
Seawater and Freshwater Circulations through Coastal Forested Wetlands on a Caribbean Island

Abstract
:1. Introduction

2. Materials and Methods
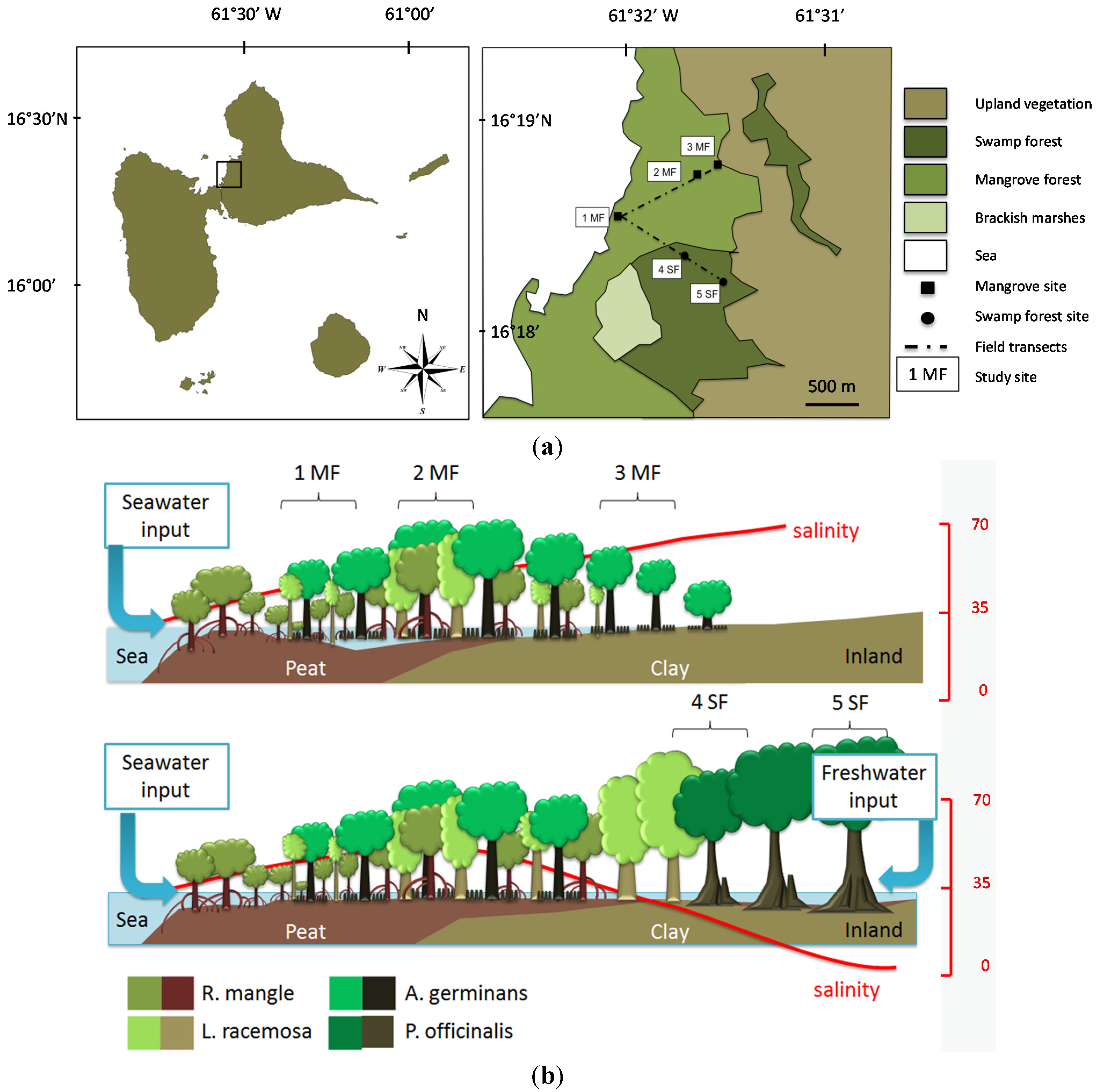
2.1. Study Sites

2.2. Water Level and Soil Salinity
2.3. Isotopic Analysis
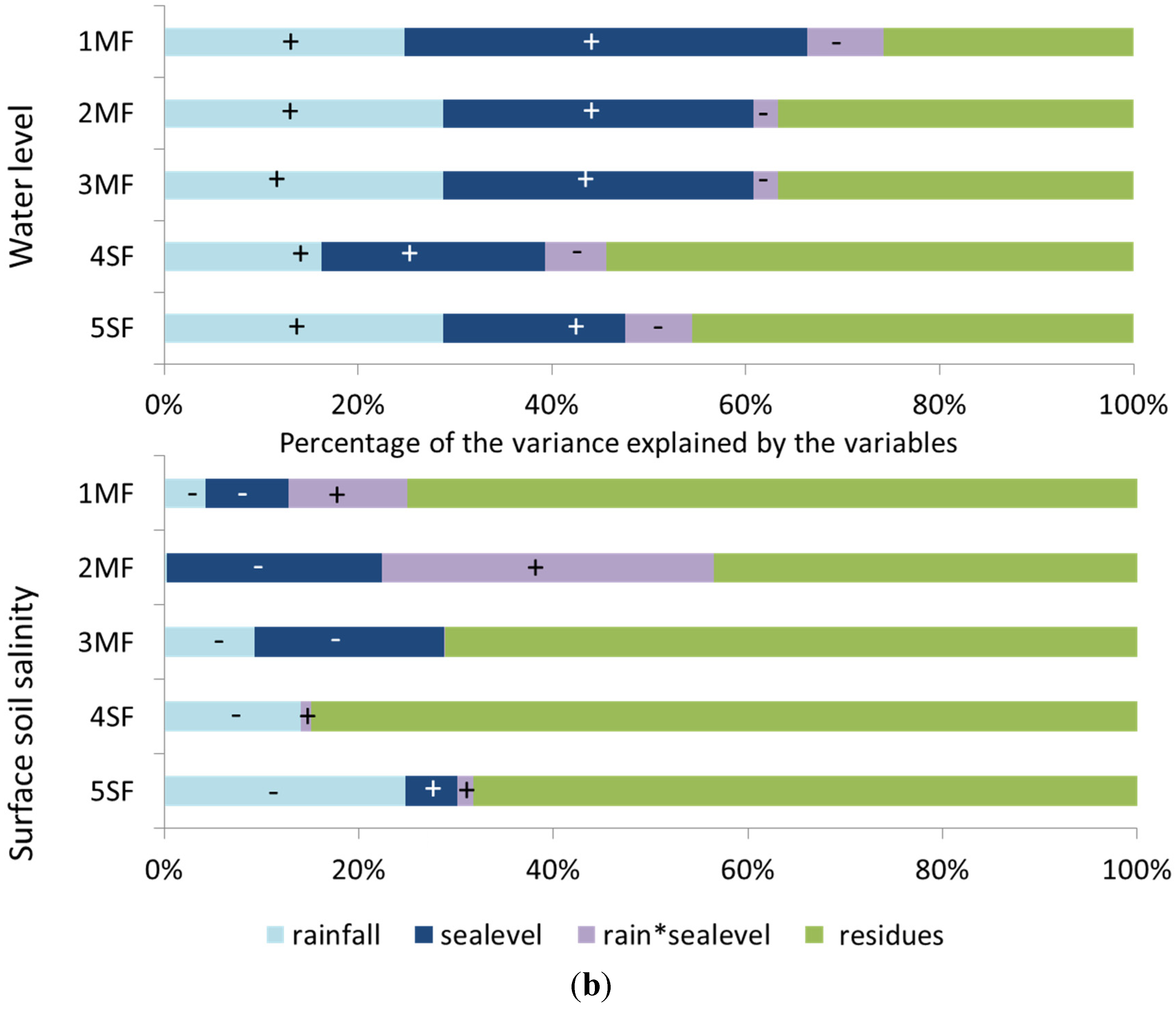
2.4. Statistical Tests
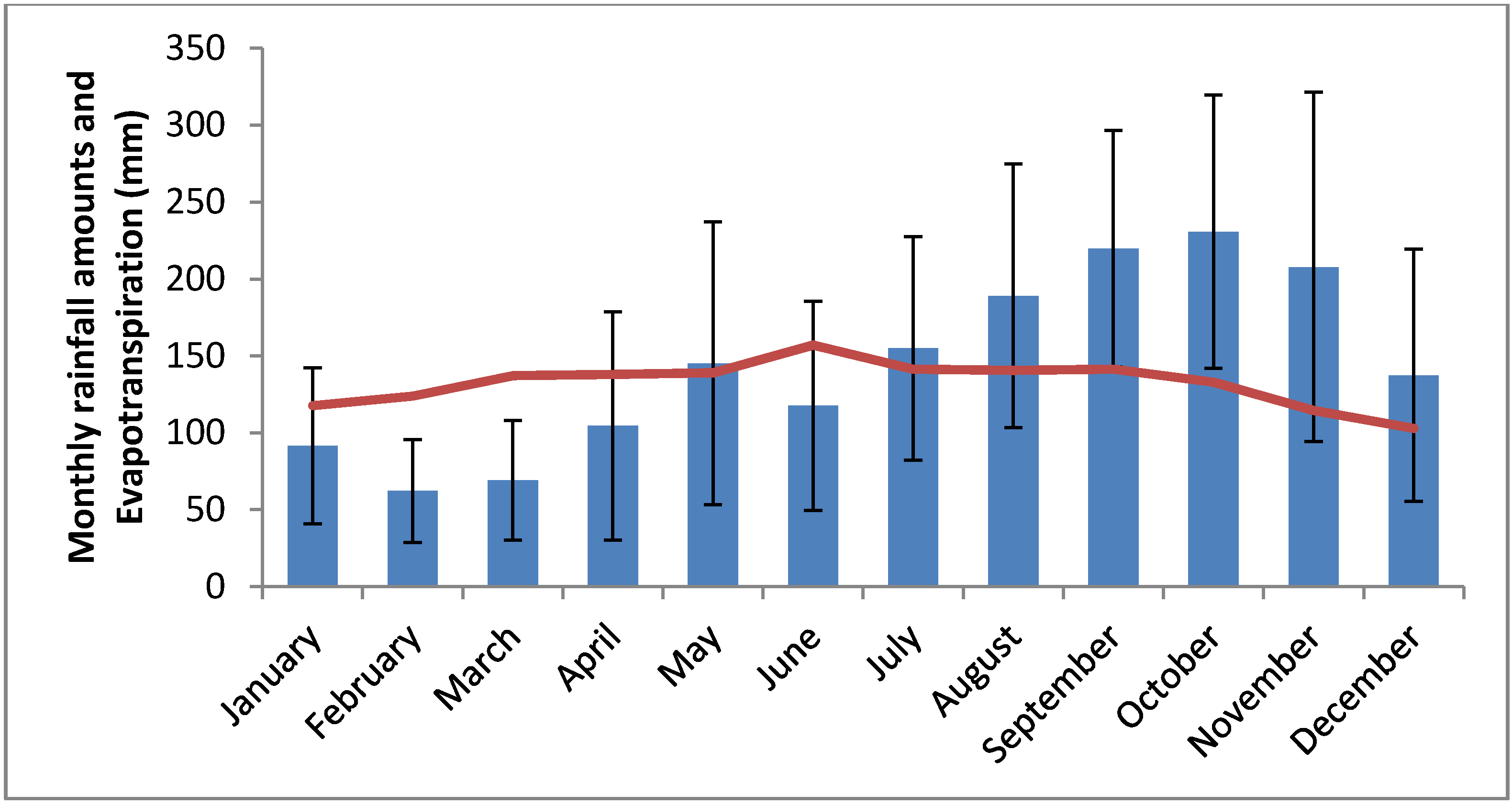
3. Results
3.1. Study Sites Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecosystem | Stations | Distance from the Sea (m) | Relative Elevation (cm) | Water Level (cm) Mean (min ; max)) | Mean Soil Salinity (g·L−1) Mean (min ; max) |
|---|---|---|---|---|---|
| Mangrove | 1MF | 50 | 0.0 | −6.2 (−34 ; 4) | 32.2 (21 ; 53) |
| 2MF | 750 | −6.8 | 3.0 (−39 ; 13) | 49.9 (33 ; 65) | |
| 3MF | 1000 | 1.6 | −11.3 (−47 ; 11) | 72.0 (35 ; 113) | |
| Swamp forest | 4SF | 1050 | −0.9 | 0.3 (−33 ; 11) | 20.8 (12 ; 40) |
| 5SF | 1350 | 2.1 | −4.9 (−26 ; 4) | 6.6 (2.8 ; 12) |
3.2. Hydrological Parameters


3.3. Groundwater Dynamics


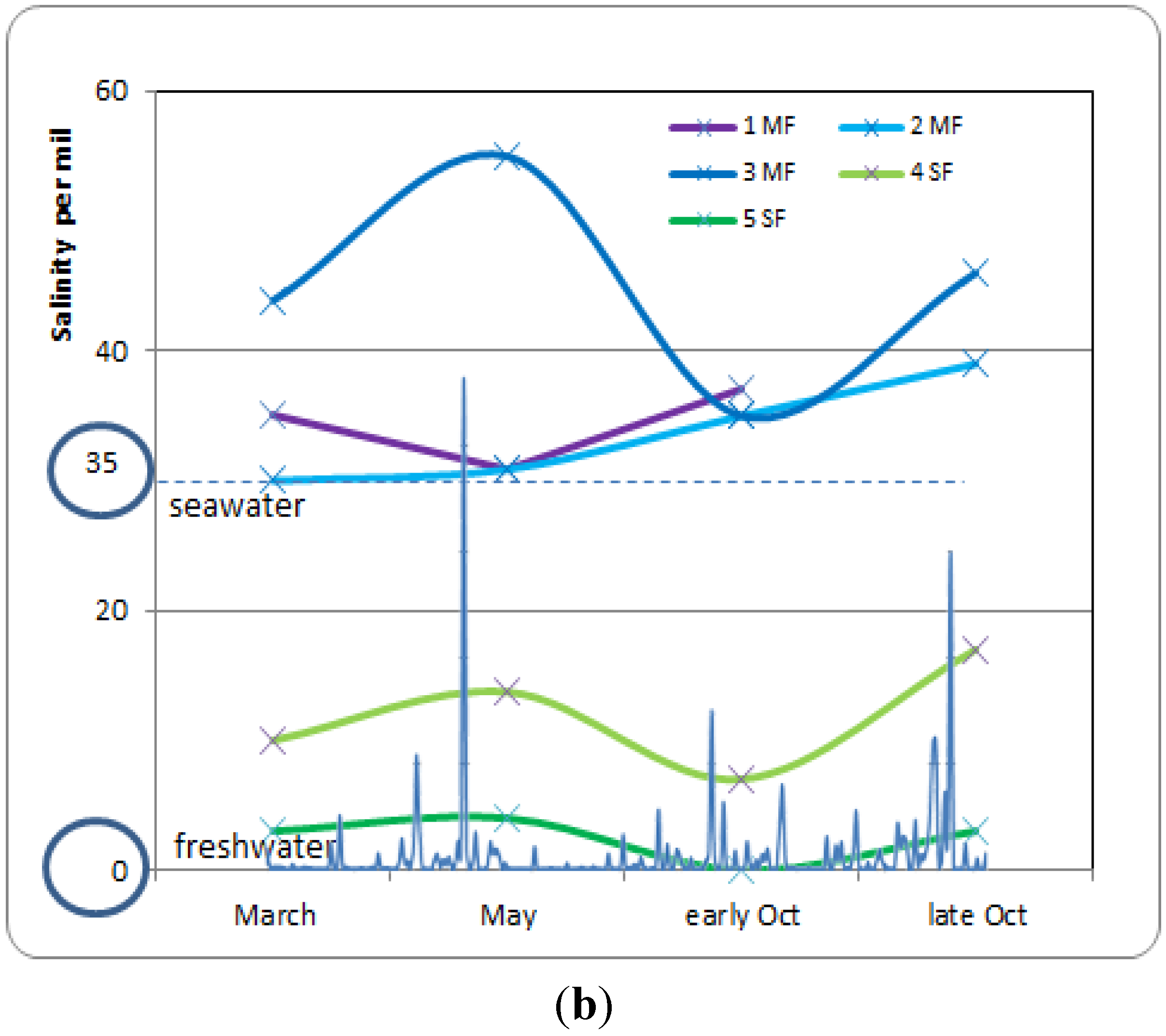
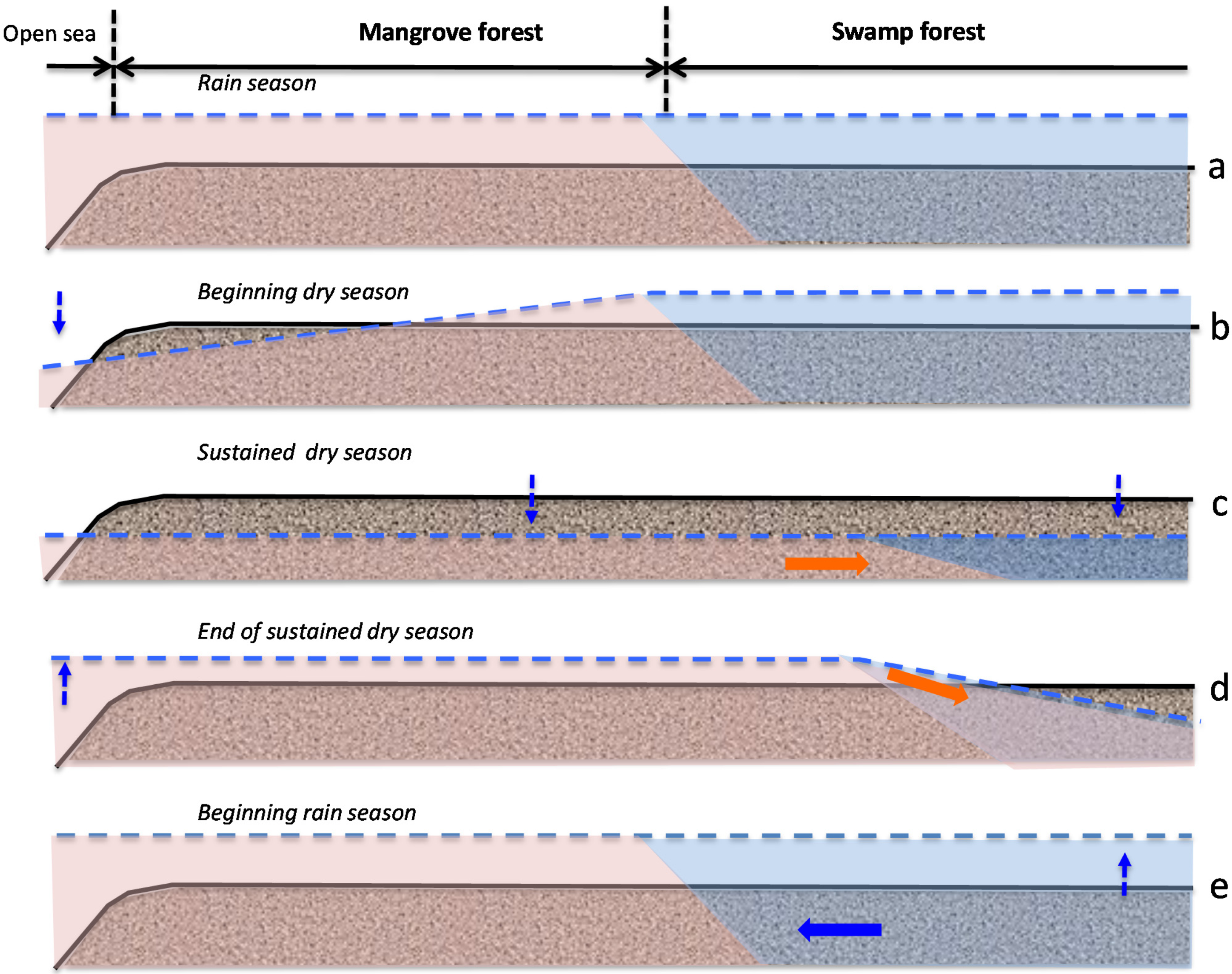
3.4. Soil Salinity and Water-Table Dynamics in the Mangrove and Swamp Forests
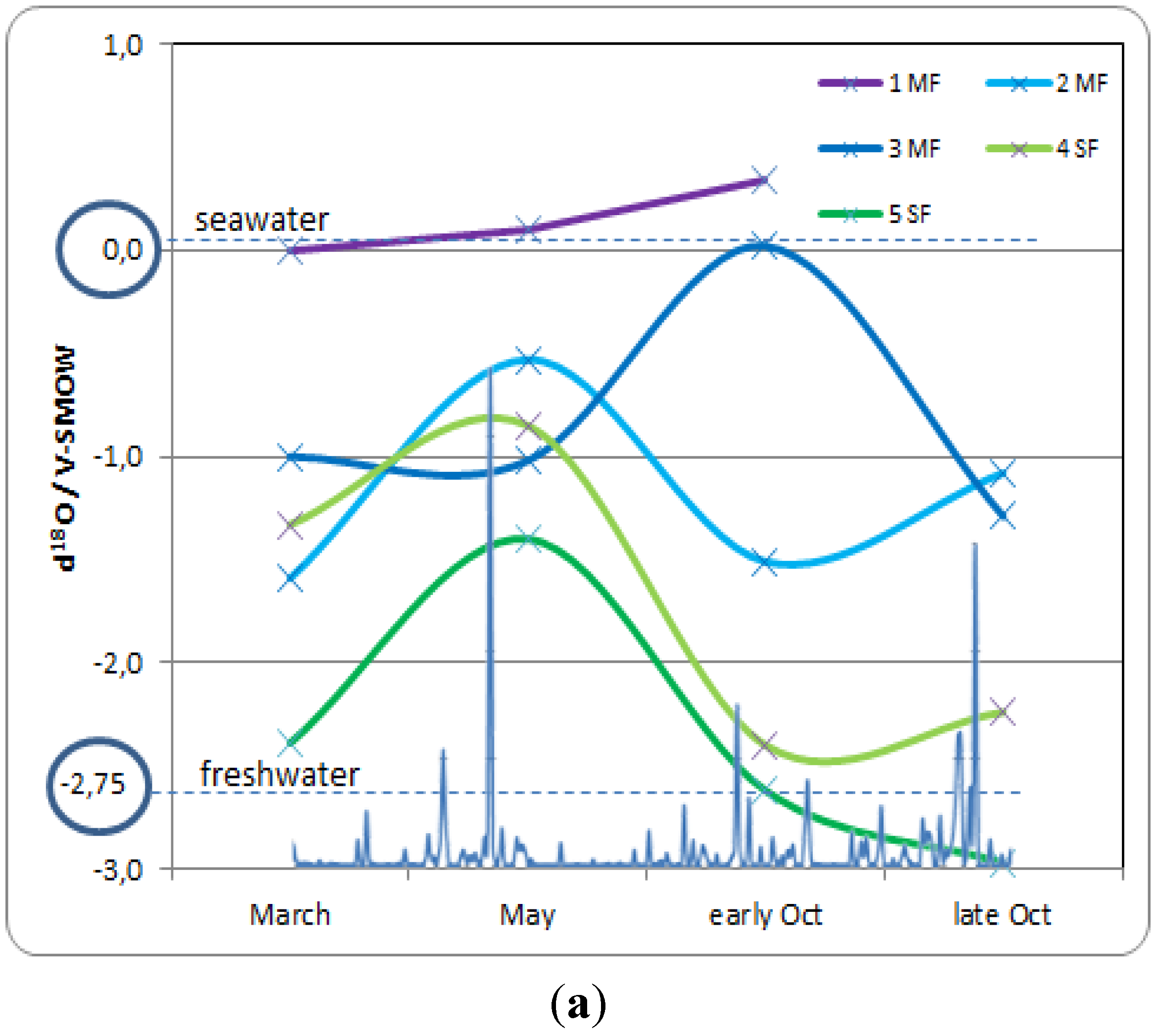
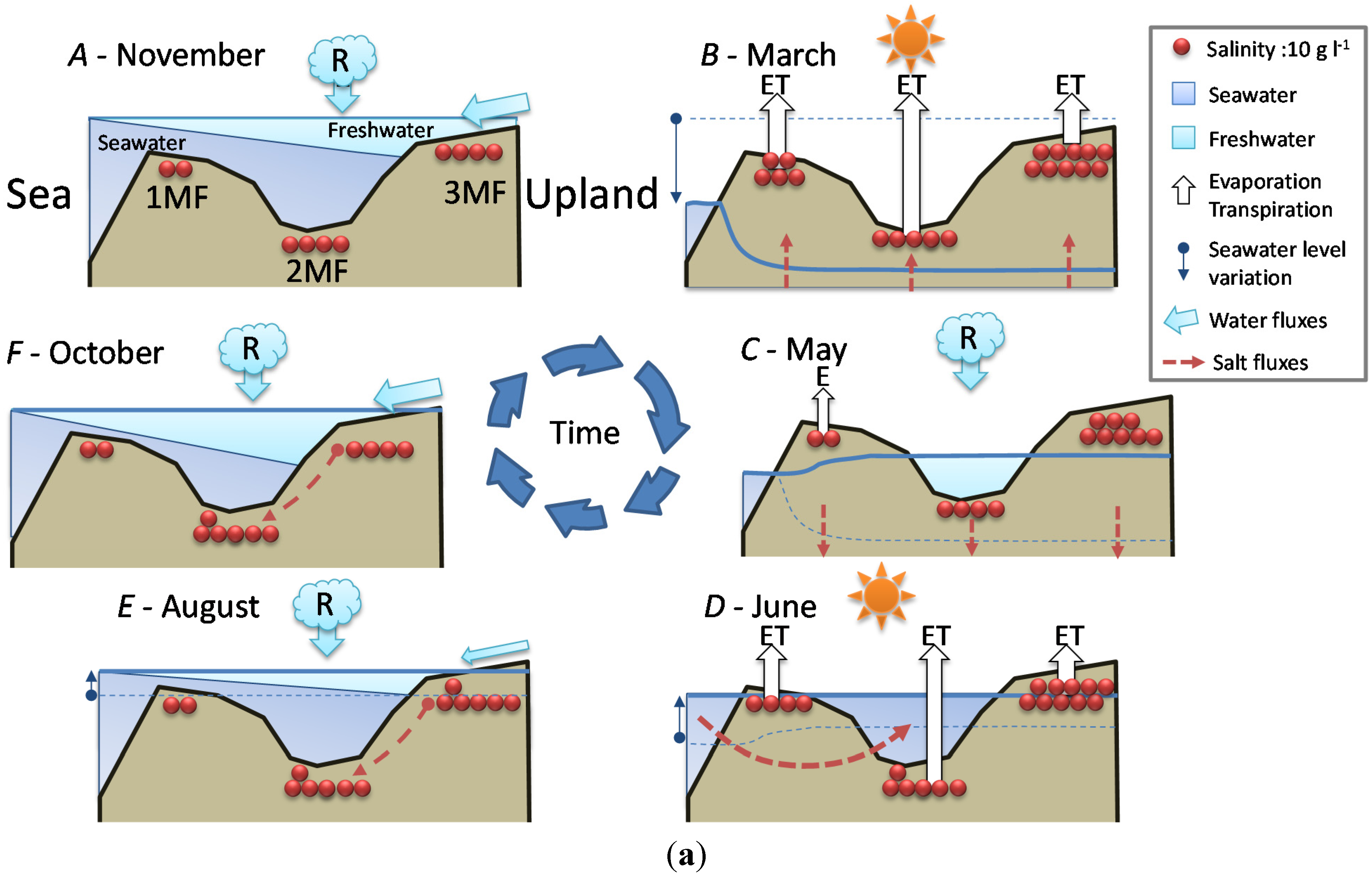
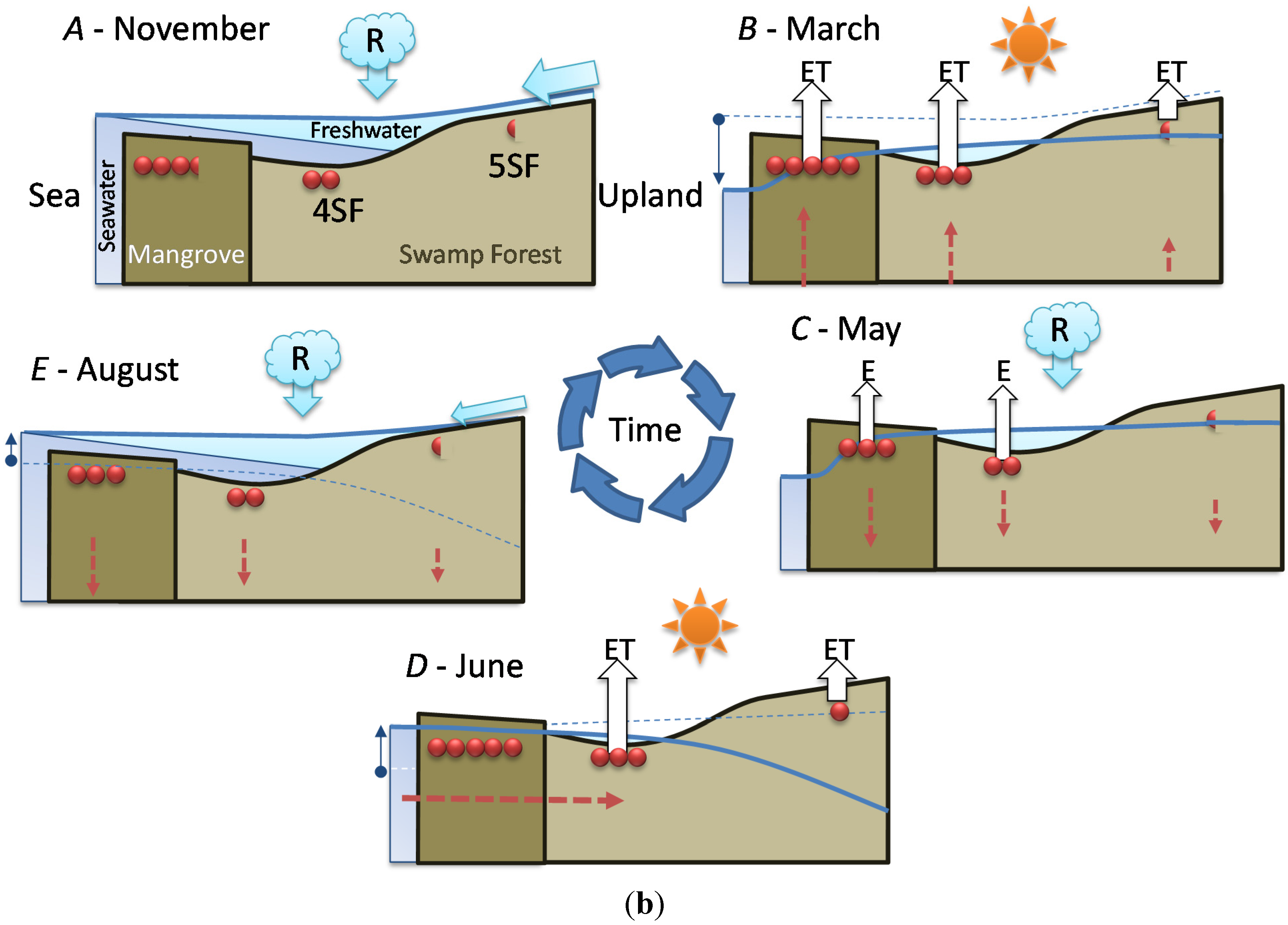
- Stage A: during the rainy season (November 2011, Figure 5a), high mean sea level and heavy rainfall homogenize soil salinity in all mangrove sites with values close to that of seawater (35 g·L−1). High water levels cause seawater to merge landward with freshwater inflow coming from the watershed through the swamp forest. As a result, soil salinity at site 4SF (located nearby the mangrove/swamp forest ecotone) is moderately brackish, whereas it is almost fresh farther inside the swamp forest (site 5SF).
- Stage B: during the dry season (March 2012), sea level and water level drop together during several months due to high atmospheric pressure and gravity forces, respectively. Concomitant evapotranspiration processes cause water level to fall even below sea level all over the mangrove forest and groundwater salinity to rise, up to the seaward edge of the swamp forest. At that time, despite lower rainfall amounts, the upland, fresh groundwater discharge is still sufficient to prevent saltwater intrusion inside the swamp forest (site 5SF).
- Stage C: as the dry season is interrupted by a heavy rainfall episode (April and May 2012), the water level is rising again and salt deposits are re-dissolved. But the sea level still remains low.
- Stage D: by the end of the dry season (June 2012), rainfall lowers again and evapotranspiration increases, causing soil salinity to rise everywhere. As atmospheric pressure lowers, sea level rises, and saltwater penetrates into the mangrove. This inflow of seawater takes the place of the (fresh) water losses by evapotranspiration, and increases salt concentration in the water table. Beyond the tidal area, the water table drops dramatically due to freshwater shortage, allowing in-depth, brackish groundwater intrusion inside the swamp forest.
- Stage E: at the beginning of the next rainy season (August 2012), heavy rains wash out topsoil salt from higher elevation sites (1MF, 3MF) onto lower ones (2MF, sea fringe). Thus, soil salinity remains high at site 2MF, whereas water level rises everywhere, especially inside the swamp forest.
- Stage F: in the middle of a rainy season (November 2011, October 2012), large amounts of freshwater flow through coastal wetlands and dilute mangrove groundwater.


4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
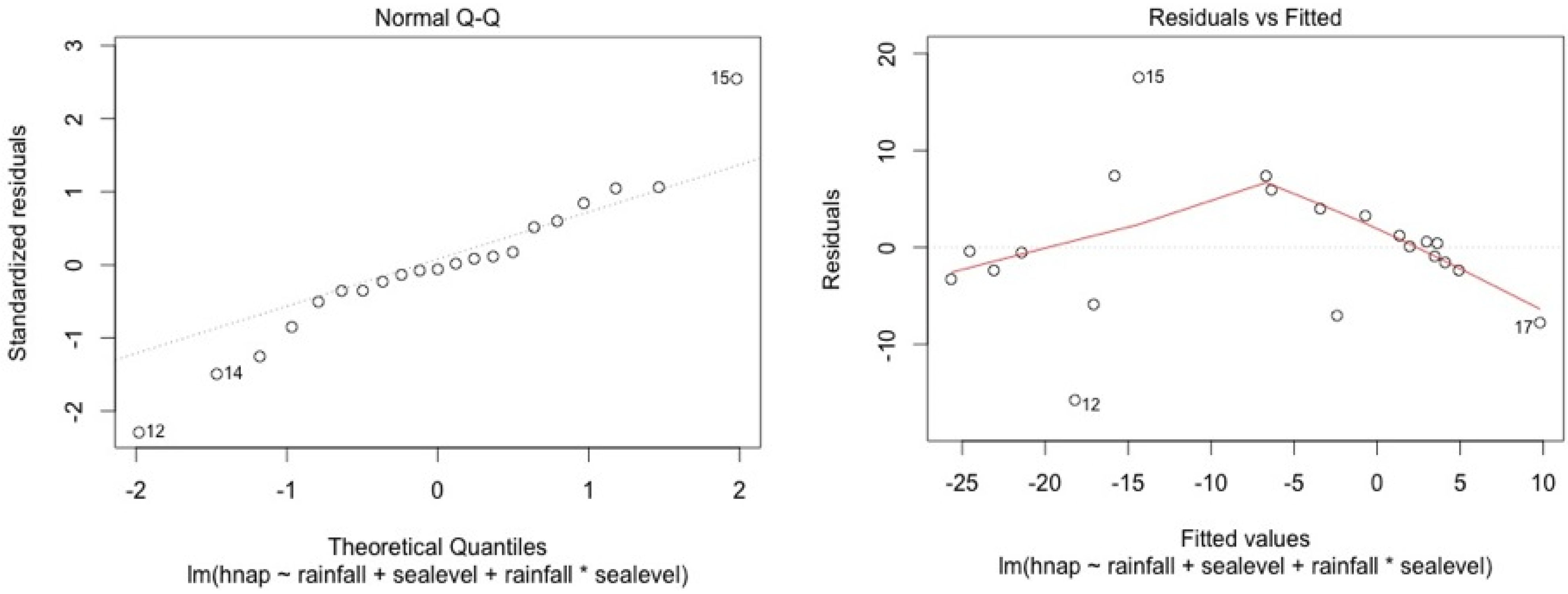
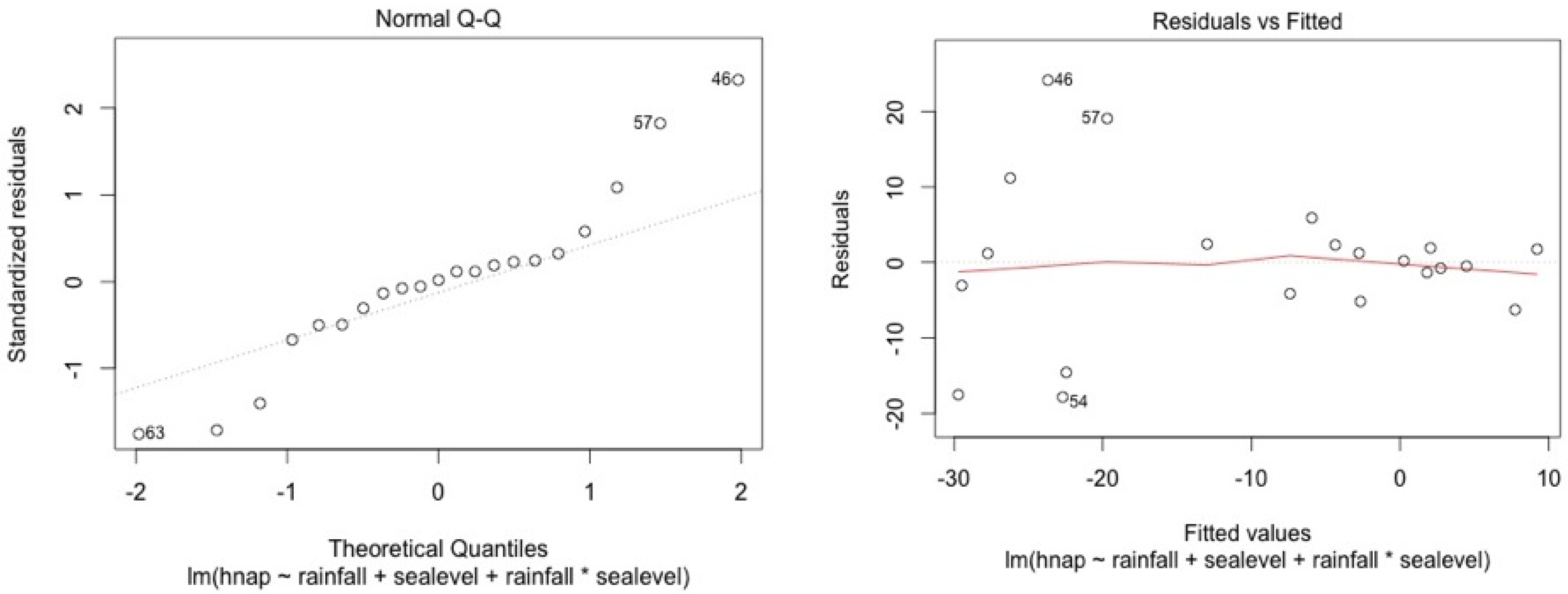
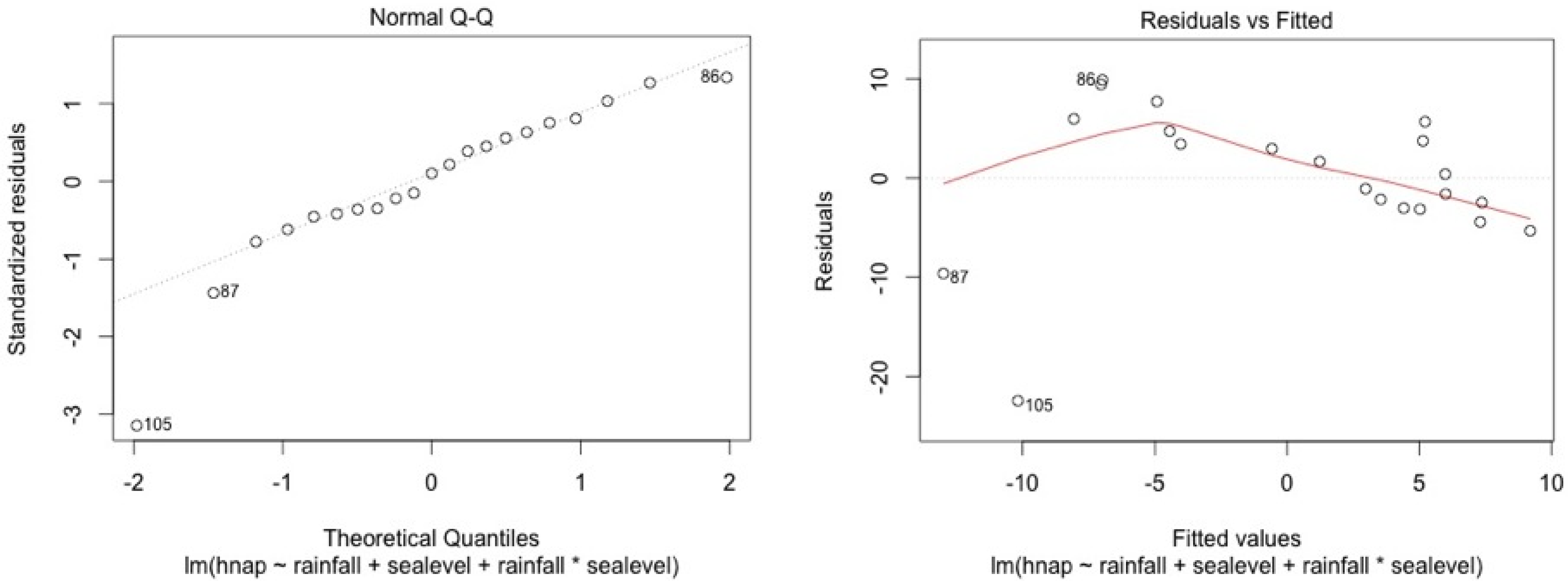
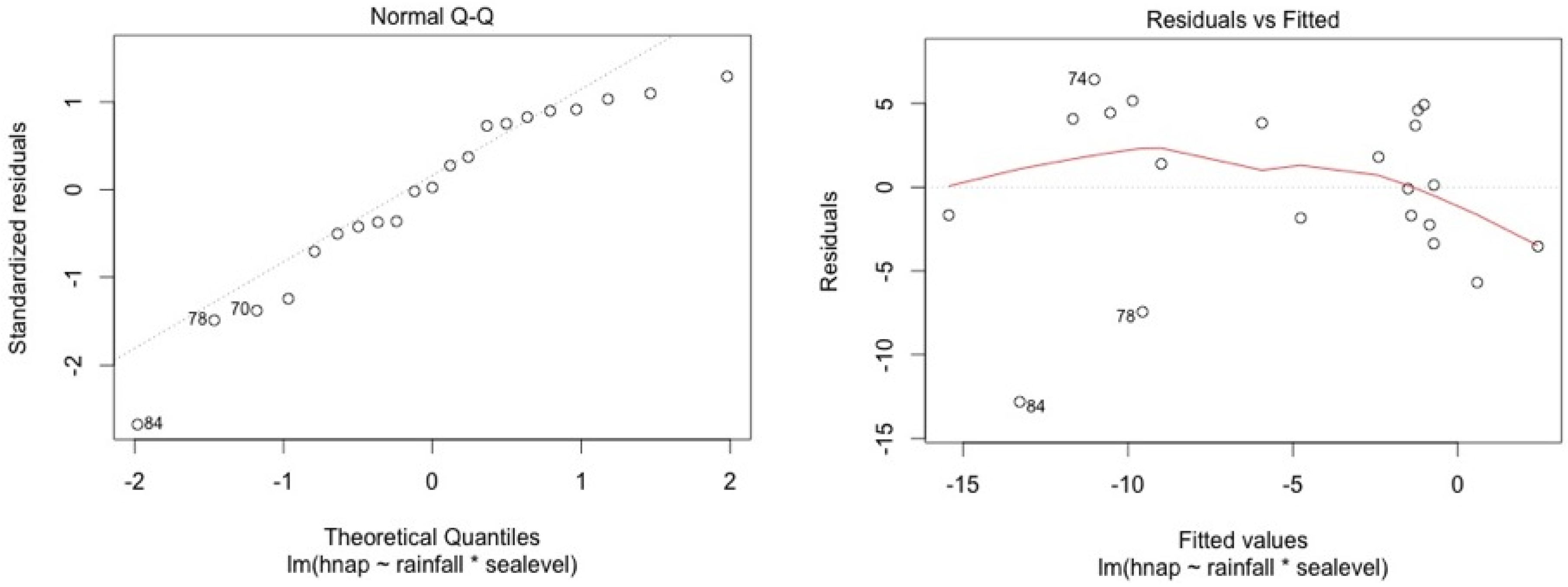
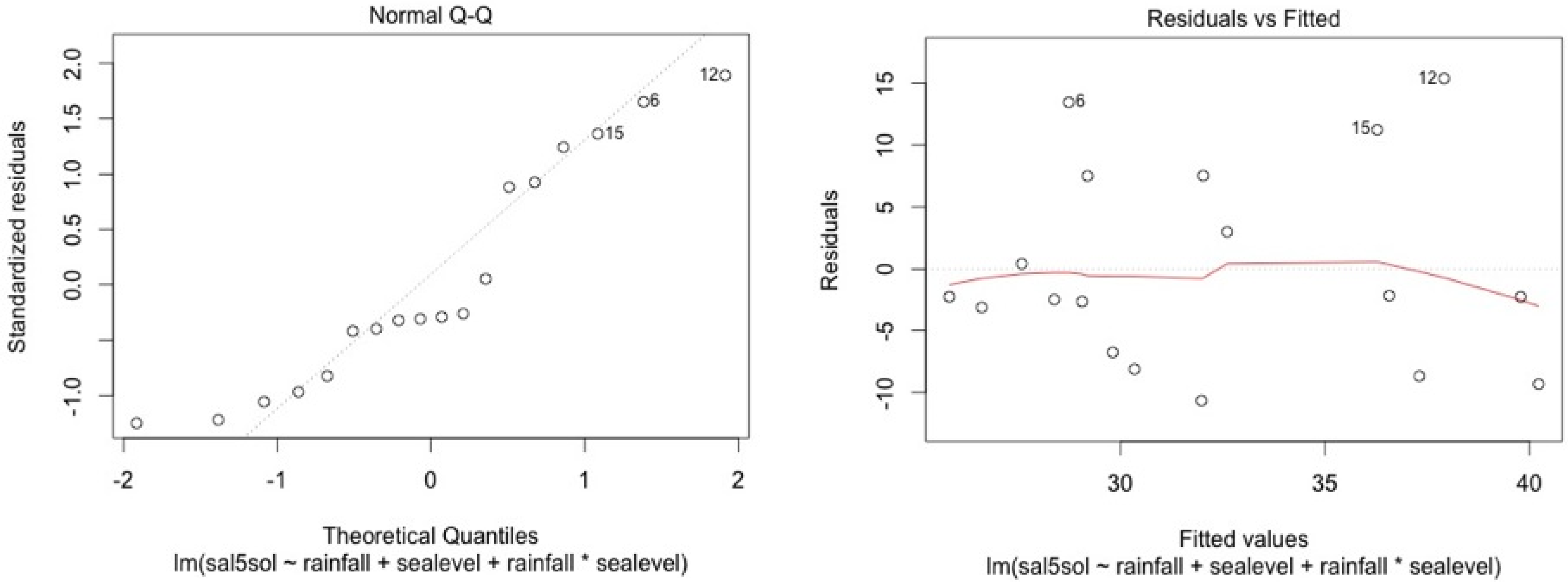
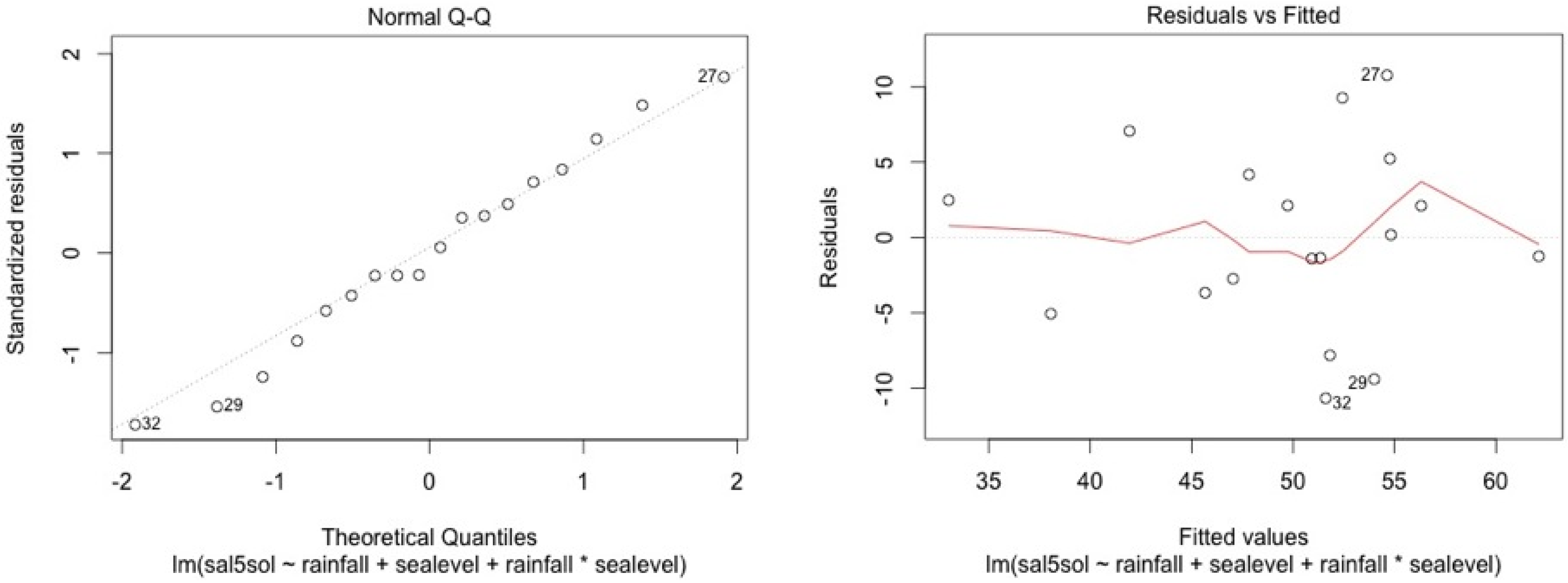
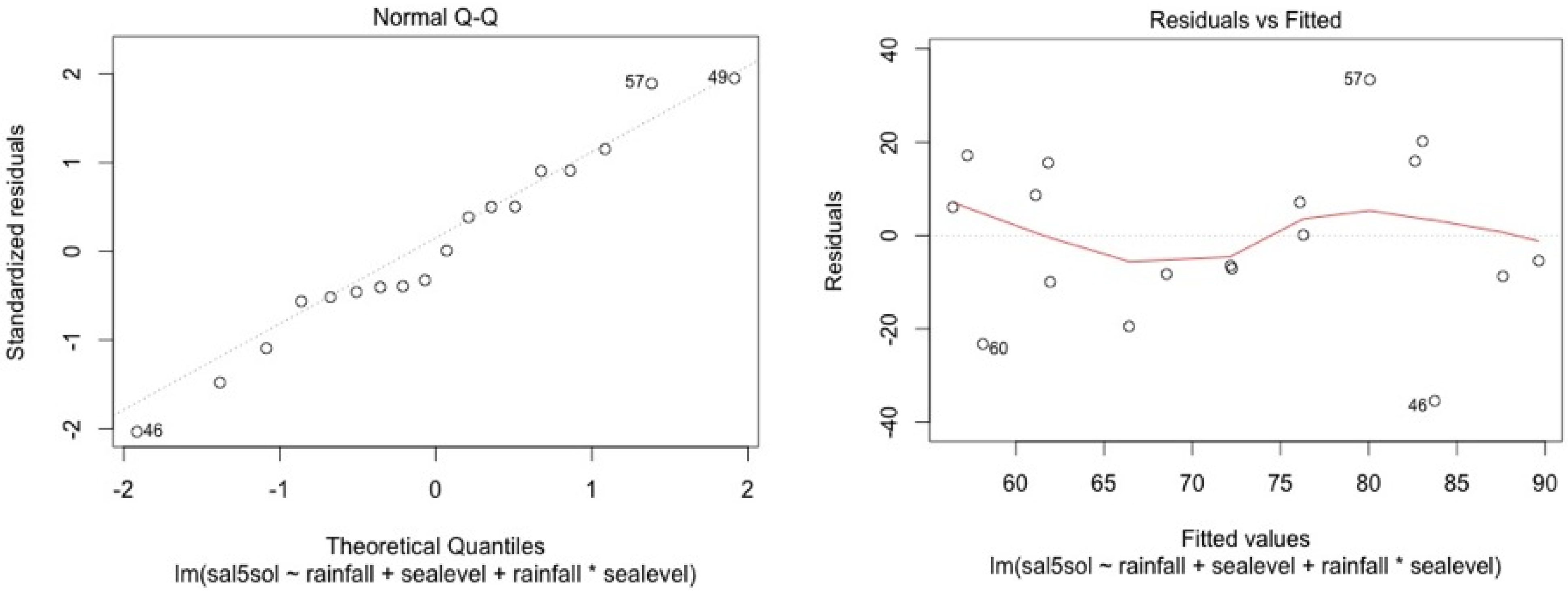
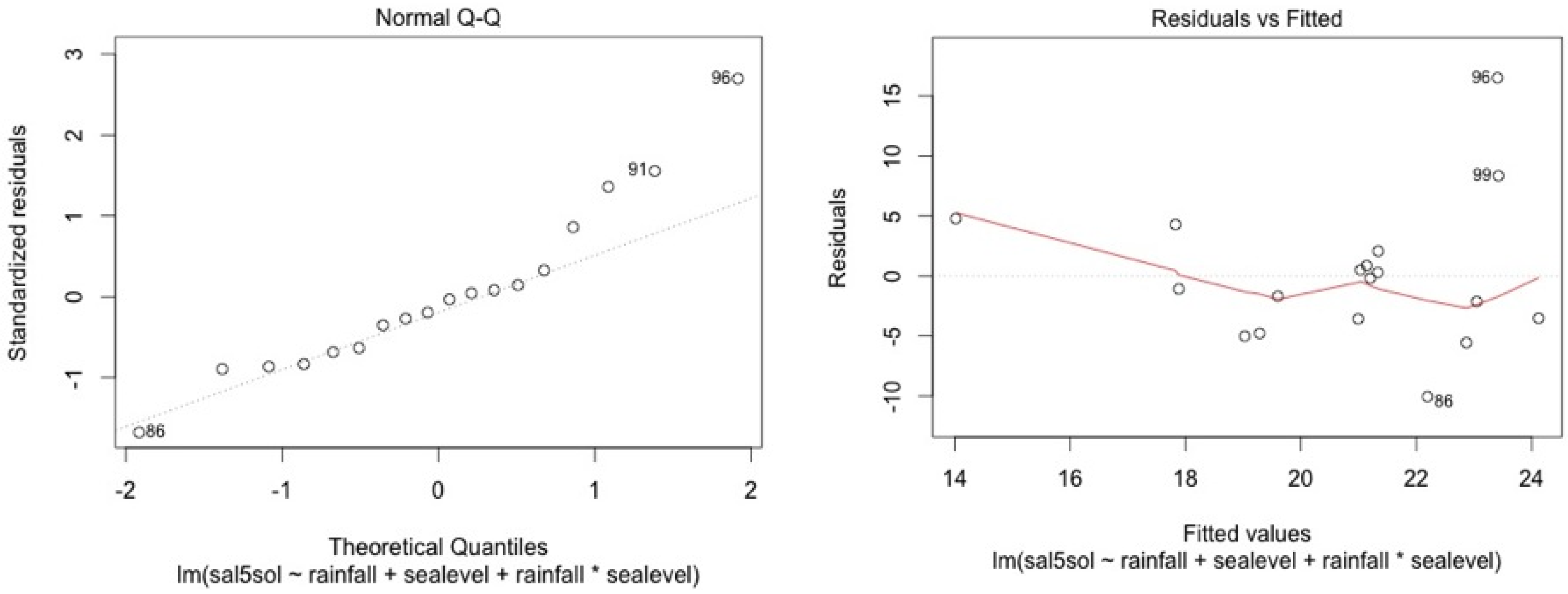
ANCOVA—Residuals’ analysis
- A table with p-value of normality test of the residuals for each model
- 2 plots for a graphic analysis of the residuals:
- ○
- Standardized residuals vs. Theorical quantiles
- ○
- Residuas vs. Predicted values
| Stations | Water Level | Top Soil Salinity |
|---|---|---|
| 5SF | 0.11 | 0.13 |
| 4SF | 0.06 | 0.15 |
| 2MF | 0.09 | 0.95 |
| 1MF | 0.34 | 0.12 |
| 3MF | 0.09 | 0.96 |










References and Notes
- Thom, B.G. Mangrove ecology: A geomorphological perspective. In Mangrove ecosystems in Australia; Clough, B.F., Ed.; Australian National University Press: Camberra, Australia, 1984; pp. 3–17. [Google Scholar]
- Lugo, A.E.; Brown, S.; Brinson, M.M. Forested Wetlands. Ecosystems of the World; Elsevier Science Publishers: Amsterdam, The Netherlands, 1989. [Google Scholar]
- Watson, J.G. Mangrove Forests of the Malay Peninsula; Fraser and Neave: Singapore, 1928; p. 275. [Google Scholar]
- Davis, J.H.J. The Ecology and Geologic Role of Mangroves in Florida; Carnegie Institution of Washington: Washington, DC, USA, 1940. [Google Scholar]
- Chapman, V.J. The 1939 Cambridge University expedition to Jamaica. J. Linn. Soc. Lond. 1944, 52, 407–533. [Google Scholar] [CrossRef]
- Macnea, W. A general account of the fauna and flora of the mangrove swamps and forests in the Indo-Pacific region. Adv. Mar. Biol. 1968, 6, 73–270. [Google Scholar]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Ann. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Ball, M. Ecophysiology of mangroves. Trees 1988, 2, 129–142. [Google Scholar] [CrossRef]
- Ball, M. Mangrove species richness in relation to salinity and waterlogging: a case study along the Adelaide River floodplain, northern Australia. Glob. Ecol. Biogeogr. Lett. 1998, 7, 73–82. [Google Scholar] [CrossRef]
- Santos, I.; Eyre, B.D.; Huettel, M. The driving forces of porewater and groundwater flow in permeable coastal sediments: A review. Estuar. Coast. Shelf Sci. 2012, 98, 1–15. [Google Scholar] [CrossRef]
- Anschutz, P.; Smith, T.; Mouret, A.; Deborde, J.; Bujan, S.; Poirier, D.; Lecroart, P. Tidal Sands as Biogeochemical Reactors. Estuar. Coast. Shelf Sci. 2009, 84, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Kjerfve, B. Manual for Investigation of Hydrological Processes in Mangrove Ecosystems; UNESCO/UNDP Regional Project “Mangrove Ecosystems in Asia and the Pacific” Report RAS/79/002 and RAS/86/120; United Nations Educational, Scientific, and Cultural Organization (UNESCO): Paris, France, 1990; pp. 1–74. [Google Scholar]
- Corenblit, D.; Steiger, J.; Charrier, G.; Darrozes, J.; Dousseau, J.; González, E.; Gurnell, A.M.; Julien, F.; Lambs, L.; Larrue, S.; et al. The biogeomorphological life cycle of poplars during the fluvial biogeomorphological succession: A special focus on Populus nigra L. Earth Surf. Process. Landf. 2014, 39, 546–563. [Google Scholar] [CrossRef] [Green Version]
- Imbert, D. Organisation Spatio-Temporelle des Communautés Végétales du Grand Cul-de-sac Marin (Guadeloupe). PhD Thesis, Montpellier, France, 1985. [Google Scholar]
- Imbert, D.; Rousteau, A.; Scherrer, P. Ecology of mangrove growth and recovery in the Lesser Antilles: State of knowledge and basis for restoration projects. Restor. Ecology 2000, 8, 230–236. [Google Scholar] [CrossRef]
- Schaeffer-Noveli, Y.; Mesquita, H.S.L.; Cintron-Molero, G. The Cananeia Lagoon estuarine system, Sao Paulo, Brazil. Estuaries 1990, 13, 193–203. [Google Scholar] [CrossRef]
- Craig, H. Isotopic variations in meteoric waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef] [PubMed]
- Dansgaard, W. Stable isotopes in precipitation. Tellus 1964, 16, 436–468. [Google Scholar] [CrossRef]
- Wagner, F.; Rossi, V.; Stahl, C.; Bonal, D.; Hérault, B. Water availability is the main climate driver of neotropical tree growth. PLos ONE 2012, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshki, S.R.; DeLaune, R.D.; Patrick, W.H., Jr. Differential response of selected mangroves to soil flooding and salinity: Gas exchange and biomass partitioning. Can. J. For. Res. 1990, 20, 869–874. [Google Scholar] [CrossRef]
- Imbert, D.; Portecop, J. Production de litière dans la mangrove du Grand Cul-de-sac Marin (Guadeloupe). Oecol. Plant 1986, 7, 379–396. [Google Scholar]
- Cintrón, G.; Lugo, A.E.; Pool, D.J.; Morris, G. Mangroves of arid environments in Puerto Rico and adjacent islands. Biotropica 1978, 10, 110–121. [Google Scholar] [CrossRef]
- Flower, J.M.; Imbert, D. Recovery deficiency following tree mortality in mangroves of two Caribbean islands: Field survey and statistical classification. Wetl. Ecol. Manag. 2006, 14, 185–199. [Google Scholar] [CrossRef]
- Imbert, D.; Flower, J.M. Les sols et la végétation de la zone côtière inondable au nord de Port-au-Prince (Haïti). Conjonctions Revue franco-haïtienne de l’Institut Français d’Haïti 2006, 213/214, 33–47. (In French) [Google Scholar]
- Lambs, L.; Muller, E.; Fromard, F. The Guianese paradox: How can the freshwater outflow from the Amazon increase the salinity of the Guianan shore? J. Hydrol. 2007, 342, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Lambs, L.; Muller, E.; Fromard, F. Mangrove trees growing in a very saline condition but not using seawater. Rapid Commun. Mass Spectrom. 2008, 22, 2835–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986; p. 419. [Google Scholar]
- Medina, E.; Cuevas, E.; Lugo, A. Nutrient and salt relations of Pterocarpus officinalis L. in coastal wetlands of the Caribbean: Assessment through leaf and soil analyses. Trees 2007, 21, 321–327. [Google Scholar] [CrossRef]
- Dulormne, M.; Musseau, O.; Muller, F.; Toribio, A.; Bâ, A. Effects of NaCl on growth, water status, N2 fixation, and ion distribution in Pterocarpus officinalis seedlings. Plant Soil 2010, 327, 23–34. [Google Scholar] [CrossRef]
- Imbert, D.; Ménard, S. Structure de la végétation et production primaire dans la mangrove de la Baie de Fort-de-France, Martinique. Biotropica 1997, 29, 413–426. [Google Scholar] [CrossRef]
- Eusse, A.M.; Aide, T.M. Patterns of litter production across a salinity gradient in a Pterocarpus officinalis tropical wetland. Plant Ecol. 1999, 145, 307–315. [Google Scholar] [CrossRef]
- Migeot, J.; Imbert, D. Phenology and production of litter in a Pterocarpus officinalis (Jacq.) swamp forest of Guadeloupe (Lesser Antilles). Aquat. Bot. 2012, 101, 18–27. [Google Scholar] [CrossRef]
- Bompy, F.; Lequeue, G.; Imbert, D.; Dulormne, M. Increasing fluctuations of soil salinity affect seedling growth performances and physiology in three Neotropical mangrove species. Plant Soil 2014, 380, 399–413. [Google Scholar] [CrossRef]
- Bompy, F.; Imbert, D.; Dulormne, M. Impact patterns of soil salinity variations on the survival rate, growth performances, and physiology of Pterocarpus officinalis seedlings. Trees 2015, 29, 119–128. [Google Scholar] [CrossRef]
- Colón-Rivera, R.J.; Feagin, R.A.; West, J.B.; López, N.B.; Benítez-Joubert, R.J. Hydrological modification, saltwater intrusion, and tree water use of a Pterocarpus officinalis swamp in Puerto Rico. Estuar. Coast. Shelf Sci. 2014, 147, 156–167. [Google Scholar] [CrossRef]
- Imbert, D.; Saur, E.; Bonhême, I.; Roseau, V. Traditional Taro (Colocasia esculenta) cultivation in the swamp forest of Guadeloupe (F.W.I.): Impact on forest structure and plant biodiversity. Revue d'Écologie 2004, 59, 181–189. [Google Scholar]
- Alvarez-Lopez, M. Ecology of Pterocarpus officinalis forested wetlands in Puerto Rico. In Ecosystems of the Worls 15. Forested Wetlands; Lugo, A.E., Brinson, M., Brown, S., Eds.; Elservier: Amsterdam, The Netherlands, 1990; pp. 251–265. [Google Scholar]
- Imbert, D.; Delbé, L. Ecology of fire-influenced Cladium jamaicense marshes in Guadeloupe, Lesser Antilles. Wetlands 2006, 26, 289–297. [Google Scholar] [CrossRef]
- Saint-Etienne, L.; Paul, S.; Imbert, D.; Dulormne, M.; Muller, F.; Toribio, A.; Plenchette, C.; Bâ, A.M. Arbuscular mycorrhizal soil infectivity in a stand of the wetland tree Pterocarpus officinalis along a salinity gradient. For. Ecol. Manag. 2006, 232, 86–89. [Google Scholar]
- Migeot, J.; Imbert, D. Structural and floristic patterns in tropical swamp forests: A case study from the Pterocarpus officinalis (Jacq.) forest in Guadeloupe, French West Indies. Aquat. Bot. 2011, 94, 1–8. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. (Eds.) Climate Change 2013: The Physical Science Basis—Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambs, L.; Bompy, F.; Imbert, D.; Corenblit, D.; Dulormne, M. Seawater and Freshwater Circulations through Coastal Forested Wetlands on a Caribbean Island. Water 2015, 7, 4108-4128. https://doi.org/10.3390/w7084108
Lambs L, Bompy F, Imbert D, Corenblit D, Dulormne M. Seawater and Freshwater Circulations through Coastal Forested Wetlands on a Caribbean Island. Water. 2015; 7(8):4108-4128. https://doi.org/10.3390/w7084108
Chicago/Turabian StyleLambs, Luc, Félix Bompy, Daniel Imbert, Dov Corenblit, and Maguy Dulormne. 2015. "Seawater and Freshwater Circulations through Coastal Forested Wetlands on a Caribbean Island" Water 7, no. 8: 4108-4128. https://doi.org/10.3390/w7084108
APA StyleLambs, L., Bompy, F., Imbert, D., Corenblit, D., & Dulormne, M. (2015). Seawater and Freshwater Circulations through Coastal Forested Wetlands on a Caribbean Island. Water, 7(8), 4108-4128. https://doi.org/10.3390/w7084108