggenb. | 95 | ||||
| Leersia sp. *** | 80 | ||||
| Paspalum fluitans (Elliott) Kunth | 29 | ||||
| Leptochloa fusca (L.) Kunth (fascicularis) | 14 | ||||
| Zizaniopsis miliacea (Michx.) Döll & Asch. | 5 | ||||
| 2859 | 2.5% | ||||
| Msc. Sedges (8 taxa, 8.0% of OTH) | Msc. Cyperaceae ** | 404 | |||
| Eleocharis spp. | 171 | ||||
| Juncus sp. *** | 41 | ||||
| Cyperus sp. *** | 33 | ||||
| Schoenoplectus tabernaemontani (C.C. Gmel.) Palla | 26 | ||||
| Cyperus surinamensis Rottb. | 10 | ||||
| Fimbristylis vahlii (Lam.) Link | 10 | ||||
| Juncus diffusissimus (Buckley) | 4 | ||||
| 699 | 0.6% | ||||
| Msc. Herbs (24 taxa, 59.2% of OTH) | Vigna luteola (Jacq.) Benth. | 1643 | 1.4% | ||
| Polygonum punctatum Elliott | 1054 | ||||
| Typha domingensis Pers. | 1022 | ||||
| Pluchea odorata (L.) Cass. | 357 | ||||
| Msc. Dicotyledons ** | 298 | ||||
| Lindernia dubia (L.) Pennell | 287 | ||||
| Sesbania drummondii (Rydb.) Cory | 151 | ||||
| Symphyotrichum sp. *** | 71 | ||||
| Colocasia esculenta (L.) Schott | 60 | ||||
| Ludwigia decurrens Walter | 36 | ||||
| Amaranthus australis (A. Gray) Sauer | 35 | ||||
| Iris sp. *** | 24 | ||||
| Polygonum lapathifolium L. | 20 | ||||
| Alternanthera philoxeroides (Mart.) Griseb. | 19 | ||||
| Rumex sp. *** | 19 | ||||
| Najas guadalupensis (Spreng.) Magnus | 17 | ||||
| Phytolacca sp. *** | 17 | ||||
| Hydrocotyle sp. *** | 11 | ||||
| Potomogeton sp. *** | 8 | ||||
| Ipomea sagittata Poir. | 5 | ||||
| Polygonum sp. *** | 3 | ||||
| Baccharis halimifolia L. | 2 | ||||
| Lythrum lineare L. | 2 | ||||
| Polygonum glabrum Willd. | 2 | ||||
| 5163 | 4.4% | ||||
| Species a | Group | p | Prop b | R | Abun c | R | Pres d | R | A + P e | Dom f | R | A + P + D g |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Schoenoplectus deltarum | FS | 0.001 | 0.2998 | 1 | 56.39 | 1 | 75.1 | 1 | 1 | 53.1 | 1 | 1 |
| Sagittaria latifolia | FS | 0.001 | 0.2845 | 2 | 15.71 | 2 | 34.9 | 2 | 2 | 18.7 | 2 | 2 |
| Sagittaria platyphylla | FS | 0.001 | 0.1285 | 3 | 4.45 | 4 | 9.2 | 8 | 5 | 8.2 | 3 | 5 |
| Leptochloa panicoides | PIO | 0.001 | 0.1123 | 4 | 1.45 | 8 | 5.9 | 9 | 7 h | 1.1 | 8 | 7 |
| Cyperus erythrorhizos | PIO | 0.001 | 0.1113 | 5 | 2.20 | 7 | 12.0 | 6 | 6 | 2.6 | 5 | 6 |
| Cyperus difformis | PIO | 0.001 | 0.0792 | 6 | 2.77 | 5 | 22.8 | 3 | 4 | 2.9 | 4 | 3 |
| Ammannia coccinea | PIO | 0.019 | 0.0427 | 7 | 0.69 | 12 | 13.2 | 5 | 8 h | 0.0 | 11 | 10 |
| Echinochloa walteri | PIO | 0.030 | 0.0382 | 8 | 4.77 | 3 | 14.0 | 4 | 3 | 2.5 | 6 | 4 |
| Polygonum punctatum | OTH | 0.034 | 0.0373 | 9 | 0.90 | 10 | 2.1 | 11 | 11 | 0.1 | 9 | 11 |
| Vigna luteola | OTH | 0.040 | 0.0346 | 10 | 1.41 | 9 | 4.3 | 10 | 9 | 0.1 | 10 | 8 |
| Sphenochlea zeylanica | PIO | 0.045 | 0.0315 | 11 | 0.64 | 13 | 9.3 | 7 | 10 | 1.5 | 7 | 9 |
| Period | Sites | Sampling Period | Years | Correlation Coefficient | p a |
|---|---|---|---|---|---|
| All b | WSc vs. BSs | 1984–1998 | 15 | 0.690 | 0.004 |
| WSc vs. BSn | 1984–1998 | 15 | 0.438 | 0.102 | |
| WSc vs. WSn | 1994–1998 | 5 | 0.743 | 0.150 | |
| BSs vs. BSn | 1984–1998 | 15 | 0.711 | 0.003 | |
| BSs vs. BSn | 1984–2004 | 20 | 0.728 | 0.003 | |
| BSs vs. WSn | 1994–2004 | 5 | 0.174 | 0.552 | |
| BSn vs. WSn | 1994–2004 | 5 | 0.580 | 0.079 | |
| PPSY | WSc vs. BSs | 1988–1998 | 11 | 0.709 | 0.015 |
| WSc vs. BSn | 1988–1998 | 11 | 0.725 | 0.012 | |
| BSs vs. BSn | 1988–1998 | 11 | 0.699 | 0.017 |
| Parameter | Total AGB | Sc. deltarum AGB | Sag. latifolia AGB |
|---|---|---|---|
| Average annual discharge (linear) | 0.1030 | 0.0882 | 0.0031 |
| Average annual discharge (quadratic) | 0.0322 | 0.0335 | 0.0194 |
| Spring discharge (linear) | <0.0001 | <0.0001 | 0.0171 |
| Spring discharge (quadratic) | <0.0001 | 0.0335 | 0.0793 |
| Nitrate loading (linear) | 0.0020 | 0.2160 | 0.8620 |
| Nitrate loading (quadratic) | 0.0006 | <0.0001 | 0.4198 |
| Growing season length | <0.0001 | 0.0208 | 0.0994 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, D.A.; Visser, J.M. Twenty-Eight Years of Plant Community Development and Dynamics in the Balize Mississippi River Delta, Louisiana, USA. Water 2023, 15, 3481. https://doi.org/10.3390/w15193481
White DA, Visser JM. Twenty-Eight Years of Plant Community Development and Dynamics in the Balize Mississippi River Delta, Louisiana, USA. Water. 2023; 15(19):3481. https://doi.org/10.3390/w15193481
Chicago/Turabian StyleWhite, David A., and Jenneke M. Visser. 2023. "Twenty-Eight Years of Plant Community Development and Dynamics in the Balize Mississippi River Delta, Louisiana, USA" Water 15, no. 19: 3481. https://doi.org/10.3390/w15193481
APA StyleWhite, D. A., & Visser, J. M. (2023). Twenty-Eight Years of Plant Community Development and Dynamics in the Balize Mississippi River Delta, Louisiana, USA. Water, 15(19), 3481. https://doi.org/10.3390/w15193481