
Hard-Bottom Polychaetes Exposed to Multiple Human Pressure along the Mediterranean Coast of Egypt

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
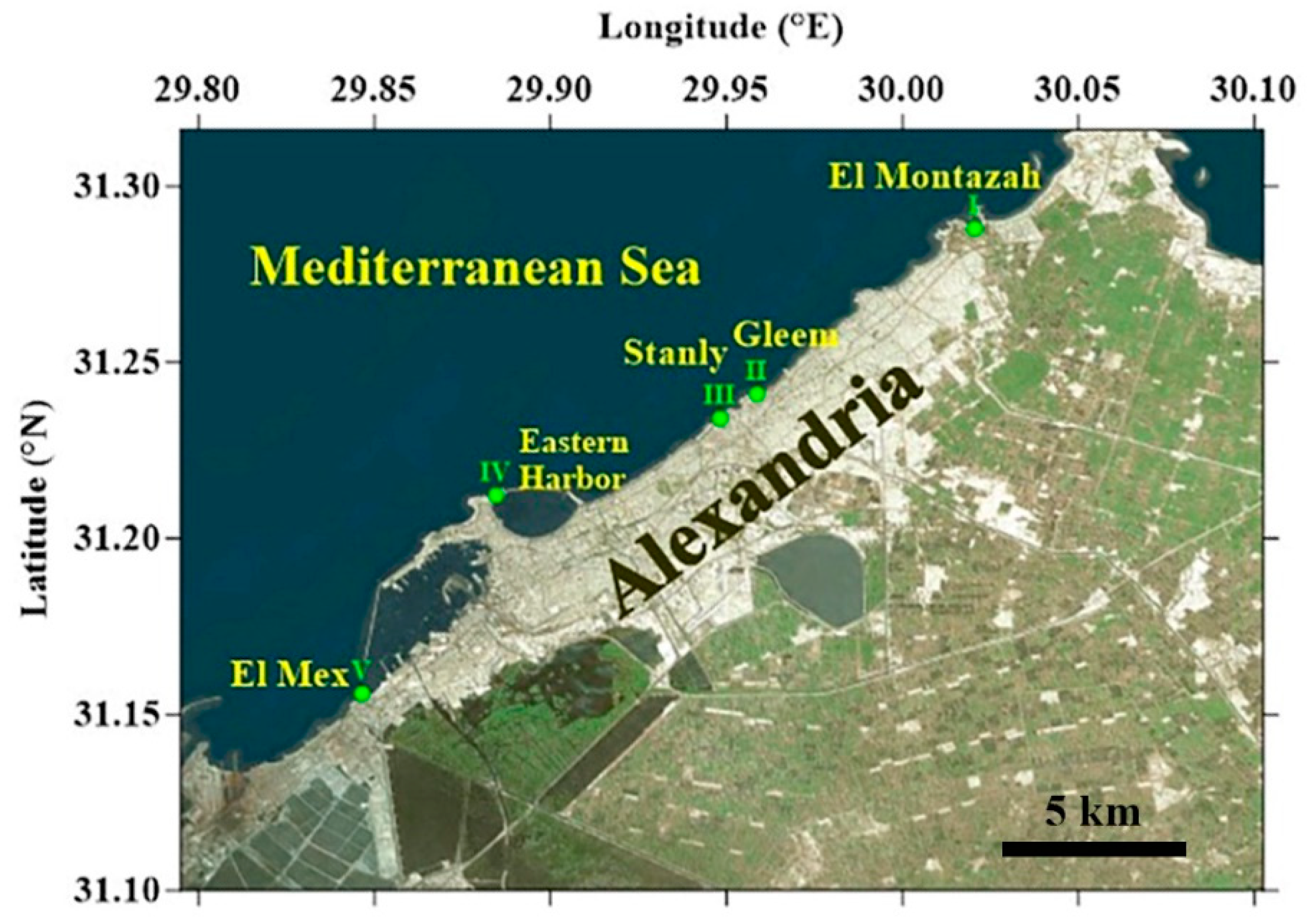
2.1. Study Area
2.2. Sampling Sites
2.3. Environmental Variables
2.4. Collection of Biotic Samples
2.5. Statistical Analysis
3. Results
3.1. Environmental Variables
3.2. Benthic Cover
3.3. Community Structure and Univariate Indexes
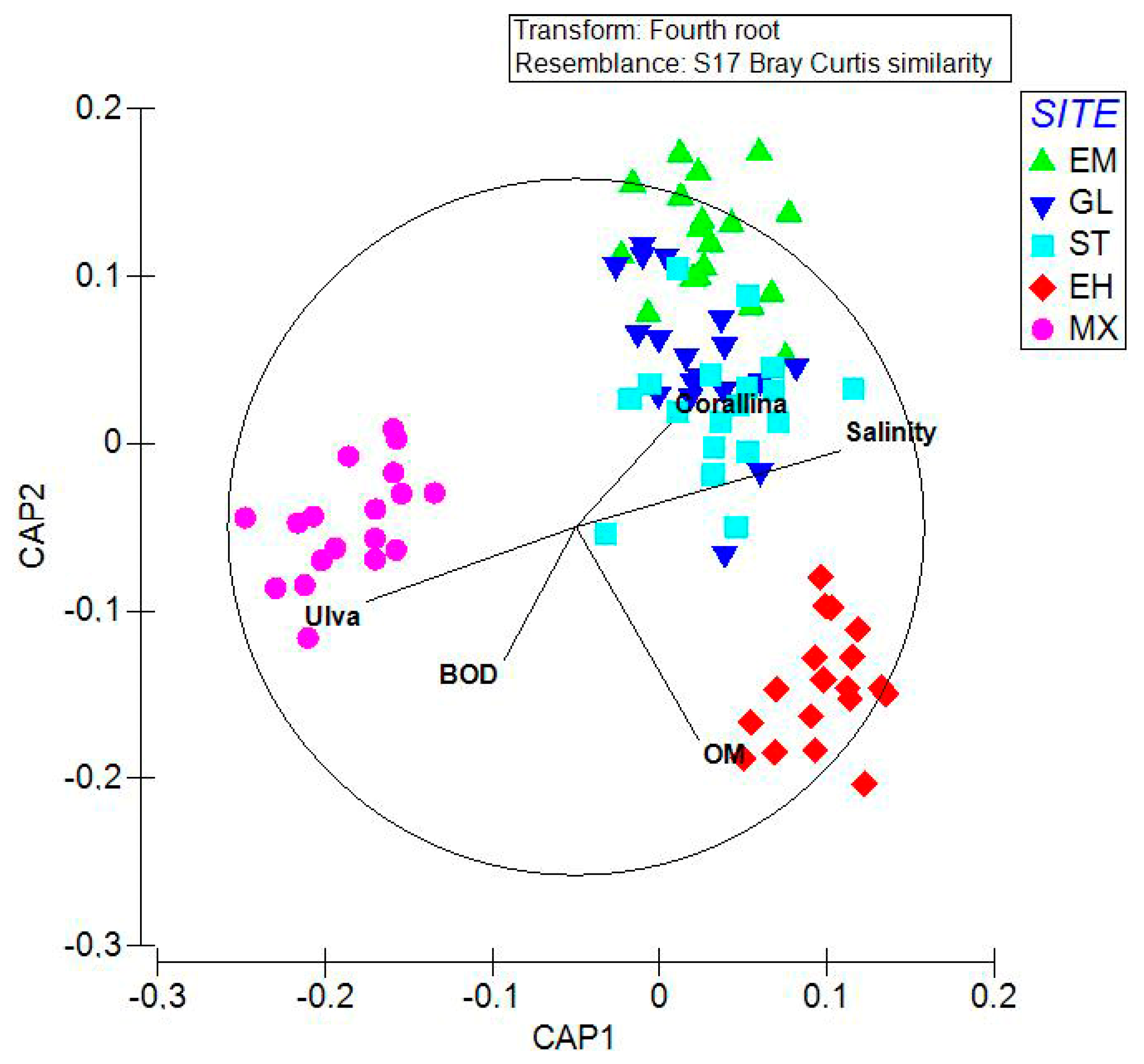
3.4. Statistical Analyses
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Myers, R.A.; Worm, B. Rapid worldwide depletion of predatory fish communities. Nature 2003, 423, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; Fujita, R. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Agardy, T. Global marine conservation policy versus site-level implementation. Mar. Ecol. Prog. Ser. 2005, 300, 242–248. [Google Scholar] [CrossRef]
- Spalding, M.D.; Ruffo, S.; Lacambra, C.; Meliane, I.; Hale, L.Z.; Shepard, C.C.; Beck, M.W. The role of ecosystems in coastal protection: Adapting to climate change and coastal hazards. Ocean. Coast. Manag. 2014, 90, 50–57. [Google Scholar] [CrossRef]
- Neumann, B.; Vafeidis, A.T.; Zimmermann, J.; Nicholls, R.J. Future coastal population growth and exposure to sea-level rise and coastal flooding-a global assessment. PLoS ONE 2015, 10, e0118571. [Google Scholar] [CrossRef] [Green Version]
- Halpern, B.S.; Selkoe, K.A.; Micheli, F.; Kappel, C.V. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 2007, 21, 1301–1315. [Google Scholar] [CrossRef]
- Fairweather, P.G. Sewage and the biota on seashores: Assessment of impact in relation to natural variability. Environ. Monit. Assess. 1990, 14, 197–210. [Google Scholar] [CrossRef]
- Otway, N.M.; Gray, C.A.; Craig, J.R.; McVea, T.A.; Ling, J.E. Assessing the impacts of deepwater sewage outfalls on spatially-and temporally-variable marine communities. Mar. Environ. Res. 1996, 41, 45–71. [Google Scholar] [CrossRef]
- Guidetti, P.; Terlizzi, A.; Fraschetti, S.; Boero, F. Changes in Mediterranean rocky-reef fish assemblages exposed to sewage pollution. Mar. Ecol. Prog. Ser. 2003, 253, 269–278. [Google Scholar] [CrossRef]
- Smith, A.K.; Ajani, P.A.; Roberts, D.E. Spatial and temporal variation in fish assemblages exposed to sewage and implications for management. Mar. Environ. Res. 1999, 47, 241–260. [Google Scholar] [CrossRef]
- Balestri, E.; Benedetti-Cecchi, L.; Lardicci, C. Variability in patterns of growth and morphology of Posidonia oceanica exposed to urban and industrial wastes: Contrasts with two reference locations. J. Exp. Mar. Biol. Ecol. 2004, 308, 1–21. [Google Scholar] [CrossRef]
- Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Anderson, M.J. Quantifying effects of pollution on biodiversity: A case study of highly diverse molluscan assemblages in the Mediterranean. Mar. Biol. 2005, 148, 293–305. [Google Scholar] [CrossRef]
- Terlizzi, A.; Benedetti-Cecchi, L.; Bevilacqua, S.; Fraschetti, S.; Guidetti, P.; Anderson, M.J. Multivariate and univariate asymmetrical analyses in environmental impact assessment: A case study of Mediterranean subtidal sessile assemblages. Mar. Ecol. Prog. Ser. 2005, 289, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; De Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Korajkic, A. Discrimination of Human and Non-Human Sources of Pollution in Gulf of Mexico Waters by Microbial Source Tracking Methods and the Investigation of the Influence of Environmental Factors on Escherichia coli Survival. Ph.D. Thesis, University of South Florida, Tampa, FL, USA, 2010. [Google Scholar]
- Bertocci, I.; Dell’Anno, A.; Musco, L.; Gambi, C.; Saggiomo, V.; Cannavacciuolo, M.; Lo Martire, M.; Passarelli, A.; Zazo, G.; Danovaro, R. Multiple human pressures in coastal habitats: Variation of meiofaunal assemblages associated with sewage discharge in a post-industrial area. Sci. Total Environ. 2019, 655, 1218–1231. [Google Scholar] [CrossRef]
- Hanson, H.; Brampton, A.; Capobianco, M.; Dette, H.H.; Hamm, L.; Laustrup, C.; Lechuga, A.; Spanhoff, R. Beach nourishment projects, practices, and objectives—A European overview. Coast. Eng. 2002, 47, 81–111. [Google Scholar] [CrossRef]
- Airoldi, L. The Effects of Sedimentation on Rocky Coast Assemblages. In Oceanography and Marine Biology, An Annual Review; CRC Press: Boca Raton, FL, USA, 2003; Volume 41, pp. 169–171. [Google Scholar]
- Costello, M.J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A census of marine biodiversity knowledge, resources, and future challenges. PLoS ONE 2010, 5, e12110. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; NyKjaer, R.; Rosenberg, A.A. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: Assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, S.; Katsanevakis, S.; Micheli, F.; Sala, E.; Rilov, G.; Sarà, G.; Malak, D.A.; Abdulla, A.; Gerovasileiou, V.; Gissi, E.; et al. The status of coastal benthic ecosystems in the Mediterranean Sea: Evidence from ecological indicators. Front. Mar. Sci. 2020, 7, 475. [Google Scholar] [CrossRef]
- Dorgham, M.M.; Hamdy, R.; El Rashidy, H.H.; Atta, M.M.; Musco, L. Distribution patterns of shallow water polychaetes (Annelida) along the Alexandria coast, Egypt (eastern Mediterranean). Mediterr. Mar. Sci. 2014, 15, 635–649. [Google Scholar] [CrossRef] [Green Version]
- Pamungkas, J.; Glasby, C.J.; Read, G.B.; Wilson, S.P.; Costello, M.J. Progress and perspectives in the discovery of polychaete worms (Annelida) of the world. Helgol. Mar. Res. 2019, 73, 4. [Google Scholar] [CrossRef]
- Grassle, J.F.; Maciolek, N.J. Deep-sea species richness: Regional and local diversity estimates from quantitative bottom samples. Am. Nat. 1992, 139, 313–341. [Google Scholar] [CrossRef]
- Ward, T.J.; Hutchings, P.A. Effects of trace metals on infaunal species composition in polluted intertidal and subtidal marine sediments near a lead smelter, Spencer Gulf, South Australia. Mar. Ecol. Prog. Ser. 1996, 135, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Arvanitidis, C.; Bellan, G.; Drakopoulos, P.; Valavanis, V.; Dounas, C.; Koukouras, A.; Eleftheriou, A. Seascape biodiversity patterns along the Mediterranean and the Black Sea: Lessons from the biogeography of benthic polychaetes. Mar. Ecol. Prog. Ser. 2002, 244, 139–152. [Google Scholar] [CrossRef]
- Bellan-Santini, D.; Ruffo, S. Biogeography of benthic marine amphipods in Mediterranean Sea. Biogeogr.–J. Integr. Biogeogr. 2003, 24, 273–292. [Google Scholar] [CrossRef] [Green Version]
- Musco, L. Ecology and diversity of Mediterranean hard-bottom Syllidae (Annelida): A community-level approach. Mar. Ecol. Prog. Ser. 2012, 461, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Olsgard, F.; Somerfield, P.J. Surrogates in marine benthic investigations-which taxonomic unit to target? J. Aquat. Ecosyst. Stress Recovery 2000, 7, 25–42. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M.; Musco, L. Polychaetes as environmental indicators revisited. Mar. Pollut. Bull. 2005, 50, 1153–1162. [Google Scholar] [CrossRef]
- Musco, L.; Mikac, B.; Tataranni, M.; Giangrande, A.; Terlizzi, A. The use of coarser taxonomy in the detection of long-term changes in polychaete assemblages. Mar. Environ. Res. 2011, 71, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Dauvin, J.C.; Andrade, H.; de-la-Ossa-Carretero, J.A.; Del-Pilar-Ruso, Y.; Riera, R. Polychaete/amphipod ratios: An approach to validating simple benthic indicators. Ecol. Indic. 2016, 63, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Franco, J.; Pérez, V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Borja, A.; Franco, J.; Valencia, V.; Bald, J.; Muxika, I.; Belzunce, M.J.; Solaun, O. Implementation of the European water framework directive from the Basque country (northern Spain): A methodological approach. Mar. Pollut. Bull. 2004, 48, 209–218. [Google Scholar] [CrossRef]
- Borja, A.; Miles, A.; Occhipinti-Ambrogi, A.; Berg, T. Current status of macroinvertebrate methods used for assessing the quality of European marine waters: Implementing the Water Framework Directive. Hydrobiologia 2009, 633, 181–196. [Google Scholar] [CrossRef]
- Sarda, R. Polychaete communities related to plant covering in the mediolittoral and infralittoral zones of the Balearic Islands (Western Mediterranean). Mar. Ecol. 1991, 12, 341–360. [Google Scholar] [CrossRef]
- Lopez, E.; Vieitez, J.M. Polychaete assemblages on non-encrusting infralittoral algae from the Chafarinas Islands (SW Mediterranean). Cah. Biol. Mar. 1999, 40, 375–384. [Google Scholar]
- Tena, J.; Capaccioni-Azzati, R.; Torres-Gavila, F.J.; García-Carrascosa, A.M. Polychaetes associated with different facies of the photophilic algal community in the Chafarinas archipelago (SW Mediterranean). Bull. Mar. Sci. 2000, 67, 55–72. [Google Scholar]
- Fraschetti, S.; Giangrande, A.; Terlizzi, A.; Miglietta, M.; Della Tommasa, L.; Boero, F. Spatio-temporal variation of hydroids and polychaetes associated with Cystoseira amentacea (Fucales: Phaeophyceae). Mar. Biol. 2002, 140, 949–957. [Google Scholar] [CrossRef]
- Giangrande, A.; Delos, A.L.; Fraschetti, S.; Musco, L.; Licciano, M.; Terlizzi, A. Polychaete assemblages along a rocky shore on the South Adriatic coast (Mediterranean Sea): Patterns of spatial distribution. Mar. Biol. 2003, 143, 1109–1116. [Google Scholar] [CrossRef]
- Cinar, M.E.; Gonlugur-Demirci, G. Polychaete assemblages on shallow-water benthic habitats along the Sinop Peninsula (Black Sea, Turkey). Cah. Biol. Mar. 2005, 46, 253. [Google Scholar]
- Serrano, A.; Preciado, I. Environmental factors structuring polychaete communities in shallow rocky habitats: Role of physical stress versus habitat complexity. Helgol. Mar. Res. 2007, 61, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Musco, L.; Terlizzi, A.; Licciano, M.; Giangrande, A. Taxonomic structure and the effectiveness of surrogates in environmental monitoring: A lesson from polychaetes. Mar. Ecol. Prog. Ser. 2009, 383, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Gambi, M.C.; Musco, L.; Giangrande, A.; Badalamenti, F.; Micheli, F.; Kroeker, K.J. Distribution and functional traits of polychaetes in a CO2 vent system: Winners and losers among closely related species. Mar. Ecol. Prog. Ser. 2016, 550, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Mikac, B.; Licciano, M.; Jaklin, A.; Iveša, L.; Giangrande, A.; Musco, L. Diversity and distribution patterns of hard bottom polychaete assemblages in the North Adriatic Sea (Mediterranean). Diversity 2020, 12, 408. [Google Scholar] [CrossRef]
- Bellan, G. Relationship of pollution to rocky substratum polychaetes on the French Mediterranean coast. Mar. Pollut. Bull. 1980, 11, 318–321. [Google Scholar] [CrossRef]
- Giangrande, A.; Delos, A.L.; Musco, L.; Licciano, M.; Pierri, C. Polychaete assemblages of rocky shore along the South Adriatic coast (Mediterranean Sea). Cah. Biol. Mar. 2004, 45, 85–96. [Google Scholar]
- Abbiati, M.; Bianchi, C.N.; Castelli, A. Polychaete vertical zonation along a littoral cliff in the western Mediterranean. Mar. Ecol. 1987, 8, 33–48. [Google Scholar] [CrossRef]
- Giangrande, A. Polychaete zonation and its relation to algal distribution down a vertical cliff in the western Mediterranean (Italy): A structural analysis. J. Exp. Mar. Biol. Ecol. 1988, 120, 263–276. [Google Scholar] [CrossRef]
- Abd-Elnaby, F.A.; San Martín, G. Eusyllinae, Anoplosyllinae, and Exogoninae (Polychaeta: Syllidae) for the Mediterranean Coasts of Egypt, together the description of one new species. Life Sci. J. 2010, 4, 132–139. [Google Scholar]
- Abd-Elnaby, F.A.; Martin, G.S. Syllinae (Syllidae: Polychaeta) from the Mediterranean coast of Egypt with the description of two new species. Mediterr. Mar. Sci. 2011, 12, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Dorgham, M.M.; Hamdy, R.; El-Rashidy, H.H.; Atta, M.M. First records of polychaetes new to Egyptian Mediterranean waters. Oceanologia 2013, 55, 235–267. [Google Scholar] [CrossRef] [Green Version]
- Hamdy, E.R.; Dorgham, M.M. Macrofauna associated with a recently described bryozoan species in the Eastern Harbour of Alexandria, Egypt. Mediterr. Mar. Sci. 2019, 20, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Bulleri, F.; Chapman, M.G. The introduction of coastal infrastructure as a driver of change in marine environments. J. Appl. Ecol. 2010, 47, 26–35. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; p. 310. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. Armstrong. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, R.K. Permanova+ for Primer: Guide to Software and Statistical Methods; Primer-E Ltd.: Auckland, New Zealand, 2008. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Antoniadou, C.; Chintiroglou, C. Biodiversity of zoobenthic hard-substrate sublittoral communities in the Eastern Mediterranean (North Aegean Sea). Estuar. Coast. Shelf Sci. 2005, 62, 637–653. [Google Scholar] [CrossRef]
- Jones, E.; Thornber, C.S. Effects of habitat-modifying invasive macroalgae on epiphytic algal communities. Mar. Ecol. Prog. Ser. 2010, 400, 87–100. [Google Scholar] [CrossRef]
- Cacabelos, E.; Olabarria, C.; Incera, M.; Troncoso, J.S. Effects of habitat structure and tidal height on epifaunal assemblages associated with macroalgae. Estuar. Coast. Shelf Sci. 2010, 89, 43–52. [Google Scholar] [CrossRef]
- Musco, L.; Franzitta, G.; Colletti, A.; Savinelli, B.; Lo Martire, M.; Corinaldesi, C. Feasibility of the Sabellarid Reef Habitat Restoration. Front. Mar. Sci. 2022, 9, 349. [Google Scholar] [CrossRef]
- Terlizzi, A.; Fraschetti, S.; Guidetti, P.; Boero, F. The effects of sewage discharge on shallow hard substrate sessile assemblages. Mar. Pollut. Bull. 2002, 44, 544–550. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Cinelli, F.; Benedetti-Cecchi, L. Patterns of spatial variability in epiphytes of Posidonia oceanica: Differences between a disturbed and two reference locations. Aquat. Bot. 2004, 79, 345–356. [Google Scholar] [CrossRef]
- Borghese, J.; Musco, L.; Arduini, D.; Tamburello, L.; Del Pasqua, M.; Giangrande, A. A Comparative Approach to Detect Macrobenthic Response to the Conversion of an Inshore Mariculture Plant into an IMTA System in the Mar Grande of Taranto (Mediterranean Sea, Italy). Water 2023, 15, 68. [Google Scholar] [CrossRef]
- Borja, A.; Muxika, I. Guidelines for the use of AMBI (AZTI’s Marine Biotic Index) in the assessment of the benthic ecological quality. Mar. Pollut. Bull. 2005, 7, 787–789. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| EM | GL | ST | EH | MX | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Min | Max | Min | Max | Min | Max | Min | Max | |
| T | 18.30 | 29.7 | 14.80 | 29.90 | 14.90 | 29.20 | 14.60 | 28.80 | 15.40 | 29.50 |
| pH | 7.85 | 8.00 | 7.76 | 8.10 | 7.63 | 8.30 | 7.53 | 8.20 | 7.30 | 8.26 |
| S | 37.64 | 39.91 | 36.46 | 39.82 | 36.49 | 39.75 | 36.50 | 38.36 | 27.44 | 36.04 |
| DO | 0.41 | 8.47 | 7.98 | 15.30 | 6.19 | 13.68 | 3.58 | 19.28 | 5.21 | 7.49 |
| BOD | 0.41 | 1.63 | 0.32 | 2.28 | 0.57 | 11.40 | 0.32 | 15.45 | 4.07 | 6.76 |
| OM | 0.40 | 1.74 | 0.20 | 3.13 | 0.40 | 1.25 | 2.66 | 5.00 | 0.60 | 1.81 |
| Av | SD | Av | SD | Av | SD | Av | SD | Av | SD | |
| T | 22.33 | ±5.22 | 22.52 | ±5.36 | 22.67 | ±5.04 | 22.50 | 5.43 | 22.15 | 5.17 |
| pH | 7.90 | ±0.07 | 7.87 | ±0.12 | 7.85 | ±0.23 | 7.80 | 0.21 | 7.78 | 0.38 |
| S | 38.19 | ±0.99 | 37.47 | ±1.19 | 37.95 | ±1.26 | 37.68 | 0.92 | 31.26 | 3.48 |
| DO | 5.82 | ±2.92 | 9.47 | ±2.70 | 8.98 | ±2.35 | 8.56 | 5.15 | 6.76 | 0.95 |
| BOD | 1.09 | ±0.54 | 1.18 | ±0.79 | 2.79 | ±3.99 | 3.24 | 5.62 | 4.44 | 1.75 |
| OM | 0.88 | ±0.54 | 1.02 | ±1.02 | 0.78 | ±0.26 | 3.31 | 0.81 | 1.09 | 0.47 |
| EM | GL | ST | EH | MX | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Av * | SE * | Av | SE | Av | SE | Av | SE | Av | SE | |
| Cladophora sp. | 0.39 | ±0.66 | 0.06 | ±0.14 | 0.50 | ±0.77 | ||||
| Ulva sp. | 10.50 | ±11.59 | 3.39 | ±5.93 | 25.22 | ±22.06 | 13.89 | ±4.57 | 68.67 | ±20.11 |
| Enteromorpha sp. | 2.78 | ±6.80 | ||||||||
| Petalonia sp. | 0.33 | ±0.69 | 0.06 | ±0.14 | 5.83 | ±8.18 | ||||
| Colpomenia sp. | 0.06 | ±0.14 | 0.61 | ±0.74 | ||||||
| Amphiroa sp. | 2.83 | ±2.51 | 0.11 | ±0.27 | 8.17 | ±10.78 | 2.33 | ±3.72 | 0.11 | ±0.19 |
| Corallina sp. | 31.44 | ±21.60 | 77.06 | ±14.53 | 38.72 | ±17.92 | 28.89 | ±22.57 | 9.83 | ±16.42 |
| Jania sp. | 0.78 | ±1.37 | 8.78 | ±6.59 | 0.11 | ±0.19 | 5.56 | ±9.54 | 0.72 | ±0.99 |
| Griffithsia sp. | 0.17 | ±0.30 | ||||||||
| Pyropia sp. | 0.39 | ±0.72 | ||||||||
| Pterocladia sp. | 5.83 | ±12.22 | 8.06 | ±13.62 | ||||||
| Brachidontes pharaonis | 23.06 | ±21.33 | 6.61 | ±8.78 | 13.33 | ±17.71 | 2.22 | ±3.17 | ||
| Other bivalves | 27.44 | ±21.12 | 4.83 | ±4.91 | 20.44 | ±17.23 | 34.33 | ±18.01 | 1.94 | ±2.21 |
| Balanus sp. | 1.67 | ±2.21 | 1.11 | ±2.11 | ||||||
| Bryozoa | 0.83 | ±1.03 | ||||||||
| Species | EM | GL | ST | EH | MX | Tot | Tot |
|---|---|---|---|---|---|---|---|
| ind/m2 | ind/m2 | ind/m2 | ind/m2 | ind/m2 | ind/m2 | % | |
| Syllis columbretensis (Campoy, 1982) | 56,390 | 31,920 | 7710 | 23,750 | 14,790 | 134,560 | 17.468 |
| Kirkegaardia heterochaeta (Laubier, 1961) | 41,600 | 780 | 57,490 | 700 | 30 | 100,600 | 13.06 |
| Syllis armillaris (O.F. Müller, 1776) | 7760 | 2220 | 21,740 | 64,610 | 3600 | 99,930 | 12.973 |
| Syllis prolifera (Krohn, 1852) | 1400 | 6350 | 13,270 | 32,850 | 11,190 | 65,060 | 8.4458 |
| Pseudonereis anomala (Gravier, 1899) | 36,980 | 4720 | 11,890 | 5470 | 3380 | 62,440 | 8.1057 |
| Naineris laevigata (Grube, 1855) | 1480 | 4040 | 19,150 | 5170 | 0 | 29,840 | 3.8737 |
| Syllis pulvinata (Langerhans, 1881) | 27,640 | 100 | 1110 | 130 | 0 | 28,980 | 3.7621 |
| Cirriformia filigera (Delle Chiaje, 1828) | 840 | 13,090 | 10,540 | 3050 | 720 | 28,240 | 3.666 |
| Syllis variegata (Grube, 1860) | 5880 | 7320 | 6560 | 7070 | 10 | 26,840 | 3.4843 |
| Amphicorina bichaeta (Capa & López, 2004) | 100 | 230 | 9050 | 14,500 | 10 | 23,890 | 3.1013 |
| Hydroides elegans (Haswell, 1883) | 560 | 620 | 250 | 9040 | 4190 | 14,660 | 1.9031 |
| Platynereis insolita (Gravier, 1899) | 7510 | 5700 | 1100 | 230 | 80 | 14,620 | 1.8979 |
| Eurythoe complanata (Pallas, 1766) | 13,430 | 10 | 0 | 30 | 0 | 13,470 | 1.7486 |
| Dorvillea similis (Crossland, 1924) | 300 | 2510 | 9240 | 10 | 0 | 12,060 | 1.5656 |
| Salvatoria clavata (Claparède, 1863) | 270 | 1560 | 1880 | 4890 | 1190 | 9790 | 1.2709 |
| Branchiomaldane vincenti (Langerhans, 1881) | 40 | 2820 | 2640 | 3420 | 0 | 8920 | 1.158 |
| Polyophthalmus pictus (Dujardin, 1839) | 1100 | 1330 | 3860 | 2350 | 60 | 8700 | 1.1294 |
| Dipolydora coeca (Örsted, 1843) | 790 | 790 | 190 | 5340 | 140 | 7250 | 0.9412 |
| Scyphoproctus cf. armatus (Fauvel, 1929) | 1790 | 2150 | 950 | 2040 | 10 | 6940 | 0.9009 |
| Spirobranchus triqueter (Linnaeus, 1758) | 1610 | 70 | 330 | 4020 | 400 | 6430 | 0.8347 |
| Linopherus canariensis (Langerhans, 1881) | 3700 | 120 | 1390 | 1110 | 0 | 6320 | 0.8204 |
| Oxydromus pallidus (Claparède, 1864) | 3360 | 300 | 850 | 1520 | 10 | 6040 | 0.7841 |
| Perkinsyllis anophthalma (Capaccioni & San Martín, 1990) | 80 | 250 | 0 | 4320 | 0 | 4650 | 0.6036 |
| Chrysopetalum debile (Grube, 1855) | 260 | 2090 | 1320 | 480 | 0 | 4150 | 0.5387 |
| Caulleriella cf. cristata (Blake, 1996) | 560 | 640 | 600 | 1800 | 0 | 3600 | 0.4673 |
| Branchiosyllis exilis (Gravier, 1900) | 560 | 2090 | 140 | 530 | 0 | 3320 | 0.431 |
| Loimia medusa (Savigny, 1822) | 210 | 0 | 930 | 2020 | 20 | 3180 | 0.4128 |
| Dipolydora armata (Langerhans, 1880) | 0 | 280 | 2300 | 0 | 10 | 2590 | 0.3362 |
| Alitta succinea (Leuckart, 1847) | 580 | 250 | 270 | 250 | 890 | 2240 | 0.2908 |
| Odontosyllis fulgurans (Audouin & Milne Edwards, 1833) | 290 | 1120 | 430 | 340 | 0 | 2180 | 0.283 |
| Caulleriella alata (Southern, 1914) | 20 | 0 | 100 | 1840 | 0 | 1960 | 0.2544 |
| Syllis gracilis (Grube, 1840) | 50 | 220 | 960 | 20 | 470 | 1720 | 0.2233 |
| Brania arminii (Langerhans, 1881) | 60 | 30 | 1400 | 120 | 30 | 1640 | 0.2129 |
| Lumbrineris perkinsi (Carrera-Parra, 2001) | 510 | 10 | 360 | 730 | 0 | 1610 | 0.209 |
| Dodecaceria concharum (Örsted, 1843) | 140 | 910 | 230 | 100 | 70 | 1450 | 0.1882 |
| Opisthosyllis brunnea (Langerhans, 1879) | 550 | 550 | 290 | 0 | 0 | 1390 | 0.1804 |
| Myrianida prolifera (O.F. Müller, 1788) | 10 | 10 | 30 | 1310 | 10 | 1370 | 0.1778 |
| Cirriformia tentaculata (Montagu, 1808) | 0 | 600 | 30 | 720 | 0 | 1350 | 0.1753 |
| Hydroides operculata (Treadwell, 1929) | 260 | 0 | 240 | 780 | 40 | 1320 | 0.1714 |
| Capitella capitata (Fabricius, 1780) | 30 | 20 | 210 | 990 | 10 | 1260 | 0.1636 |
| Exogone africana (Hartmann-Schröder, 1974) | 20 | 430 | 230 | 390 | 10 | 1080 | 0.1402 |
| Lepidonotus clava (Montagu, 1808) | 30 | 30 | 110 | 830 | 0 | 1000 | 0.1298 |
| Trypanosyllis sanmartini (Çinar, 2007) | 0 | 10 | 10 | 920 | 0 | 940 | 0.122 |
| Thelepus setosus (Quatrefages, 1866) | 10 | 440 | 300 | 170 | 0 | 920 | 0.1194 |
| Dipolydora giardi (Mesnil, 1893) | 140 | 120 | 10 | 560 | 0 | 830 | 0.1077 |
| Lysidice unicornis (Grube, 1840) | 310 | 60 | 340 | 60 | 10 | 780 | 0.1013 |
| Boccardia polybranchia (Haswell, 1885) | 0 | 720 | 60 | 0 | 0 | 780 | 0.1013 |
| Nereiphylla rubiginosa (de Saint-Joseph, 1888) | 70 | 50 | 70 | 400 | 0 | 590 | 0.0766 |
| Schistomeringos rudolphi (Delle Chiaje, 1828) | 10 | 0 | 0 | 550 | 0 | 560 | 0.0727 |
| Ctenodrilus serratus (Schmidt, 1857) | 0 | 0 | 460 | 0 | 90 | 550 | 0.0714 |
| Brania pusilla (Dujardin, 1851) | 10 | 0 | 50 | 440 | 0 | 500 | 0.0649 |
| Syllidia armata (Quatrefages, 1866) | 210 | 0 | 30 | 200 | 0 | 440 | 0.0571 |
| Pseudopolydora antennata (Claparède, 1869) | 110 | 90 | 70 | 170 | 0 | 440 | 0.0571 |
| Myrianida convoluta (Cognetti, 1953) | 0 | 10 | 40 | 290 | 30 | 370 | 0.048 |
| Scoletoma laurentiana (Grube, 1863) | 90 | 10 | 50 | 210 | 0 | 360 | 0.0467 |
| Eumida sanguinea (Örsted, 1843) | 60 | 50 | 120 | 130 | 0 | 360 | 0.0467 |
| Spirorbis sp. | 10 | 0 | 0 | 310 | 0 | 320 | 0.0415 |
| Haplosyllis spongicola (Grube, 1855) | 10 | 0 | 230 | 70 | 0 | 310 | 0.0402 |
| Syllides convolutes (Webster & Benedict, 1884) | 0 | 0 | 60 | 170 | 10 | 240 | 0.0312 |
| Ophryotrocha cf. adherens (Paavo, Bailey-Brock & Akesson, 2000) | 0 | 30 | 0 | 180 | 0 | 210 | 0.0273 |
| Serpula hartmanae (Reish, 1968) | 10 | 0 | 70 | 120 | 0 | 200 | 0.026 |
| Nereis splendida (Grube, 1840) | 0 | 0 | 0 | 100 | 80 | 180 | 0.0234 |
| Sabellaria spinulosa (Leuckart, 1849) | 110 | 20 | 30 | 10 | 0 | 170 | 0.0221 |
| Sphaerosyllis hystrix (Claparède, 1863) | 40 | 0 | 120 | 0 | 0 | 160 | 0.0208 |
| Branchiomma bairdi (McIntosh, 1885) | 0 | 30 | 90 | 30 | 0 | 150 | 0.0195 |
| Armandia casuarina (Moreira & Parapar, 2017) | 0 | 20 | 0 | 110 | 0 | 130 | 0.0169 |
| Amphiglena mediterranea (Leydig, 1851) | 0 | 0 | 110 | 10 | 0 | 120 | 0.0156 |
| Marphysa sanguinea (Montagu, 1813) | 10 | 0 | 10 | 90 | 0 | 110 | 0.0143 |
| Mesochaetopterus sagittarius (Claparède, 1870) | 0 | 0 | 90 | 0 | 0 | 90 | 0.0117 |
| Trypanosyllis zebra (Grube, 1860) | 0 | 20 | 50 | 10 | 0 | 80 | 0.0104 |
| Capitella minima (Langerhans, 1880) | 0 | 20 | 50 | 0 | 0 | 70 | 0.0091 |
| Oxydromus agilis (Ehlers, 1864) | 30 | 0 | 0 | 30 | 0 | 60 | 0.0078 |
| Spirobranchus tetraceros (Schmarda, 1861) | 0 | 0 | 0 | 60 | 0 | 60 | 0.0078 |
| Pionosyllis sp. | 10 | 0 | 20 | 30 | 0 | 60 | 0.0078 |
| Neanthes acuminata (Ehlers, 1868) | 0 | 0 | 0 | 50 | 0 | 50 | 0.0065 |
| Arabella iricolor (Montagu, 1804) | 40 | 0 | 10 | 0 | 0 | 50 | 0.0065 |
| Halla parthenopeia (Delle Chiaje, 1828) | 20 | 0 | 10 | 20 | 0 | 50 | 0.0065 |
| Vermiliopsis infundibulum (Philippi, 1844) | 0 | 0 | 40 | 10 | 0 | 50 | 0.0065 |
| Leodice antennata Savigny in (Lamarck, 1818) | 30 | 0 | 10 | 0 | 0 | 40 | 0.0052 |
| Pharyngocirrus goodrichi (Jouin-Toulmond & Gambi, 2007) | 10 | 0 | 10 | 20 | 0 | 40 | 0.0052 |
| Prionospio lighti (Maciolek, 1985) | 0 | 0 | 0 | 40 | 0 | 40 | 0.0052 |
| Pettiboneia urciensis (Campoy & San Martín, 1980) | 0 | 0 | 0 | 30 | 0 | 30 | 0.0039 |
| Leodice miurai (Carrera-Parra & Salazar-Vallejo, 1998) | 20 | 0 | 0 | 0 | 0 | 20 | 0.0026 |
| Perinereis cultrifera (Grube, 1840) | 20 | 0 | 0 | 0 | 0 | 20 | 0.0026 |
| Phyllodoce longifrons (Ben-Eliahu, 1972) | 20 | 0 | 0 | 0 | 0 | 20 | 0.0026 |
| Aphrogenia sp. | 0 | 10 | 0 | 0 | 0 | 10 | 0.0013 |
| Protodorvillea cf. kefersteini (McIntosh, 1869) | 0 | 0 | 10 | 0 | 0 | 10 | 0.0013 |
| Podarkeopsis capensis (Day, 1963) | 0 | 0 | 0 | 10 | 0 | 10 | 0.0013 |
| Leonnates decipiens (Fauvel, 1929) | 0 | 10 | 0 | 0 | 0 | 10 | 0.0013 |
| Platynereis dumerilii (Audouin & Milne Edwards, 1833) | 10 | 0 | 0 | 0 | 0 | 10 | 0.0013 |
| Subadyte cf. pellucida (Ehlers, 1864) | 0 | 0 | 10 | 0 | 0 | 10 | 0.0013 |
| Claudrilus helgolandicus (von Nordheim, 1983) | 10 | 0 | 0 | 0 | 0 | 10 | 0.0013 |
| Polydora cornuta (Bosc, 1802) | 0 | 0 | 0 | 10 | 0 | 10 | 0.0013 |
| Pseudopolydora paucibranchiata (Okuda, 1937) | 0 | 0 | 10 | 0 | 0 | 10 | 0.0013 |
| Spio blakei (Maciolek, 1990) | 0 | 0 | 0 | 10 | 0 | 10 | 0.0013 |
| Salvatoria limbata (Claparède, 1868) | 10 | 0 | 0 | 0 | 0 | 10 | 0.0013 |
| Syllis sp. | 0 | 0 | 0 | 10 | 0 | 10 | 0.0013 |
| Syllis rosea (Langerhans, 1879) | 0 | 0 | 0 | 0 | 10 | 10 | 0.0013 |
| A | Source | df | MS | Pseudo-F | P(perm) | Unique perms |
|---|---|---|---|---|---|---|
| IM | 1 | 514.31 | 0.3036 | 0.9270 | 9956 | |
| TI | 5 | 472.78 | 5.6613 | 0.0037 | 9964 | |
| SI(IM) | 3 | 1883.30 | 22.5510 | 0.0001 | 9955 | |
| IM×TI | 5 | 85.94 | 1.0291 | 0.4407 | 9954 | |
| SI(IM)×TI | 15 | 83.51 | 1.6247 | 0.0906 | 9932 | |
| Res | 60 | 51.40 | ||||
| Total | 89 | |||||
| PERMDISP F1,88 19.539; P(perm) 0.0001 | ||||||
| B | IM | 1 | 135070000 | 0.5047 | 0.7746 | 9943 |
| TI | 5 | 530420000 | 3.9392 | 0.0225 | 9945 | |
| SI(IM) | 3 | 432760000 | 3.2139 | 0.0481 | 9949 | |
| IM×TI | 5 | 101640000 | 0.7549 | 0.6033 | 9947 | |
| SI(IM)×TI | 15 | 134650000 | 1.7747 | 0.0521 | 9907 | |
| Res | 60 | 75875000 | ||||
| Total | 89 | |||||
| PERMDISP F1,88 0.58742; P(perm) 0.6196 | ||||||
| C | IM | 1 | 5.8163 | 0.33693 | 0.9091 | 9949 |
| TI | 5 | 3.3201 | 5.6279 | 0.0038 | 9963 | |
| SI(IM) | 3 | 18.4000 | 31.1900 | 0.0001 | 9953 | |
| IM×TI | 5 | 0.6140 | 1.0407 | 0.4260 | 9943 | |
| SI(IM)×TI | 15 | 0.5899 | 1.1899 | 0.3049 | 9918 | |
| Res | 60 | 0.4958 | ||||
| Total | 89 | |||||
| PERMDISP F1,88 23.886; P(perm) 0.0001 | ||||||
| D | IM | 1 | 0.043 | 0.6513 | 0.6582 | 9951 |
| TI | 5 | 0.015 | 0.6865 | 0.6366 | 9945 | |
| SI(IM) | 3 | 0.084 | 3.7603 | 0.0331 | 9952 | |
| IM×TI | 5 | 0.017 | 0.7655 | 0.5967 | 9963 | |
| SI(IM)×TI | 15 | 0.022 | 2.0388 | 0.028 | 9926 | |
| Res | 60 | 0.011 | ||||
| Total | 89 | |||||
| PERMDISP F1,88 0.55745; P(perm) 0.4741 | ||||||
| Source | df | MS | Pseudo-F | P(perm) | Unique Perms |
|---|---|---|---|---|---|
| IM | 1 | 16,170 | 1.219 | 0.2509 | 9910 |
| TI | 5 | 3480 | 2.0459 | 0.0003 | 9872 |
| SI(IM) | 3 | 12,745 | 7.4927 | 0.0001 | 9930 |
| IM × TI | 5 | 1915.2 | 1.126 | 0.2661 | 9873 |
| SI(IM) × TI | 15 | 1701 | 2.2651 | 0.0001 | 9794 |
| Res | 60 | 750.95 | |||
| Total | 89 | ||||
| PERMDISP F1,88 9.0908; P(perm) 0.0085 | |||||
| August | October | December | February | April | July | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | P(MC*) | t | P(MC) | T | P(MC) | t | P(MC) | t | P(MC) | t | P(MC) | |
| Within level ‘UR’ of factor ‘IM’ | ||||||||||||
| EM, GL | 1.9619 | 0.0488 | 3.2624 | 0.0069 | 2.116 | 0.0295 | 1.4134 | 0.188 | 2.8186 | 0.0075 | 2.3494 | 0.0165 |
| EM, ST | 1.3533 | 0.1852 | 2.9314 | 0.0064 | 2.8485 | 0.0109 | 0.90941 | 0.457 | 2.7358 | 0.0123 | 1.9353 | 0.0537 |
| GL, ST | 1.7026 | 0.0695 | 2.7167 | 0.0093 | 3.1222 | 0.0084 | 1.6249 | 0.1225 | 2.0335 | 0.0309 | 1.9586 | 0.0465 |
| Within level ‘DI’ of factor ‘IM’ | ||||||||||||
| EH, MX | 3.2934 | 0.0058 | 3.5489 | 0.0048 | 1.6908 | 0.0771 | 2.2494 | 0.0217 | 3.4577 | 0.0041 | 2.6893 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdy, R.; Elebiary, N.; Naby, F.A.; Borghese, J.; Dorgham, M.; Hamdan, A.; Musco, L. Hard-Bottom Polychaetes Exposed to Multiple Human Pressure along the Mediterranean Coast of Egypt. Water 2023, 15, 997. https://doi.org/10.3390/w15050997
Hamdy R, Elebiary N, Naby FA, Borghese J, Dorgham M, Hamdan A, Musco L. Hard-Bottom Polychaetes Exposed to Multiple Human Pressure along the Mediterranean Coast of Egypt. Water. 2023; 15(5):997. https://doi.org/10.3390/w15050997
Chicago/Turabian StyleHamdy, Rasha, Noha Elebiary, Faiza Abdel Naby, Jacopo Borghese, Mohamed Dorgham, Amira Hamdan, and Luigi Musco. 2023. "Hard-Bottom Polychaetes Exposed to Multiple Human Pressure along the Mediterranean Coast of Egypt" Water 15, no. 5: 997. https://doi.org/10.3390/w15050997
APA StyleHamdy, R., Elebiary, N., Naby, F. A., Borghese, J., Dorgham, M., Hamdan, A., & Musco, L. (2023). Hard-Bottom Polychaetes Exposed to Multiple Human Pressure along the Mediterranean Coast of Egypt. Water, 15(5), 997. https://doi.org/10.3390/w15050997