
Use of Ecohydraulic-Based Mesohabitat Classification and Fish Species Traits for Stream Restoration Design

Abstract
:1. Introduction
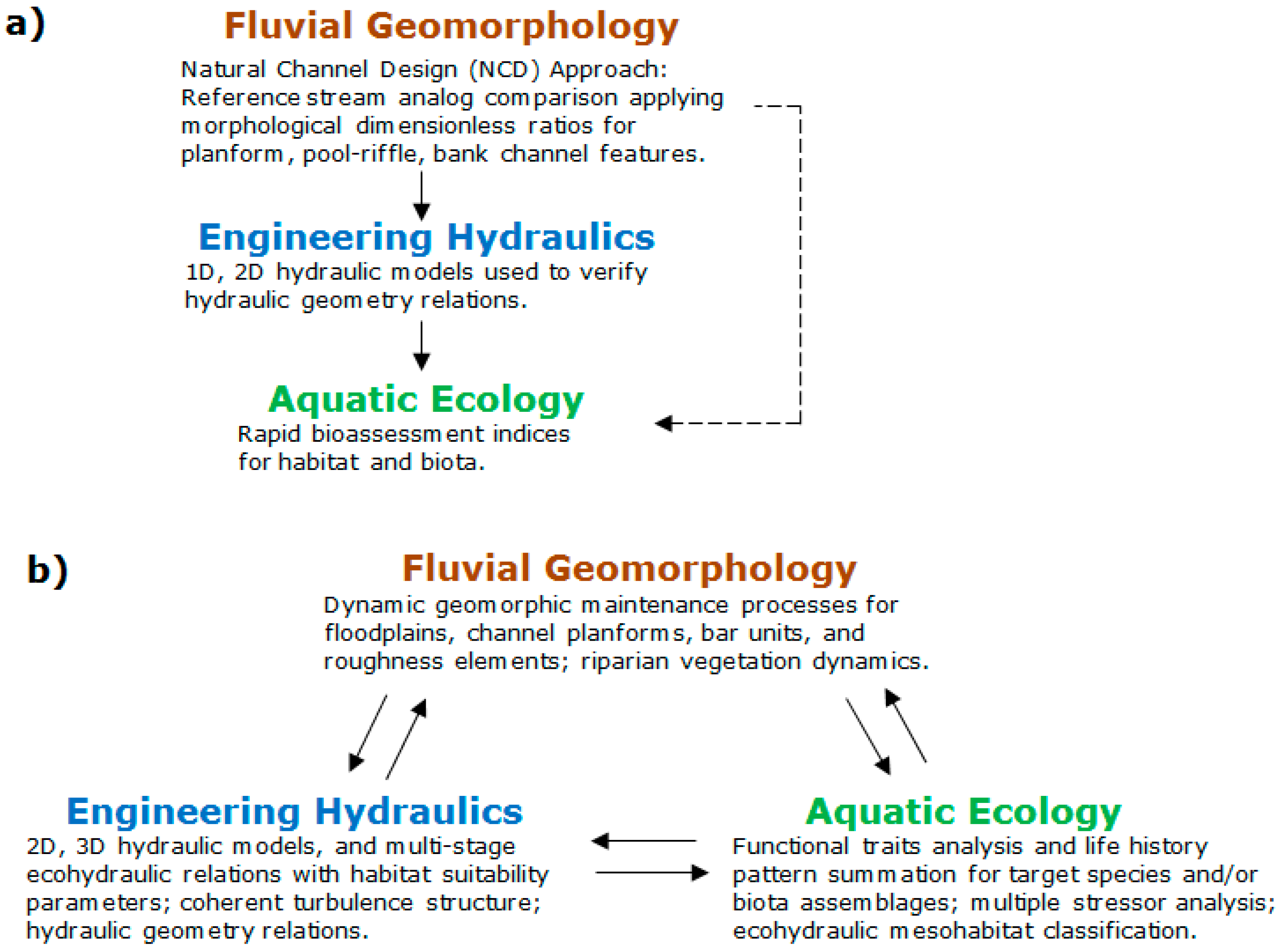
1.1. Ecological Concepts: Broad Applications for Stream Restoration
1.2. Habitat and Biological Assessments
1.3. Hydraulic Habitat Modeling
1.4. Developing Ecological Criteria for Stream Restoration Design
- Applied geomorphic processes for mesohabitat self-maintenance;
- Mesohabitat-scale ecological concepts;
- Species functional traits relationships with mesohabitats; and
- Ecohydraulic-based, multiple flow-stage mesohabitat classification.
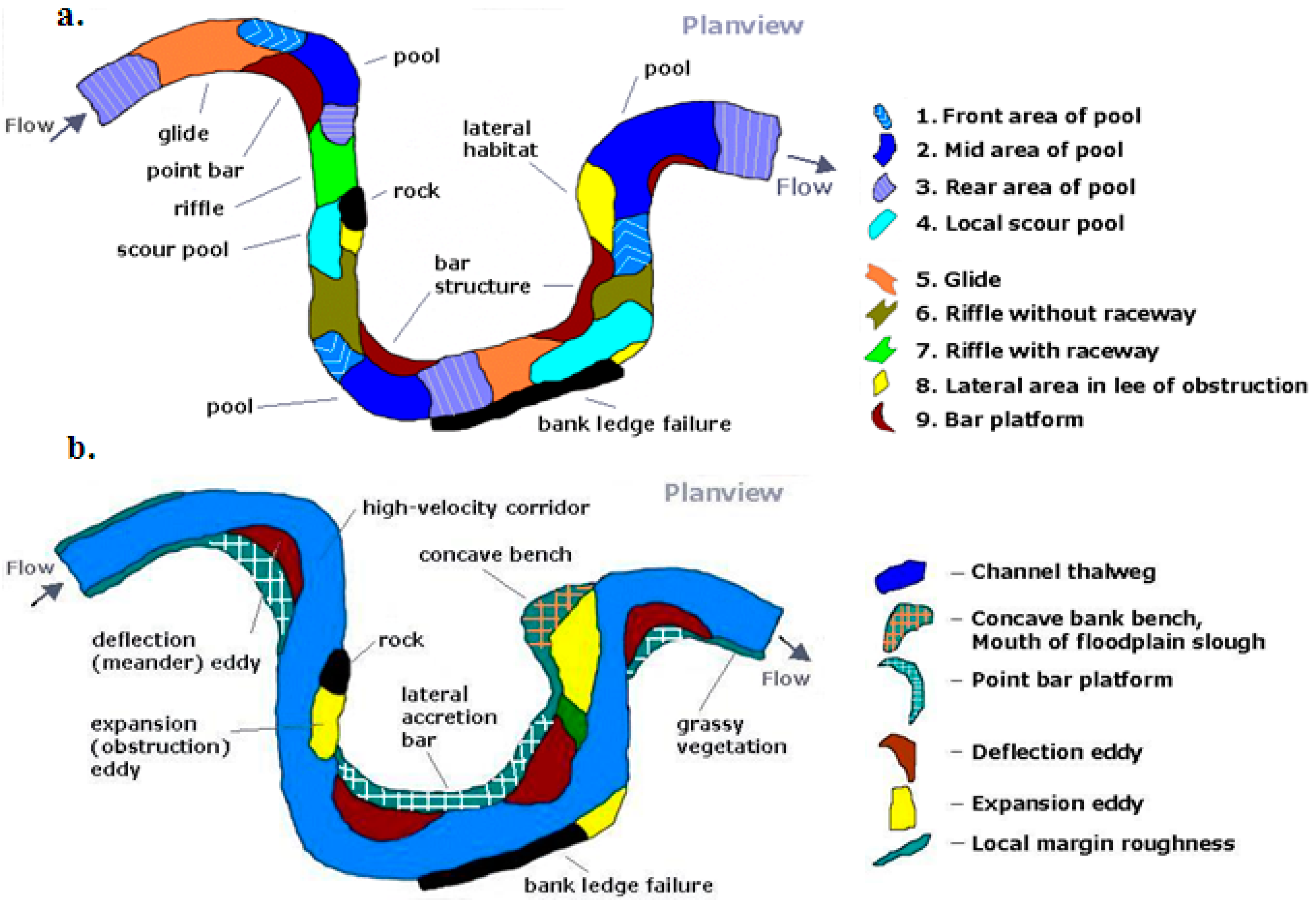
2. Applied Geomorphic Processes for Mesohabitat Maintenance
- Lateral Migrating Floodplains: Lateral point-bar accretion consisting of progressive sediment deposition on the convex bank of a meander bend from helical and divergent flow through the bend [136], which creates elevated and vegetated remnant point bar surfaces;
- Counterpoint Floodplains: Counterpoint accretion occurs within a hydraulic recirculation zone (or large stream eddy) formed against the upstream limb of the convex bank in sharply curving and active migrating bends allowing for fine sediment deposition, which over time form an elevated and vegetated concave bank bench [134]; and
- Abandoned Channel Floodplains: Abandoned-channel accretion occurs in actively migrating channels creating cutoffs followed by overbank vertical accretion, which depending on sediment deposition and location to the active channel the floodplains features include elevated and vegetated remnant channels, and a backwater slough near the mouth of the original channel [137].
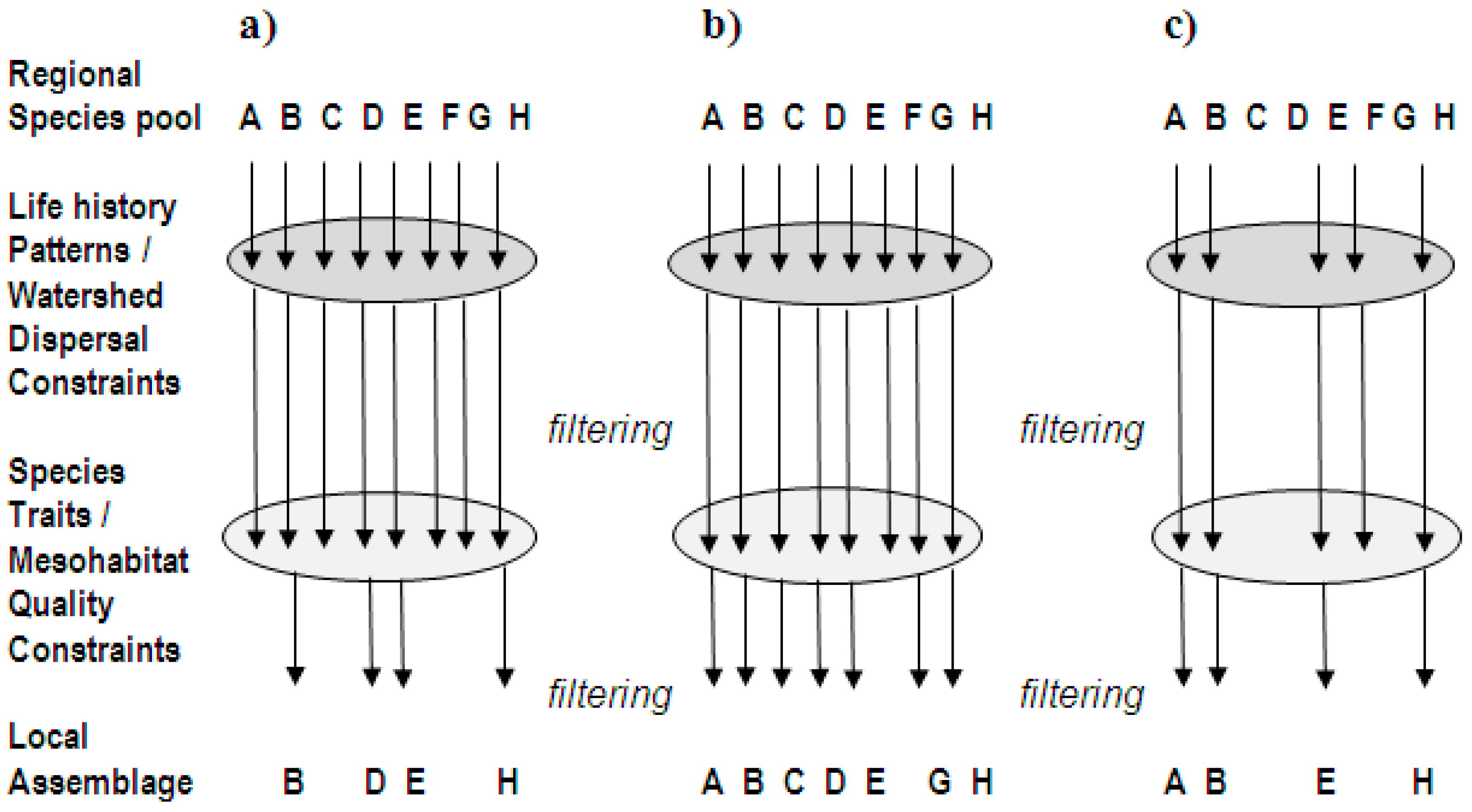
3. Applied Ecological Concepts at the Mesohabitat Scale
- habitat selection and relative availability of structural requirements in terms of complexity, duration, frequency, and juxtaposition;
- abiotic disturbances such as extremes in flows and temperature; and physiological tolerances to local water quality conditions;
- food availability and food-space-cover relationships;
- resource specialization of fish species; species traits related to body morphology and mode of foraging;
- predator-prey interactions and trophic controls, intra-and inter-specific competitive interactions related to individual size and behavior, and population densities; and
- immigration-extinction and recolonization dynamics.
4. Species Functional Traits
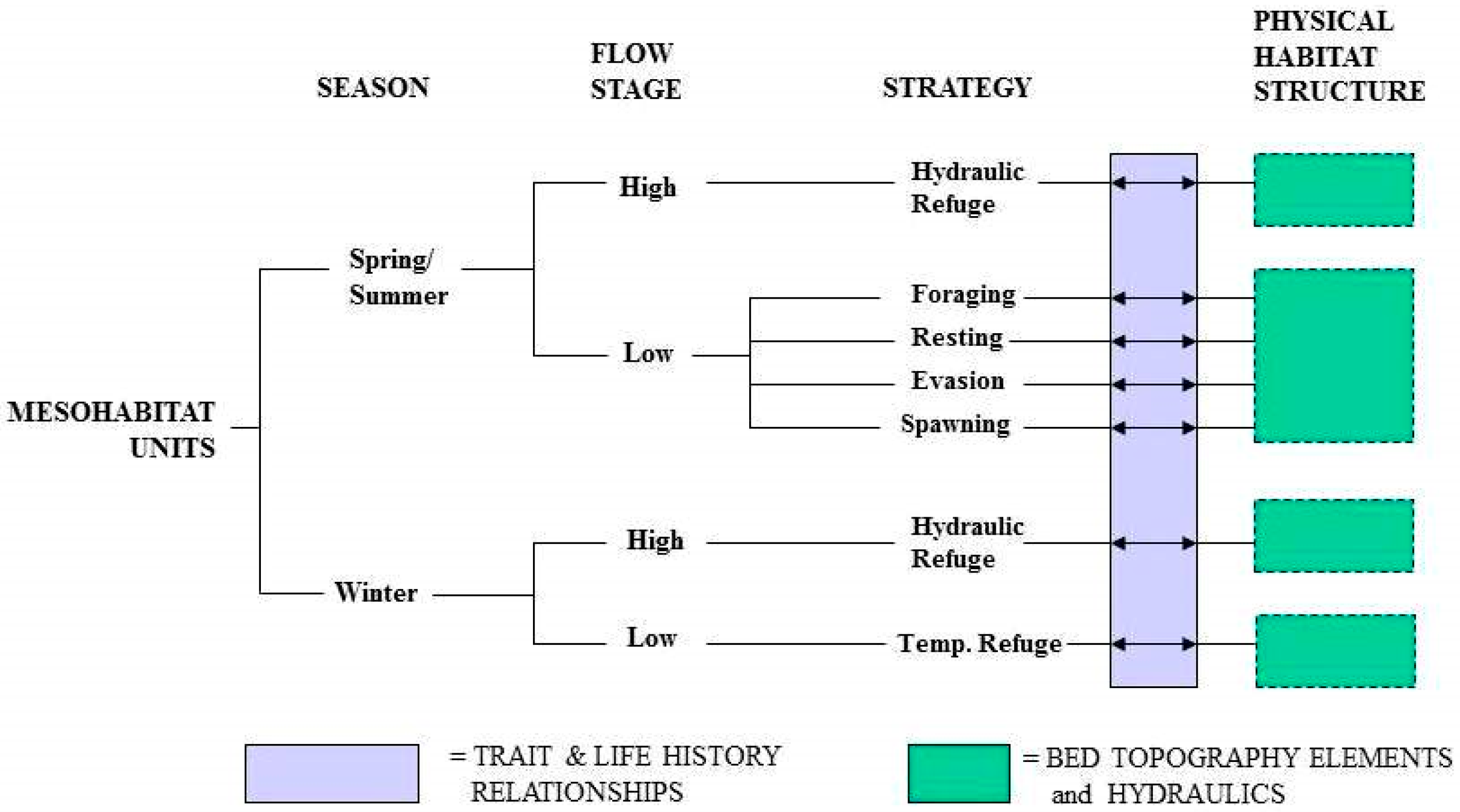
5. Multi-Stage Ecohydraulics-Based Mesohabitat Classification
5.1. Framework for Classification of Multi-Stage Ecohydraulics-Based Mesohabitat Units
5.2. Mesohabitat Units at Low-Flow Stages
5.3. Mesohabitat Units at High-Flow Stages
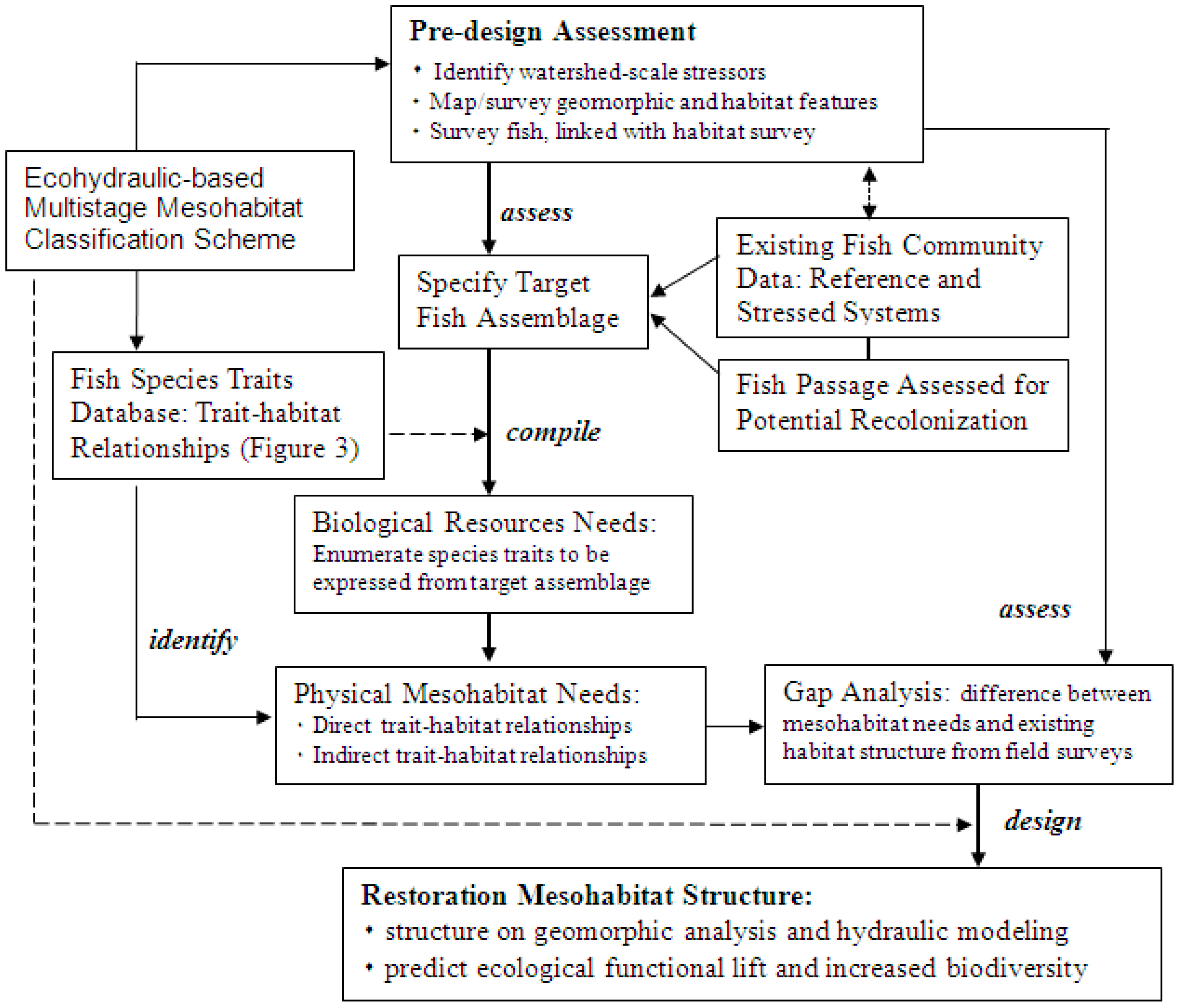
6. Ecohydraulic-Based Stream Restoration: Proposed Application
- (1)
- Enumerate the functional traits expressed per fish species in the target community, summarizing the frequency of traits expressed because various traits will be expressed by multiple species in the target assemblage;
- (2)
- Summarize physical habitat needs based on the enumerated functional traits of the target assemblage, which constitutes the functional traits analysis (Figure 3); it should be noted that habitat needs could be direct or indirect, where a direct need is the explicit space occupancy for a specified trait expression (e.g., mid-pool resting) and indirect need is space used by another organism needed for a target species, for example pool-front feeding with the food source (benthic macroinvertebrates) generated in the adjacent upstream riffle;
- (3)
- (4)
- Compare the existing mesohabitat units from the stream surveys with a set of habitat units derived from the functional traits analysis, identifying any departures between the two (gap analysis); and
- (5)
- Of the mesohabitat unit types found to be limited, summarize measures of physical structural heterogeneity and complexity from field surveys, and design the restored habitat based on trait-habitat relationships, and geomorphic and hydraulic principles.
7. Conclusions
- (1)
- physical habitat space is specifically linked with species functional traits so that mesohabitat units form the basic “building blocks” for stream restoration design, recognizing that habitat unit use shifts with season and flow stage governed by life histories and trait strategies;
- (2)
- high-flow refugia must be considered in restoration design, characterized by elevated geomorphic surfaces at the channel-floodplain interface;
- (3)
- mesohabitat units must be associated with mesoscale 3D hydraulic and turbulence patterns that relate to trait strategies, i.e., feeding and spawning positions; and
- (4)
- the restoration design process must recognize the self-maintenance geomorphic processes for both low- and high-flow mesohabitats, the spatial-process linkages for reach-scale bed topography, bank, and elevated floodplain-channel geomorphic surfaces.
Acknowledgments
Conflicts of Interest
References
- Alexander, G.G.; Allan, J.D. Stream restoration in the upper Midwest, U.S.A. Restor. Ecol. 2006, 14, 595–604. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Sudduth, E.B.; Palmer, M.A.; Allan, J.D.; Meyer, J.L.; Alexander, G.; Follstad Shah, J.; Hassett, B.; Jenkinson, R.; Lave, R.; et al. Restoring rivers one reach at a time: Results from a survey of U.S. river restoration practitioners. Restor. Ecol. 2007, 15, 82–93. [Google Scholar] [CrossRef]
- Lave, R. Field and Streams: Stream Restoration, Neoliberalism and the Future of Environmental Science; The University of Georgia Press: Athens, GA, USA, 2012; p. 170. [Google Scholar]
- Palmer, M.A.; Hondula, K.L.; Koch, B.J. Ecological restoration of stream and rivers: Shifting strategies and shifting goals. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 247–269. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A.; Allan, J.D.; Alexander, G.; Barnas, K.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.; Follstad-Shah, J.; et al. Synthesizing U.S. river restoration efforts. Science 2005, 308, 636–637. [Google Scholar] [CrossRef] [PubMed]
- Rosgen, D. Applied Stream Geomorphology; Wildland Hydrology: Pagoda Springs, CO, USA, 1996. [Google Scholar]
- Hey, R.D. Fluvial geomorphological methodology for natural stable channel design. J. Am. Water Resour. Assoc. 2006, 42, 357–374. [Google Scholar] [CrossRef]
- Jenkinson, R.G.; Barnas, K.A.; Braatne, J.H.; Bernhardt, E.S.; Palmer, M.A.; Allan, J.D.; The National River Restoration Science Synthesis. Stream restoration databases and case studies: A guide to information resources and their utility in advancing the science and practice of restoration. Restor. Ecol. 2006, 14, 177–186. [Google Scholar] [CrossRef]
- Rosgen, D.L. Natural channel design: Fundamental concepts, assumptions, and methods. In Stream Restoration in Dynamic Fluvial Systems: Scientific Approaches, Analysis, and Tools; Simon, A., Bennett, S.J., Castro, J.M., Eds.; Geophysical Monograph Series 194; American Geophysical Union: Washington, DC, USA, 2011; pp. 69–94. [Google Scholar]
- Rosgen, D.L. Rosgen geomorphic channel design. In Part 654 Stream Restoration Design National Engineering Handbook; U.S.D.A. National Resources Conservation Service: Washington, DC, USA, 2007; pp. 1–76. [Google Scholar]
- Shields, F.D.; Copeland, R.R.; Klingeman, P.C.; Doyle, M.W.; Simon, A. Design for Stream Restoration. ASCE J. Hydraul. Eng. 2003, 129, 575–584. [Google Scholar] [CrossRef]
- Copeland, R.R.; McComas, D.N.; Thorne, C.R.; Soar, P.J.; Jonas, M.M.; Fripp, J.B. Hydraulic Design of Stream Restoration Projects; ERDC/CHL TR-01-28; U.S. Army Corps of Engineers: Washington, DC, USA, 2001; p. 109.
- Downs, P.W.; Thorne, C.R. Rehabilitation of a lowland river: Reconciling flood defence with habitat diversity and geomorphological sustainability. J. Environ. Manag. 2000, 58, 249–268. [Google Scholar] [CrossRef]
- Niezgoda, S.L.; Johnson, P.A. Modeling the Long Term Impacts of Using Rigid Structures in Stream Channel Restoration. J. Am. Water Resour. Assoc. 2006, 42, 1597–1613. [Google Scholar] [CrossRef]
- Miller, J.R.; Kochel, R.C. Assessment of channel dynamics, in-stream structures and post-project channel adjustments in North Carolina and its implications to effective stream restoration. Environ. Earth Sci. 2010, 59, 1681–1692. [Google Scholar] [CrossRef]
- Buchanan, B.P.; Nagle, G.N.; Walter, M.T. Long-term monitoring of a stream restoration project in central New York. River Res. Appl. 2014, 30, 245–258. [Google Scholar] [CrossRef]
- Kondolf, G.M. Five elements for effective evaluation of stream restoration. Restor. Ecol. 1995, 3, 133–136. [Google Scholar] [CrossRef]
- Aarts, B.G.W.; Van Den Brink, F.W.B.; Nienhuis, P.H. Habitat loss as the main cause of the slow recovery of fish faunas of regulated large rivers in Europe: The transversal floodplain gradient. River Res. Appl. 2004, 20, 3–23. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Herricks, E.E. Evaluation of pool-riffle naturalization structures on habitat complexity and the fish community in an urban Illinois stream. River Res. Appl. 2007, 23, 451–466. [Google Scholar] [CrossRef]
- Tullos, D.D.; Penrose, D.L.; Jennings, G.D.; Cope, W.G. Analysis of functional traits in reconfigured channels: Implications for the bioassessment and disturbance of river restoration. J. N. Am. Benthol. Soc. 2009, 28, 80–92. [Google Scholar] [CrossRef]
- Palmer, M.A.; Menninger, H.L.; Bernhardt, E. River restoration, habitat heterogeneity and biodiversity: A failure of theory or practice? Freshw. Biol. 2010, 55, 205–222. [Google Scholar] [CrossRef]
- Haase, P.; Hering, D.; Jahnig, S.C.; Lorenz, A.W.; Sundermann, A. The impact of hydromorphological restoration on the river ecological status: A comparison of fish, benthic invertebrates, and macrophytes. Hydrobiologia 2013, 704, 475–488. [Google Scholar] [CrossRef]
- Doll, B.A.; Jennings, G.D.; Spooner, J.; Penrose, D.L.; Usset, J.L. Evaluating the eco-geomorphological condition of restored streams using visual assessment and macroinvetebrate metrics. J. Water Resour. Assoc. 2015, 51, 68–83. [Google Scholar] [CrossRef]
- Doll, B.A.; Jennings, G.D.; Spooner, J.; Penrose, D.L.; Usset, J.L.; Blackwell, J.; Fernandez, M. Can rapid assessments predict the biotic condition of restored streams? Water 2016, 8, 143. [Google Scholar] [CrossRef]
- Palmer, M.A.; Allan, J.D.; Meyer, J.; Bernhardt, E.S. River restoration in the twenty-first century: Data and experimental knowledge to inform future efforts. Restor. Ecol. 2007, 15, 472–481. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Hassett, B.A.; Cada, P.; Bernhardt, E.S. Testing the field of dreams hypothesis: Functional responses to urbanization and restoration in stream ecosystems. Ecol. Appl. 2011, 21, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Sedell, J.R.; Reeves, G.H.; Hauer, F.R.; Stanford, J.A.; Hawkins, C.P. Role of refugia in recovery from disturbances: Modern fragmented and disconnected river systems. Environ. Manag. 1990, 14, 711–724. [Google Scholar] [CrossRef]
- Schlosser, I.J. Environmental variation, life history attributes, and community structure in stream fishes: Implications for environmental management and assessment. Environ. Manag. 1990, 14, 621–628. [Google Scholar] [CrossRef]
- Kondolf, G.M. Geomorphological stream channel classification in aquatic habitat restoration: Uses and limitations. Aquat. Conserv. Mar. Freshw. Ecosyst. 1995, 5, 127–141. [Google Scholar] [CrossRef]
- Sundermann, A.; Stoll, S.; Hasse, P. River restoration success depends on the species pool of the immediate surroundings. Ecol. Appl. 2009, 21, 1962–1971. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. River restoration: The fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecol. Appl. 2011, 21, 1921–1931. [Google Scholar] [CrossRef]
- Schwartz, J.S. Stream Habitat Characterized by Stage-specific Flows and Three-dimensional Geomorphological Complexity: Development of Ecological Criteria for Stream Restoration Design. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, June 2002; p. 246. [Google Scholar]
- Lake, P.S.; Bond, N.; Reich, P. Linking ecological theory with stream restoration. Freshw. Biol. 2007, 52, 597–615. [Google Scholar] [CrossRef]
- Schwartz, J.S. Influence of Geomorphology and Land Use on Distribution and Abundance of Salmonids in a Coastal Oregon Basin. Master’s Thesis, Oregon State University, Corvallis, OR, USA, June 1991; p. 207. [Google Scholar]
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones. BioScience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Frothingham, K.M.; Rhoads, B.L.; Herricks, E.E. A multiscale conceptual framework for integrated eco-geomorphological research to support stream naturalization in the agricultural Midwest. Environ. Manag. 2002, 29, 16–23. [Google Scholar] [CrossRef]
- Beechie, T.J.; Sear, D.A.; Olden, J.D.; Pess, G.R.; Buffington, J.M.; Moir, H.; Roni, P.; Pollock, M.M. Process-based principles for restoring river ecosystems. BioScience 2010, 60, 209–222. [Google Scholar] [CrossRef]
- Frissell, C.A.; Liss, W.J.; Warren, C.E.; Hurley, M.D. A hierarchical framework for stream habitat classification: Viewing streams in a watershed context. Environ. Manag. 1986, 10, 199–214. [Google Scholar] [CrossRef]
- Pringle, C.M.; Naiman, R.J.; Bretschko, G.; Karr, J.R.; Oswood, M.W.; Webster, J.R.; Welcome, R.L.; Winterbourn, M.J. Patch dynamics in lotic systems: The stream as a mosaic. J. N. Am. Benthol. Soc. 1988, 7, 503–524. [Google Scholar] [CrossRef]
- Townsend, C.R.; Doledec, S.; Scarbrook, M.R. Species traits in relation to temporal and spatial heterogeneity in streams: A test of habitat templet theory. Freshw. Biol. 1997, 37, 367–387. [Google Scholar] [CrossRef]
- Townsend, C.R. The patch dynamics concept of stream community ecology. J. N. Am. Benthol. Soc. 1989, 8, 36–50. [Google Scholar] [CrossRef]
- Wu, J.; Loucks, O.L. From balance of nature to hierarchical patch dynamics: A paradigm shift in ecology. Q. Rev. Biol. 1995, 70, 439–466. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Habitat, the template for ecological strategies? J. Anim. Ecol. 1977, 46, 337–365. [Google Scholar] [CrossRef]
- The Federal Interagency Stream Restoration Working Group (15 US Federal Agencies). Stream Corridor Restoration: Principles, Processes, and Practices; GPO Item No. 0120-A; FISRWG: Washington, DC, USA, 1998.
- Yochum, S. Guidance for Stream Restoration; Engineering Technical Note No. 27.3; USDA Natural Resources Conservation Service: Denver, CO, USA, 2014; p. 77.
- Ward, J.V. The four-dimensional nature of lotic ecosystems. J. N. Am. Benthol. Soc. 2089, 8, 3–8. [Google Scholar] [CrossRef]
- Tockner, K.; Pennetzdorfer, D.; Reiner, N.; Schiemer, F.; Ward, J.V. Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river-floodplain system (Danube, Austria). Freshw. Biol. 1999, 41, 521–535. [Google Scholar] [CrossRef]
- Kondolf, G.M.; Boulton, A.J.; O’Daniel, S.; Poole, G.C.; Rahel, F.J.; Stanley, E.H.; Wohl, E.; Bang, A.; Carlstrom, J.; Cristoni, C.; et al. Process-based ecological river restoration: Visualizing three-dimensional connectivity and dynamic vectors to recover lost linkages. Ecol. Soc. 2006, 11, 1–5. [Google Scholar]
- Thorp, J.H.; Thoms, M.C.; Delong, M.D. The riverine ecosystem synthesis: Biocomplexity in river networks across space and time. River Res. Appl. 2006, 22, 123–147. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Wiley, M.J.; Osborne, L.L.; Larimore, R.W. Longitudinal structure of an agricultural prairie river system and its relationship to current stream ecosystem theory. Can. J. Fish. Aquat. Sci. 1989, 47, 373–384. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The flood pulse concept in river-floodplain systems. In Proceedings of the International Large River Symposium, Honey Harbour, Toronto, ON, Canada, 14–21 September 1989; Dodge, D.P., Ed.; Canadian Special Publication of Fisheries and Aquatic Sciences 106. Department of Fisheries and Oceans: Ottawa, ON, Canada, 1989; pp. 110–127. [Google Scholar]
- Jansson, R.; Nilsson, C.; Malmqvist, B. Restoring freshwater ecosystems in riverine landscapes: The roles of connectivity and recovery processes. Freshw. Biol. 2007, 52, 589–596. [Google Scholar] [CrossRef]
- Klocker, C.A.; Kaushal, S.S.; Groffman, P.M.; Mayer, P.M.; Morgan, R.P. Nitrogen uptake and denitrification in restored and unrestored streams in urban Maryland, USA. Aquat. Sci. 2009, 71, 411–424. [Google Scholar] [CrossRef]
- Roley, S.S.; Tank, J.L.; Stephen, M.L.; Johnson, L.T.; Beaulieu, J.J.; Witter, J.D. Floodplain restoration enhances denitrification and reach-scale nitrogen removal in an agricultural stream. Ecol. Appl. 2012, 22, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Marzadri, A.; Tonina, D.; Bellin, A.; Tank, J.L. A hydrologic model demonstrates nitrous oxide emissions depend on streambed morphology. Geophys. Res. Lett. 2014, 41, 5484–5491. [Google Scholar] [CrossRef]
- Palmer, M.A.; Ambrose, R.F.; Poff, N.L.R. Ecological theory and community restoration ecology. Restor. Ecol. 1997, 5, 291–300. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Kershner, J.L.; Bisson, P.A.; Bryant, M.D.; Decker, L.M.; Gregory, S.V.; McCullough, D.A.; Overton, C.K.; Reeves, G.H.; Steedman, R.J.; et al. A hierarchical approach to classifying stream habitat features. Fisheries 1993, 18, 3–12. [Google Scholar] [CrossRef]
- Newbury, R.; Gaboury, M. Exploration and rehabilitation of hydraulic habitats in streams using principals of fluvial behavior. Freshw. Biol. 1993, 29, 195–210. [Google Scholar] [CrossRef]
- Ombredane, D.; Haury, J.; Chapon, P.M. Heterogeneity and typology of fish habitat in the main stream of a Breton coastal river. Hydrobiologia 1995, 300/301, 259–268. [Google Scholar] [CrossRef]
- Bisson, P.A.; Montgomery, D.R. Valley segments, stream reaches, and channel units. In Methods in Stream Ecology; Hauer, R.F., Lambert, G.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 23–42. [Google Scholar]
- Vadas, R.L.; Orth, D.J. Use of physical variables to discriminate visually determined mesohabitat types in North American streams. Rivers 1998, 6, 143–159. [Google Scholar]
- Bain, M.B.; Stephenson, N.J. Aquatic Habitat Assessment, Common Methods; American Fisheries Society: Bethesda, MD, USA, 1999; p. 216. [Google Scholar]
- Kaufmann, P.R.; Levine, P.; Robison, E.G.; Seeliger, C.; Peck, D.V. Quantifying Physical Habitat in Wadeable Streams; EPA/620/R-99/003; U.S. Environmental Protection Agency: Washington, DC, USA, 1999.
- U.S. Forest Service. Stream Inventory Handbook: Levels I and II; U.S. Forest Service, Pacific Northwest Region 6 Office: Portland, OR, USA, 2012; p. 127.
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.EPA 841-B-99-002; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Shields, F.D.; Knight, S.S.; Cooper, C.M. Use of the index of biotic integrity to assess physical habitat degradation in warmwater streams. Hydrobiologia 1995, 312, 191–208. [Google Scholar] [CrossRef]
- Maddock, I. The importance of physical habitat assessment for evaluating river health. Freshw. Biol. 1999, 41, 373–391. [Google Scholar] [CrossRef]
- Metzeling, L.; Chessman, B.; Hardwick, R.; Wong, V. Rapid assessment of rivers using macroinvertebrates: The role of experience, and comparisons with quantitative methods. Hydrobiologia 2003, 510, 39–52. [Google Scholar] [CrossRef]
- Asmus, B.; Magner, J.A.; Vondracek, B.; Perry, J. Physical integrity: The missing link in biological monitoring and TMDLs. Environ. Monit. Assess. 2009, 159, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Knighton, D. Fluvial Forms and Processes; Edward Arnold Publisher: London, UK, 1998; p. 388. [Google Scholar]
- Keller, E.A.; Melhorn, W.N. Rhythmic spacing and origin of pools and riffles. Geol. Soc. Am. Bull. 1978, 89, 723–730. [Google Scholar] [CrossRef]
- Gregory, K.J.; Gurnell, A.M.; Hill, C.T.; Tooth, S. Stability of the pool-riffle sequence in changing river channels. Regul. Rivers Res. Manag. 1994, 9, 35–43. [Google Scholar] [CrossRef]
- Sear, D.A. Sediment transport processes in pool-riffle sequences. Earth Surface Process. Landf. 1996, 21, 241–262. [Google Scholar] [CrossRef]
- Clifford, N.J. Differential bed sedimentology and the maintenance of riffle-pool sequences. Catena 1993, 20, 447–468. [Google Scholar] [CrossRef]
- Carling, P.A.; Orr, H.G. Morphology of riffle-pool sequences in the River Severn, England. Earth Surface Process. Landf. 2000, 25, 369–384. [Google Scholar] [CrossRef]
- Rabeni, C.F.; Jacobson, J.B. The importance of fluvial hydraulics to fish-habitat restoration in low-gradient alluvial streams. Freshw. Biol. 1993, 29, 211–220. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M. Stream Ecology, Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007; p. 436. [Google Scholar]
- Schwartz, J.S.; Herricks, E.E. Fish use of ecohydraulic-based mesohabitat units in a low-gradient Illinois stream: Implications for stream restoration. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 852–866. [Google Scholar] [CrossRef]
- Clifford, N.J.; Harmar, O.P.; Harvey, G.; Petts, G.E. Physical habitat, ecohydraulics and river design: A review and re-evaluation of some popular concepts and methods. Aquat. Conserv. Mar. Freshw. Ecosyst. 2006, 16, 389–408. [Google Scholar] [CrossRef]
- Rhoads, B.L.; Engel, F.L.; Abad, J.D. Pool-riffle design based on geomorphological principles for naturalizing straight channels. In Stream Restoration in Dynamic Fluvial Systems: Scientific Approaches, Analysis, and Tools; Simon, A., Bennett, S.J., Castro, J.M., Eds.; Geophysical Monograph Series 194; American Geophysical Union: Washington, DC, USA, 2011; pp. 367–384. [Google Scholar]
- Doyle, M.W.; Shields, F.D. Compensatory mitigation for streams under the Clean Water Act: Reassessing science and redirecting policy. J. Am. Water Recour. Assoc. 2012, 48, 494–509. [Google Scholar] [CrossRef]
- Harman, W.; Starr, R.; Carter, M.; Tweedy, K.; Clemmons, M.; Suggs, K.; Miller, C. A Function-Based Framework for Stream Assessment and Restoration Projects; EPA 843-K-12-006; U.S. Environmental Protection Agency, Office of Wetlands, Oceans, and Watersheds: Washington, DC, USA, 2012; p. 340.
- Palmer, M.A.; Bernhardt, E.S.; Allan, J.D.; Lake, P.S.; Alexander, G.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.N.; Galat, D.L.; et al. Standards for ecologically successful river restoration. J. Appl. Ecol. 2005, 42, 208–217. [Google Scholar] [CrossRef]
- Woolsey, S.; Capelli, F.; Gonser, T.; Hoehn, E.; Hostmann, M.; Junker, B.; Paetzold, A.; Roulier, C.; Schweizer, S.; Tiegs, S.D.; et al. A strategy to assess river restoration success. Freshw. Biol. 2007, 52, 752–769. [Google Scholar] [CrossRef]
- Pander, J.; Geist, J. Ecological indicators for stream restoration success. Ecol. Indic. 2013, 30, 106–118. [Google Scholar] [CrossRef]
- Wortley, L.; Hero, J.-M.; Howes, M. Evaluating ecological restoration success: A review of the literature. Restor. Ecol. 2013, 21, 537–543. [Google Scholar] [CrossRef]
- Statzner, B.; Gore, J.A.; Resh, V.H. Hydraulic stream ecology: Observed patterns and potential applications. J. N. Am. Benthol. Soc. 1988, 7, 307–360. [Google Scholar] [CrossRef]
- Shields, F.D.; Langendoen, E.J.; Doyle, M.W. Adapting existing models to examine effects of agricultural conservation programs on stream habitat quality. J. Am. Water Resour. Assoc. 2006, 42, 25–33. [Google Scholar] [CrossRef]
- Maddock, I.; Harby, A.; Kemp, P.; Wood, P. Ecohydraulics: An Integrated Approach; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 1–6. [Google Scholar]
- Schwartz, J.S.; Neff, K.J.; Dworak, F.J.; Woockman, R.R. Restoring riffle-pool structure in an incised, straightened urban stream channel using an ecohydraulic modeling approach. Ecol. Eng. 2015, 78, 112–126. [Google Scholar] [CrossRef]
- Orth, D.J.; Maughan, O.E. Evaluation of incremental methodology for recommending instream flows for fishes. Trans. Am. Fish. Soc. 1982, 111, 413–445. [Google Scholar] [CrossRef]
- Bovee, K.D.; Lamb, B.L.; Bartholow, J.M.; Stalnaker, C.B.; Taylor, J.; Henroksen, J. Stream Habitat Analysis Using the Instream Flow Incremental Methodology; U.S. Geological Survey, Biological Resources Division Information and Technology Report: Fort Collins, CO, USA, March 1998.
- Vadas, R.L.; Orth, D.J. Formulation of habitat suitability models for stream fish guilds: Do the standard methods work? Trans. Am. Fish. Soc. 2001, 130, 217–235. [Google Scholar] [CrossRef]
- Persinger, J.W.; Orth, D.J.; Averett, A.W. Using habitat guilds to development habitat suitability criteria for warmwater stream fish assemblages. River Res. Appl. 2011, 27, 956–966. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Herricks, E.E. Use of prepositioned areal electrofishing devices with rod electrodes in small streams. N. Am. J. Fish. Manag. 2004, 24, 1330–1340. [Google Scholar] [CrossRef]
- Lanchester, J.; Downes, B.J. Linking the hydraulic world of individual organisms to ecological processes: Putting ecology into ecohydraulics. River Res. Appl. 2010, 26, 385–403. [Google Scholar] [CrossRef]
- Benjankar, R.; Tonina, D.; McKean, J. One-dimensional and two-dimensional hydrodynamic modeling derived flow properties: Impacts on aquatic habitat quality predictions. Earth Surface Process. Landf. 2015, 40, 340–356. [Google Scholar] [CrossRef]
- Tonina, D.; Jorde, K. Hydraulic modeling approaches for ecohydraulic studies: 3D. 2D, 1D and non-numerical models. In Ecohydraulics: An Integrated Approach; Maddock, I., Harby, A., Kemp, P., Wood, P., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 31–74. [Google Scholar]
- LeClerc, M.; Boudreault, A.; Bechara, J.A.; Corfa, G. Two-dimensional hydrodynamic modeling: A neglected tool in the instream flow incremental methodology. Trans. Am. Fish. Soc. 1995, 124, 645–662. [Google Scholar] [CrossRef]
- Booker, D.J.; Dunbar, M.J. Application of physical habitat simulation (PHABSIM) modelling to modified urban river channels. River Res. Appl. 2004, 20, 167–183. [Google Scholar] [CrossRef]
- Pasternack, G.B.; Wang, C.L.; Merz, J.E. Application of a 2D hydrodynamic model to design of reach-scale spawning gravel replenishment on the Mokelumme River, California. River Res. Appl. 2004, 20, 205–225. [Google Scholar] [CrossRef]
- Steffler, P.; Blackburn, J. River2D, Two-Dimensional Depth Averaged Model of River Hydrodynamics and Fish Habitat, Introduction to Depth Averaged Modeling and User’s Manual; University of Alberta: Edmonton, AB, Canada, 2002. [Google Scholar]
- Schwartz, J.S. Use of a 2D Hydrodynamic Model for Stream Restoration Design of High-flow Habitat in Low-gradient Midwest Streams. In Proceedings of the Symposium on Protection and Restoration of Urban and Rural Streams, Philadelphia, PA, USA, 23–25 June 2003; Clar, M., Carpenter, D., Gracie, J., Slate, L., Eds.; American Society of Civil Engineers: Reston, VA, USA, 2003; pp. 242–251. [Google Scholar]
- Parasiewicz, P. MesoHABSIM: A concept for application of instream flow models in river restoration. Fisheries 2001, 26, 6–13. [Google Scholar] [CrossRef]
- Parasiewicz, P. Using MesoHABSIM to develop reference habitat template and ecological management scenarios. River Res. Appl. 2007, 23, 924–932. [Google Scholar] [CrossRef]
- Lamouroux, N.; Capra, H.; Pouilly, M. Predicting habitat suitability for lotic fish: Linking statistical models with multivariate habitat use models. Regul. Rivers Res. Manag. 1998, 14, 1–11. [Google Scholar] [CrossRef]
- Bain, M.B.; Meixler, M.S. A target fish community to guide river restoration. River Res. Appl. 2008, 24, 453–458. [Google Scholar] [CrossRef]
- Mueller, M.; Pander, J.; Geist, J. The ecological value of stream restoration measures: An evaluation on ecosystem and target species scales. Ecol. Eng. 2014, 62, 129–139. [Google Scholar] [CrossRef]
- Lacey, R.W.J.; Millar, R.G. Reach scale hydraulic assessment of instream salmonid habitat restoration. J. Am. Water Resour. Assoc. 2004, 40, 1631–1644. [Google Scholar] [CrossRef]
- Parasiewicz, P. Applications of MesoHABSIM and target fish community approaches to restoration of the Quinebaug River, Connecticut and Massachusetts, USA. River Res. Appl. 2008, 24, 459–471. [Google Scholar] [CrossRef]
- Brown, R.A.; Pasternack, G.B. Comparison of methods for analyzing salmon habitat rehabilitation designs for regulated rivers. River Res. Appl. 2009, 25, 745–772. [Google Scholar] [CrossRef]
- Frimpong, E.A.; Angermeier, P.L. Trait-based approaches in the analysis of fish communities. Am. Fish. Soc. Symp. 2010, 73, 109–136. [Google Scholar]
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Belyea, L.R.; Lancaster, J. Assembly rules within a contingent ecology. Oikos 1999, 86, 402–416. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Nieira, K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef]
- Statzner, B.; Beche, L.A. Can biological traits resolve effects of multiple stressors on running water ecosystems? Freshw. Biol. 2010, 55, 80–119. [Google Scholar] [CrossRef]
- Keck, B.P.; Marion, Z.H.; Martin, D.J.; Kaufman, J.C.; Harden, C.P.; Schwartz, J.S.; Strange, R.J. Fish functional traits correlated with environmental traits in a temperate biodiversity hot spot. PLoS ONE 2014, 9, e93237. [Google Scholar] [CrossRef] [PubMed]
- Poff, N.L.; Allan, J.D. Functional organization of stream fish assemblages in relation to hydrological variability. Ecology 1995, 76, 606–627. [Google Scholar] [CrossRef]
- Kemp, J.L.; Harper, D.M.; Cross-Giuseppe, A. Use of “functional habitats” to link ecology with morphology and hydrology in river restoration. Aquat. Conserv. 1999, 9, 159–178. [Google Scholar] [CrossRef]
- Newson, M.D.; Newson, C.L. Geomorphology, ecology and river channel habitat: Mesoscale approaches to basin-scale challenges. Prog. Phys. Geogr. 2000, 24, 195–217. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities—The roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar]
- Lamouroux, N.; Olivier, J.; Persat, H.; Pouilly, M.; Souchon, Y.; Statzner, B. Predicting community characteristics from habitat conditions: Fluvial fish and hydraulics. Freshw. Biol. 1999, 42, 275–299. [Google Scholar] [CrossRef]
- Lamouroux, N.; Poff, N.L.; Angermeier, P.L. Intercontinental convergence of fish community traits along geomorphic and hydraulic gradients. Ecology 2002, 83, 1792–1807. [Google Scholar] [CrossRef]
- Bayley, P.B.; Li, H.W. Riverine fishes. In River Biota: Diversity and Dynamics; Petts, G., Calow, P., Eds.; Blackwell Science Ltd.: London, UK, 1996; pp. 92–122. [Google Scholar]
- Palmer, M.A.; Hakenkamp, C.C.; Nelson-Baker, K. Ecological heterogeneity in streams: Why variance matters. J. N. Am. Benthol. Soc. 1997, 16, 189–202. [Google Scholar] [CrossRef]
- Amoros, C. The concept of habitat diversity between and within ecosystems applied to river side-arm restoration. Environ. Manag. 2001, 28, 805–817. [Google Scholar] [CrossRef]
- Buffington, J.M.; Montgomery, D.R. Geomorphic classification of rivers. In Treatise on Geomorphology; Fluvial Geomorphology; Volume 9, Shroder, J., Wohl, E., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 730–767. [Google Scholar]
- Wolman, M.G.; Miller, J.P. Magnitude and frequency of forces in geomorphic processes. J. Geol. 1960, 68, 54–74. [Google Scholar] [CrossRef]
- Ackers, P.; Charlton, F.G. Dimensional analysis of alluvial channels with special reference to meander length. J. Hydraul. Res. 1976, 8, 287–316. [Google Scholar] [CrossRef]
- Richards, K.S. The morphology of riffle-pool sequences. Earth Surf. Process. Landf. 1976, 1, 71–88. [Google Scholar] [CrossRef]
- Rosgen, D.L. A classification of natural rivers. Catena 1994, 22, 169–199. [Google Scholar] [CrossRef]
- Wolman, M.G.; Leopold, L.B. River flood plains: Some observations on their formation. U.S. Geol. Surv. Prof. Pap. 1957, 282C, 87–109. [Google Scholar]
- Page, K.; Nanson, G. Concave-bank benches and associated floodplain formation. Earth Surf. Process. Landf. 1982, 7, 529–543. [Google Scholar] [CrossRef]
- Nanson, G.C.; Croke, J.C. A genetic classification of floodplains. Geomorphology 1992, 4, 459–486. [Google Scholar] [CrossRef]
- Dietrich, W.E. Mechanics of flow and sediment transport in river bends. In River Channel Environment and Process; Richards, K., Ed.; Blackwell Press: Basil, Russia, 1987; pp. 179–227. [Google Scholar]
- Hooke, J.M. River channel adjustment to meander cutoffs on the River Bolin and River Dane, northwest England. Geomorphology 1995, 14, 235–253. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M. Channel-reach morphology in mountain drainage basins. Geol. Soc. Am. Bull. 1997, 109, 596–611. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M.; Smith, R.D.; Schmidt, K.M.; Pess, G. Pool spacing in forest channels. Water Resour. Res. 1995, 31, 1097–1105. [Google Scholar] [CrossRef]
- Keller, E.A.; Florsheim, J.L. Velocity-reversal hypothesis: A model approach. Earth Surf. Process. Landf. 1993, 18, 733–740. [Google Scholar]
- Cao, Z.; Carling, P.; Oakley, R. Flow reversal over a natural pool-riffle sequence: A computational study. Earth Surf. Process. Landf. 2003, 28, 689–705. [Google Scholar] [CrossRef]
- Wilkinson, S.N.; Keller, R.J.; Rutherfund, I.D. Phase-shifts in shear stress as an explanation for the maintenance of pool-riffle sequences. Earth Surf. Process. Landf. 2004, 29, 737–753. [Google Scholar] [CrossRef]
- Smith, D.L.; Brannon, E.L. Influence of cover on mean column hydraulic characteristics in small pool-riffle morphology streams. River Res. Appl. 2007, 23, 125–139. [Google Scholar] [CrossRef]
- Colombini, M.; Seminara, G.; Tubino, M. Finite-amplitude alternate bars. J. Fluid Mech. 1987, 181, 213–232. [Google Scholar] [CrossRef]
- Whiting, P.J.; Dietrich, W.E. Convective accelerations and boundary shear stress over a channel bar. Water Resour. Res. 1991, 27, 783–796. [Google Scholar] [CrossRef]
- Lanzoni, S.; Tubino, M. Grain sorting and bar instability. J. Fluid Mech. 1999, 393, 149–174. [Google Scholar] [CrossRef]
- Frothingham, K.M.; Rhoads, B.L. Three-dimensional flow structure and channel change in an asymmetrical compound meander loop, Embarras River, Illinois. Earth Surf. Process. Landf. 2003, 28, 625–644. [Google Scholar] [CrossRef]
- Biron, P.M.; Carver, R.B.; Carre, D.M. Sediment transport and flow dynamics around a restored pool in a fish habitat rehabilitation project: Field and 3D numerical modeling experiments. River Res. Appl. 2012, 28, 926–939. [Google Scholar] [CrossRef]
- Dietrich, W.E.; Smith, J.D. Influence of the point bar on flow through curved channels. Water Resour. Res. 1983, 19, 1173–1192. [Google Scholar] [CrossRef]
- Nelson, J.M.; Bennett, J.P.; Wiele, S.M. Flow and sediment-transport modeling. In Tools in Fluvial Geomorphology; Kondolg, M.G., Piegay, H., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2003; pp. 539–576. [Google Scholar]
- Rhoads, B.L.; Kenworthy, S.T. Flow structure at an asymmetrical stream confluence. Geomorphology 1995, 11, 273–293. [Google Scholar] [CrossRef]
- Rodriguez, J.F.; Garcia, C.M.; Garcia, M.H. Three-dimensional flow in centered pool-riffle sequences. Water Resour. Res. 2013, 49, 202–215. [Google Scholar] [CrossRef]
- Booker, D.; Sear, D.; Payne, A. Modelling three-dimensional flow structures and patterns of boundary shear stress in a natural pool-riffle sequence. Earth Surf. Process. Landf. 2001, 26, 553–576. [Google Scholar] [CrossRef]
- MacWilliams, M.L.; Wheaton, J.M.; Pasternack, G.B.; Street, R.L.; Kitanidis, P.K. Flow convergence routing hypothesis for pool-riffle maintenance in alluvial rivers. Water Resour. Res. 2006, 42, 1–22. [Google Scholar] [CrossRef]
- Caamano, D.; Goodwin, P.; Buffington, J.M.; Liou, J.C.; Daley-Laursen, S. Unifying criterion for the velocity reversal hypothesis in gravel-bed rivers. J. Hydraul. Eng. 2009, 135, 66–70. [Google Scholar] [CrossRef]
- MacVicar, B.J.; Roy, A.G. Sediment mobility in a forced riffle-pool. Geomorphology 2011, 125, 445–456. [Google Scholar] [CrossRef]
- Caamano, D.; Goodwin, P.; Buffington, J.M. Flow structure through pool-riffle sequences and a conceptual model for their sustainability in gravel-bed rivers. River Res. Appl. 2012, 28, 377–389. [Google Scholar] [CrossRef]
- Milan, D.J. Sediment routing hypothesis for pool-riffle maintenance. Earth Surf. Process. Landf. 2013, 38, 1623–1641. [Google Scholar] [CrossRef]
- Pasternack, G.B.; Brown, R.A. Ecohydraulic design of riffle-pool relief and morphological unit geometry in support of regulated gravel-bed river rehabilitation. In Ecohydraulics: An Integrated Approach; Maddock, I., Harby, A., Kemp, P., Wood, P., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 337–355. [Google Scholar]
- Crowder, D.W.; Diplas, P. Vorticity and circulation: Spatial metrics for evaluating flow complexity in stream habitats. Can. J. Fish. Aquat. Sci. 2002, 59, 633–645. [Google Scholar] [CrossRef]
- Gurnell, A.M.; Sweet, R. The distribution of large woody debris accumulations and pools in relation to woodland stream management in a small, low-gradient stream. Earth Surf. Process. Landf. 1998, 23, 1101–1121. [Google Scholar] [CrossRef]
- Myers, T.; Swanson, F.J. Variability of pool characteristics with pool type and formative feature on small Great Basin rangeland streams. J. Hydrol. 1997, 201, 62–81. [Google Scholar] [CrossRef]
- Thompson, D.M.; Hoffman, K.S. Equilibrium pool dimensions and sediment-sorting patterns in coarse-grained, New England channels. Geomorphology 2001, 38, 301–316. [Google Scholar] [CrossRef]
- Buffington, J.M.; Lisle, T.E.; Woodsmith, R.D.; Hilton, S. Controls on the size and occurrence of pools in coarse-grained forest rivers. River Res. Appl. 2002, 18, 507–531. [Google Scholar] [CrossRef]
- Power, M.E.; Stout, R.J.; Cushing, C.E.; Harper, P.P.; Hauer, F.R.; Matthews, W.J.; Moyle, P.B.; Statzner, B.; Wais De Badgen, I.R. Biotic and abiotic controls in rivers and stream communities. J. N. Am. Benthol. Soc. 1988, 7, 456–479. [Google Scholar] [CrossRef]
- Horwitz, R.J. Temporal variability patterns and the distributional patterns of stream fishes. Ecol. Monogr. 1978, 48, 307–321. [Google Scholar] [CrossRef]
- Schlosser, I.J. Fish community structure and function along two habitat gradients in a headwater stream. Ecol. Monogr. 1982, 52, 395–414. [Google Scholar] [CrossRef]
- Grenouillet, G.; Pont, D.; Herisse, C. Within-basin fish assemblage structure: The relative influence of habitat versus stream spatial position on local species richness. Can. J. Fish. Aquat. Sci. 2004, 61, 93–102. [Google Scholar] [CrossRef]
- Li, R.Y.; Gelwick, F.P. The relationship of environmental factors to spatial and temporal variation of fish assemblages in a floodplain river in Texas, USA. Ecol. Freshw. Fish 2005, 14, 319–330. [Google Scholar] [CrossRef]
- Merigoux, S.; Doledec, S.; Statzner, B. Species traits in relation to habitat variability and state: Neotrophic juvenile fish in floodplain creeks. Freshw. Biol. 2001, 46, 1251–1267. [Google Scholar] [CrossRef]
- Taylor, C.M.; Warren, M.L., Jr. Dynamics in species composition of stream fish assemblages: Environmental variability and nested subsets. Ecology 2001, 82, 2320–2330. [Google Scholar] [CrossRef]
- Angermeier, P.L. Resource seasonality and fish diets in an Illinois stream. Environ. Biol. Fish. 1982, 7, 251–264. [Google Scholar] [CrossRef]
- Ingersoll, C.G.; Hlohowskyj, I.; Mundahl, N.D. Movements and densities of the darters Etheostoma Flabellare, E. Spectabile, and E. Nigrum During Spring Spawning. J. Freshw. Ecol. 1984, 2, 345–351. [Google Scholar] [CrossRef]
- Langhurst, R.W.; Schoenike, D.L. Seasonal migration of smallmouth bass in the Embarrass and Wolf Rivers, Wisconsin. N. Am. J. Fish. Manag. 1990, 10, 224–227. [Google Scholar] [CrossRef]
- Matthews, W.J. Spatial and temporal variation in fishes of riffle habitats: A comparison of analytical approaches for the Roanoke River. Am. Midl. Nat. 1990, 124, 31–45. [Google Scholar] [CrossRef]
- Vadas, R.L. Seasonal habitat use, species associations, and assemblage structure of forage fishes in Goose Creek, northern Virginia. I. macrohabitat patterns. Freshw. Ecol. 1991, 6, 403–417. [Google Scholar] [CrossRef]
- Dewey, M.R. Seasonal abundance, movement and diversity of fishes in an Ozark stream. Ark. Acad. Sci. Proc. 1981, 35, 33–39. [Google Scholar]
- Smith, P.W. A study of seasonal distribution of fishes in the Kaskaskia River ditch, a highly modified stream in eastern Illinois. Copeia 1963, 2, 251–259. [Google Scholar] [CrossRef]
- Smith, P.W. The Fishes of Illinois; University of Illinois Press: Urbana, IL, USA, 1979; pp. 1–314. [Google Scholar]
- Poff, N.L.; Ward, J.V. Implications of streamflow variability and predictability for lotic community structure: A regional analysis of streamflow patterns. Can. J. Fish. Aquat. Sci. 1989, 46, 1805–1818. [Google Scholar] [CrossRef]
- Kelsch, S.W. Lotic fish-community structure following transition from severe drought to high discharge. J. Freshw. Ecol. 1994, 9, 331–341. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Ross, S.T.; Baker, J.A. The response of fishes to periodic spring floods in a Southeastern stream. Am. Midl. Nat. 1983, 109, 1–14. [Google Scholar] [CrossRef]
- Kwak, T.J. Lateral movement and use of floodplain habitat by fishes of the Kankakee River, Illinois. Am. Midl. Nat. 1988, 120, 241–249. [Google Scholar] [CrossRef]
- Lancaster, J.; Hildrew, A.G. Characterizing in-stream flow refugia. Can. J. Fish. Aquat. Sci. 1993, 50, 1663–1675. [Google Scholar] [CrossRef]
- Niemi, G.J.; DeVore, P.; Detenbeck, N.; Taylor, D.; Lima, A.; Pastor, J.; Yount, J.D.; Naiman, R.J. Overview of case studies on recovery aquatic ecosystems from disturbance. Environ. Manag. 1990, 14, 571–587. [Google Scholar] [CrossRef]
- Reice, S.R.; Wissmar, R.C.; Naiman, R.J. Disturbance regimes, resilience, and recovery of animal communities and habitats in lotic ecosystems. Environ. Manag. 1990, 14, 647–659. [Google Scholar] [CrossRef]
- Matthews, W.J. Fish faunal structure in an Ozark stream: Stability, persistence, and a catastrophic flood. Copeia 1986, 2, 388–397. [Google Scholar] [CrossRef]
- Pearsons, T.N.; Li, H.W.; Lamberti, G.A. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Am. Fish. Soc. 1992, 121, 427–436. [Google Scholar] [CrossRef]
- Lobón-Cerviá, J. Response of a stream fish assemblage to a severe spate in northern Spain. Trans. Am. Fish. Soc. 1996, 125, 913–919. [Google Scholar] [CrossRef]
- Fausch, K.D.; Bramblett, R.G. Disturbance and fish communities in intermittent tributaries of a Western Great Plains river. Copeia 1991, 3, 659–673. [Google Scholar] [CrossRef]
- Lohr, S.C.; Fausch, K.D. Multiscale analysis of natural variability in stream fish assemblages of a western Great Plains watershed. Copeia 1997, 4, 706–724. [Google Scholar] [CrossRef]
- Larimore, R.W.; Garrels, D.D. Assessing habitats used by warmwater stream fishes. Fisheries 1985, 10, 10–16. [Google Scholar] [CrossRef]
- Schlosser, I.J. The role of predation in age- and size-related habitat use by stream fishes. Ecology 1987, 68, 651–659. [Google Scholar] [CrossRef]
- Gelwick, F.P. Longitudinal and temporal comparisons of riffle and fish pool assemblages in a northeastern Oklahoma Ozark stream. Copeia 1990, 4, 1072–1082. [Google Scholar] [CrossRef]
- Lobb, M.D.; Orth, D.J. Habitat use by an assemblage of fish in a large warmwater stream. Trans. Am. Fish. Soc. 1991, 120, 65–78. [Google Scholar] [CrossRef]
- Aadland, L.P. Stream habitat types: Their assemblages and relationship to flow. N. Am. J. Fish. Manag. 1993, 13, 790–806. [Google Scholar] [CrossRef]
- Matthews, W.J.; Harvey, B.C.; Power, M.E. Spatial and temporal patterns in fish assemblages of individual pools in a Midwestern stream. Environ. Biol. Fish. 1994, 39, 381–397. [Google Scholar] [CrossRef]
- Lonzarich, D.G.; Lonzarich, M.R.; Warren, M.L., Jr. Effects of riffle length on the short-term movement of fishes among stream pools. Can. J. Fish. Aquat. Sci. 2000, 57, 1508–1514. [Google Scholar] [CrossRef]
- Bisson, P.A.; Sullivan, K.; Nielsen, J.L. Channel hydraulics, habitat use, and body form of juvenile coho salmon, steelhead, and cutthroat trout in streams. Trans. Am. Fish. Soc. 1988, 117, 262–273. [Google Scholar] [CrossRef]
- Schlosser, I.J. Predation rates and the behavioral response of adult brassy minnows (Hybognathus hankinsoni) to creek chub and smallmouth bass predators. Copeia 1988, 3, 691–697. [Google Scholar] [CrossRef]
- Hearn, W.E. Interspecific competition and habitat segregation among stream-dwelling trout and salmon: A review. Fisheries 1987, 12, 24–31. [Google Scholar] [CrossRef]
- Frissell, C.A.; Lonzarich, D.G. Habitat use and competition among stream fishes. In Methods in Stream Ecology; Hauer, R.F., Lambert, G.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 493–510. [Google Scholar]
- Schlosser, I.J. Flow regime, juvenile abundance, and the assemblage structure on stream fishes. Ecology 1985, 66, 1484–1490. [Google Scholar] [CrossRef]
- Moore, K.M.S.; Gregory, S.V. Summer habitat utilization and ecology of cutthroat trout fry (Salmo clarki) in Cascade Mountain streams. Can. J. Fish. Aquat. Sci. 1988, 45, 1921–1930. [Google Scholar] [CrossRef]
- Payne, B.A.; Lapointe, M.F. Channel morphology and lateral stability: Effects on distribution of spawning and rearing habitat for Atlantic salmon in a wandering cobble-bed river. Can. J. Fish. Aquat. Sci. 1997, 54, 2627–2636. [Google Scholar] [CrossRef]
- Bain, M.B.; Finn, J.T.; Brooke, H.E. Streamflow regulations and fish community structure. Ecology 1988, 69, 382–392. [Google Scholar] [CrossRef]
- Rakocinski, C. Population structure of stream-dwelling darters: Correspondence with habitat structure. Environ. Biol. Fish. 1988, 23, 215–224. [Google Scholar] [CrossRef]
- Hayes, J.W.; Jowett, I.G. Microhabitat models of large drift-feeding brown trout in three New Zealand rivers. N. Am. J. Fish. Manag. 1994, 14, 710–725. [Google Scholar] [CrossRef]
- McMahon, T.E.; Hartman, G.F. Influence of cover complexity and current velocity on winter habitat use by juvenile coho salmon (Oncorhynchus kisutch). Can. J. Fish. Aquat. Sci. 1989, 46, 1551–1557. [Google Scholar] [CrossRef]
- Blanck, A.; Tedesco, P.A.; Lamouroux, N. Relationships between life-history strategies of European freshwater fish species and their habitat preferences. Freshw. Biol. 2007, 52, 843–859. [Google Scholar] [CrossRef]
- Jowett, I.G.; Duncan, M.J. Flow variability in New Zealand Rivers and its relationship to instream habitat and biota. N. Z. J. Mar. Freshw. Res. 1990, 24, 305–318. [Google Scholar] [CrossRef]
- Yu, S.; Peters, E.J. Use of Froude number to determine habitat selection by fish. Rivers 1997, 6, 10–18. [Google Scholar]
- Tyler, J.A.; Clapp, D.P. Perceptual constraints on stream fish habitat selection: Effects of food availability and water velocity. Ecol. Freshw. Fish 1995, 4, 9–16. [Google Scholar] [CrossRef]
- Litvak, M.K.; Hansell, R.I.C. Investigation of food habit and niche relationships in a cyprinid community. Can. J. Zool. 1990, 68, 1873–1897. [Google Scholar] [CrossRef]
- Schlosser, I.J.; Ebel, K.K. Effects of flow regime and cyprind predation on a headwater stream. Ecol. Monogr. 1989, 59, 41–57. [Google Scholar] [CrossRef]
- Curry, K.D.; Spacies, A. Differential use of stream habitat by spawning Catostomids. Am. Midl. Nat. 1983, 111, 267–279. [Google Scholar] [CrossRef]
- Emery, J.C.; Gurnell, A.M.; Clifford, N.J.; Petts, G.E.; Morrissey, I.P.; Soar, P.J. Classifying the hydraulic performance of riffle-pool bedforms for habitat assessment and river rehabilitation design. River Res. Appl. 2003, 19, 553–549. [Google Scholar] [CrossRef]
- Clifford, N.J.; Soar, P.J.; Harmar, O.P.; Gurnell, A.M.; Petts, G.E.; Emery, J.C. Assessment of hydrodynamic simulation results for eco-hydraulic and eco-hydrological applications: A spatial semivariance approach. Hydrol. Process. 2005, 19, 3631–3648. [Google Scholar] [CrossRef]
- Nester, J.M.; Stewardson, M.J.; Gilvear, D.; Webb, J.A.; Smith, D.L. Ecohydraulics exemplifies the emerging “paradigm of the interdisciplines”. J. Ecohydraul. 2016, in press. [Google Scholar]
- Holm, E. Environmental restraints and life strategies: A habitat templat matrix. Oecologia 1988, 75, 141–145. [Google Scholar] [CrossRef]
- Matthews, W.J. Patterns in Freshwater Ecology; Kluwer Academic Press: Norwell, MA, USA, 1988; p. 616. [Google Scholar]
- Persat, H.; Olivier, J.; Pont, D. Theoretical habitat templats, species traits, and species richness: Fish in the Upper Rhone River and its floodplain. Freshw. Biol. 1994, 31, 439–454. [Google Scholar] [CrossRef]
- Doledec, S.; Statzner, B.; Bournard, M. Species traits for future biomonitoring across ecoregions: Patterns along a human-impacted river. Freshw. Biol. 1999, 42, 737–758. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Meador, M.R. Comparison of fish species traits from small streams to large rivers. Trans. Am. Fish. Soc. 2004, 133, 971–983. [Google Scholar] [CrossRef]
- Lamouroux, N.; Cattaneo, F. Fish assemblages and stream hydraulics consistent relations across spatial scales and regions. River Res. Appl. 2006, 22, 727–737. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Meador, M.R. Multilevel assessment of fish species traits to evaluate habitat degradation in streams of the upper Midwest. N. Am. J. Fish. Manag. 2005, 25, 180–194. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Building community ecology from functional traits. Trends Ecol. Evolut. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D.; Dayan, T. The guild concept and the structure of ecological communities. Ann. Rev. Ecol. Syst. 1991, 22, 115–143. [Google Scholar] [CrossRef]
- Austen, D.J.; Bayley, P.B.; Menzel, B.W. Importance of the guild concept to fisheries research and management. Fisheries 1994, 19, 12–20. [Google Scholar] [CrossRef]
- Welcomme, R.L.; Winemiller, K.O.; Cowx, I.G. Ecological guilds as a tool for environmental impact assessment in rivers. River Res. Appl. 2006, 22, 377–396. [Google Scholar] [CrossRef]
- Karr, J.R. Biological integrity: A long-neglected aspect of water resource management. Ecol. Appl. 1991, 1, 165–222. [Google Scholar] [CrossRef] [PubMed]
- Karr, J.R.; Yant, P.R.; Fausch, K.D.; Schlosser, I.J. Spatial and temporal variability of the index of biological integrity in three Midwestern streams. Trans. Am. Fish. Soc. 1987, 116, 1–11. [Google Scholar] [CrossRef]
- Karr, J.R. Defining and assessing ecological integrity: Beyond water quality. In Environmental Toxicology and Chemistry; Ward, C.H., Ed.; Pergamon Press: Tarrytown, NY, USA, 1993; pp. 1521–1531. [Google Scholar]
- Hughes, R.M.; Kaufman, P.R.; Herlihy, A.T.; Kincald, T.M.; Reynolds, L.; Larsen, D.P. A process for developing and evaluating indices of fish assemblage integrity. Can. J. Fish. Aquat. Sci. 1998, 55, 1618–1631. [Google Scholar] [CrossRef]
- Simon, T.P. Assessing the Sustainability and Biological Integrity of Water Resources Using Fish Communities; CRC Press, LLC.: Boca Raton, FL, USA, 1999; p. 671. [Google Scholar]
- Rheinhardt, R.D.; Rheinhardt, M.C.; Brinson, M.M.; Faser, K.E. Application of reference data for assessing and restoring headwater ecosystems. Restor. Ecol. 1999, 7, 241–251. [Google Scholar] [CrossRef]
- Berkman, H.E.; Rabeni, C.F. Effects of siltation on stream communities. Environ. Biol. Fish. 1987, 18, 285–294. [Google Scholar] [CrossRef]
- Rashleigh, B. Relation of environmental characteristics to fish assemblages in the Upper French Broad River Basin, North Carolina. Environ. Monit. Assess. 2004, 93, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.H.; Freeman, M.C.; Freeman, B.J.; Wenger, S.J.; Ensign, W.E. Investigating hydrologic alternation as a mechanism of fish assemblage shifts in urbanizing streams. J. N. Am. Benthol. Soc. 2005, 24, 656–678. [Google Scholar] [CrossRef]
- Brown, L.R.; Gregory, T.R.; May, J.T. Relation of urbanization to stream fish assemblages and species traits in nine metropolitan areas in the United States. Urban Ecosyst. 2009, 12, 391–416. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Simon, A.; Klimetz, L. Use of fish functional traits to associate in-stream suspended sediment transport metrics with biological impairment. Environ. Monit. Assess. 2011, 179, 347–369. [Google Scholar] [CrossRef] [PubMed]
- Doledec, S.; Chessel, D.; ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the species traits-environmental relationships: Thefourth corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Frimpong, E.A.; Angermeier, P.L. Fish traits: A database of ecological and life-history traits of freshwater fishes of the United States. Fisheries 2009, 34, 487–495. [Google Scholar] [CrossRef]
- Padmore, C.L. The role of physical biotopes in determining the conservation status and flow requirements of British rivers. Aquat. Ecosyst. Health Manag. 1998, 1, 25–35. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M. Channel processes, classification, and response. In River Ecology and Management; Naiman, R., Bilby, R., Eds.; Springer: New York, NY, USA, 1998; pp. 13–42. [Google Scholar]
- Kocik, J.F.; Ferreri, C.P. Juvenile production variation in salmonids: Population dynamics, habitat, and the role of spatial relationships. Can. J. Fish. Aquat. Sci. 1998, 55, 191–200. [Google Scholar] [CrossRef]
- Newson, M.D.; Harper, D.M.; Padmore, C.L.; Kemp, J.K.; Vogel, B. A cost-effective approach for linking habitats, flow types and species requirements. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 431–446. [Google Scholar] [CrossRef]
- Giberson, D.J.; Caissie, D. Stream habitat hydraulics: Interannual variability in three reaches of the Catamaran Brook, New Brunswick. Can. J. Fish. Aquat. Sci. 1998, 55, 485–494. [Google Scholar] [CrossRef]
- Wadeson, R.A.; Rowntree, K.M. Application of the hydraulic biotope concept to the classification of instream habitats. Aquat. Ecosyst. Health Manag. 1998, 1, 143–157. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Meador, M.R.; Ruhl, K.E. Relative influence of streamflows in assessing temporal variability in stream habitat. J. Am. Water Resour. Assoc. 2007, 43, 642–650. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Herricks, E.E. Fish use of stage-specific fluvial habitats as refuge patches during a flood in a low-gradient Illinois stream. Can. J. Fish. Aquat. Sci. 2005, 62, 1540–1552. [Google Scholar] [CrossRef]
- Lawless, M.; Robert, A. Three-dimensional flow structure around small-scale bedforms in a simulated gravel-bed environment. Earth Surf. Process. Landf. 2001, 26, 507–522. [Google Scholar] [CrossRef]
- Lawless, M.; Robert, A. Scales of boundary resistance in course-grained channels: Turbulent velocity profiles and implications. Geomorphology 2001, 39, 221–238. [Google Scholar] [CrossRef]
- Noss, C.; Salzmann, T.; Storchenegger, I. Turbulent and advective momentum fluxes in streams. Water Resour. Res. 2010, 46, 1–13. [Google Scholar] [CrossRef]
- Yalin, M.S. River Mechanics; Pergamon Press: New York, NY, USA, 1992; p. 219. [Google Scholar]
- Buffin-Belanger, T.; Roy, A.G.; Kirkbride, A.D. On large-scale flow structures in a gravel-bed river. Geomorphology 2000, 32, 417–435. [Google Scholar] [CrossRef]
- Schvidchenko, A.; Pender, G. Macroturbulent structure of open-channel flow over gravel beds. Water Resour. Res. 2001, 37, 709–719. [Google Scholar] [CrossRef]
- Carnie, R.; Tonina, D.; McKean, J.A.; Isaak, D.J. Habitat connectivity as a metric for aquatic microhabitat quality: Application to Chinook salmon spawning habitat. Ecohydrology 2016, 9, 982–994. [Google Scholar] [CrossRef]
- Waddle, T. Field evaluation of a two-dimensional hydrodynamic model near boulders for habitat calculations. River Res. Appl. 2009, 26, 730–741. [Google Scholar] [CrossRef]
- Rhoads, B.L.; Schwartz, J.S.; Porter, S. Stream geomorphology, bank vegetation and three-dimensional habitat hydraulics for fish in midwestern agricultural streams. Water Resour. Res. 2003, 39, 1218–1230. [Google Scholar] [CrossRef]
- Schmidt, J.C.; Rubin, D.M.; Ikeda, H. Flume simulation of recirculating flow and sedimentation. Water Resour. Res. 1993, 29, 2925–2939. [Google Scholar] [CrossRef]
- Tamburrino, A.; Gulliver, J.S. Large flow structures in turbulent open channel flow. J. Hydraul. Res. 1999, 37, 363–380. [Google Scholar] [CrossRef]
- Crowder, D.W.; Diplas, P. Evaluating spatially explicit metrics of stream energy gradients using hydrodynamic model simulations. Can. J. Fish. Aquat. Sci. 2000, 57, 1497–1507. [Google Scholar] [CrossRef]
- Thompson, D.M.; Wohl, E.E.; Jarrett, R.D. Velocity reversals and sediment sorting in pools and riffles controlled by channel constrictions. Geomorphology 1999, 27, 229–241. [Google Scholar] [CrossRef]
- Thompson, A. Secondary flows and the pool-riffle unit: A case study of the processes of meander development. Earth Surf. Process. Landf. 1986, 11, 631–641. [Google Scholar] [CrossRef]
- Geldof, H.J.; deVriend, H.J. Distribution of main flow velocity in alternating river bends. Spec. Publ. Int. Assoc. Sedimentol. 1983, 6, 85–95. [Google Scholar]
- Thompson, D.M.; Nelson, J.M.; Wohl, E.E. Interactions between pool geometry and hydraulics. Water Resour. Res. 1998, 34, 3673–3681. [Google Scholar] [CrossRef]
- Howard, A.D. Modeling channel migration and floodplain sedimentation in meandering streams. In Lowland Floodplain Rivers: Geomorphological Perspectives; Carling, P.A., Petts, G.E., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 1992; pp. 1–41. [Google Scholar]
- Nicholas, A.P.; Walling, D.E. Modeling flood hydraulics and overbank deposition on river floodplains. Earth Surf. Process. Landf. 1998, 22, 59–77. [Google Scholar] [CrossRef]
- Tsujimoto, T. Fluvial processes in streams with vegetation. J. Hydraul. Res. 1999, 37, 789–803. [Google Scholar] [CrossRef]
- Poulsen, J.B.; Hansen, F.; Ovesen, N.B.; Larsen, S.E.; Kronvang, B. Linking floodplain hydraulics and sedimentation patterns along a restored river channel: River Odense, Denmark. Ecol. Eng. 2014, 66, 120–128. [Google Scholar] [CrossRef]
- Needelman, B.A.; Kleinman, P.J.A.; Stock, J.S.; Allen, A.L. Drainage water management for water quality protection. J. Soil Water Conserv. 2010, 65, 131–136. [Google Scholar]
- D'Ambrosio, J.L.; Williams, L.R.; Witter, J.D.; Ward, A. Effects of geomorphology, habitat, and spatial location on fish assemblages in a watershed in Ohio, USA. Environ. Monit. Assess. 2009, 148, 325–341. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, J.L.; Ward, A.; Witter, J.D. Evaluating geomorphic change in constructed two-stage ditches. J. Am. Water Resour. Assoc. 2015, 51, 910–922. [Google Scholar] [CrossRef]
- Imhof, J.G.; Fitzgibbon, J.; Annable, W.K. A hierarchical evaluation system for characterizing watershed ecosystems for fish habitat. Can. J. Fish. Aquat. Sci. 1996, 53, 312–326. [Google Scholar]
- Wohl, E.; Angermeier, P.L.; Bledsoe, B.; Kondof, G.M.; MacDonnell, L.; Merritt, D.M.; Palmer, M.A.; Poff, N.L.; Tarboton, D. River restoration. Water Resour. Res. 2005, 41, W10301. [Google Scholar] [CrossRef]
- Nickelson, T.E.; Solazzi, M.F.; Johnson, S.L.; Rogers, J.D. Effectiveness of selected stream improvement techniques to create suitable summer and winter rearing habitat for juvenile coho salmon (Oncorhynchus kisutch) in Oregon coastal streams. Can. J. Fish. Aquat. Sci. 1992, 49, 790–794. [Google Scholar] [CrossRef]
- Zale, A.V.; Parrish, D.L.; Sutton, T.M. Fisheries Techniques, 3rd ed.; American Fisheries Society: Bethesda, MD, USA, 2013; p. 1009. [Google Scholar]
- Chessman, B.C.; Royal, M.J. Bioassessment without reference sites: Use of environmental filters to predict natural assemblages of river macroinvertebrates. J. N. Am. Benthol. Soc. 2004, 23, 599–615. [Google Scholar] [CrossRef]
- Stoddard, J.L.; Larsen, D.P.; Hawkins, C.P.; Johnson, R.K.; Norris, R.H. Setting expectations for the ecological condition of streams: The concept of reference condition. Ecol. Appl. 2006, 16, 1267–1276. [Google Scholar] [CrossRef]
- Herring, D.; Borja, A.; Cartensen, J.; Carvalho, L.; Elliott, M.; Fled, C.K.; Heiskanen, A.-S.; Johnson, R.K.; Moe, J.; Pont, D.; et al. The European Water Framework Directive at the age of 10: A critical review of the achievements with recommendations for the future. Sci. Total Environ. 2010, 408, 4007–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stream Flow Condition | Season | |
|---|---|---|
| Summer/Fall | Winter | |
| Low and Moderate Base Flows | Feeding Resting Escape from Predation Spawning | Temperature Refuge |
| High Flows—Floods | Flow Refuge | Flow Refuge |
| Formative Geomorphic Process: Erosion | |||
| Mesohabitat Unit | Geomorphic Characteristics | Hydraulic/Flow Depth Characteristics | Ecological Trait/Strategies |
| Pool Front | Entrance slope to a pool; downward-directed bed slope oriented with flow. | Flow acceleration, and strong outward velocities in meanders; relatively high turbulence [147,154,159]. Moderately deep (0.25–0.35 m). | Feeding by insectivores |
| Pool Mid | Topographic low along streambed; level bed. | Transition from flow acceleration to deceleration flow, and strong secondary circulation; submergence of high-velocity core; moderate turbulence [136,147,150,154,159]. Deep (0.4–1.2 m). | Feeding by piscivores and topminnows. Broadcast spawners. |
| Pool Rear | Exit slope to a pool; upward-directed bed slope oriented with flow. | Flow deceleration, and diminishing secondary circulation; relatively low turbulence [147,154,159]. Moderately deep (0.25 m). | Gravel nesting spawners. |
| Local Scour Pool | Small area of topographic low in bed; length smaller than channel width. | Local flow acceleration due to deflection and constriction of flow field [63,261,263,264,265]. Moderately deep (0.3 m). | Feeding by omnivores. |
| Formative Geomorphic Process: Deposition | |||
| Mesohabitat Unit | Geomorphic Characteristics | Hydraulic/Flow Depth Characteristics | Ecological Trait/Strategies |
| Glide | Intermediate bed topographic elevation; level and uniform bed. | Uniform downstream velocity vectors; minimal secondary circulation [63,212,246,251]. Moderately Shallow (0.25 m). | Feeding by omnivores, herbivores. Spawning by nest builders, other |
| Riffle without Raceway | Topographic intermediate to high along streambed; lateral bed morphology diversity. | Downstream velocities accelerate from increasing bed slope, weak surface-divergent secondary circulation; relatively moderate turbulence [63,136,150]. Moderately shallow (0.20 m). | Feeding by generalist and benthic insectivores. |
| Riffle with Raceway | Topographic high along streambed; sinuous flow path through alluvium during low flow; diverse bed morphology with small depressions. | Downstream velocities accelerate from increasing bed slope, weak surface-divergent secondary circulation relatively moderate turbulence [63,136,147]. Very shallow with deep “pockets“ (0.1–0.3 m). | Feeding by omnivores and herbivore. Spawners by egg adherence to gravel; darter resting |
| Submerged Bar | Lateral topographic high adjacent to pool, and extending into riffle structure. | Low velocities due to shoaling and lateral deflection of flow by the point bar; flow separation adjacent to or in lee of the point bar [136,147,149]. Shallow (0.15 m). | Evasion from predation by cyprinid young-of-the-year |
| Lateral Area in Lee of an Obstruction | Intermediate topographic elevation laterally positioned behind instream or bank structural element; area in lee of obstruction. | Separated, stagnant water or recirculating flow in lee of obstacle [251,261,263]. Shallow (0.15 m). | Evasion—escape from predators; resting; spawners by egg adherence to vegetation |
| Habitat Unit | Geomorphological Characteristics | Topographic Elevation | Hydraulic Characteristics |
|---|---|---|---|
| Hydrodynamic Condition: High-velocity corridor within the main channel | |||
| Channel Thalweg | Path of maximum depth and velocity within the channel; often shifts laterally across channel in conjunction with pool-riffle-bar morphology [71] | Lowest bed elevation in active channel [71,156] | Convergent-divergent flow pattern associated with pool-riffle-bar morphology; helical flow in pools; strong downstream velocities; high shear stress in pools [71,136,147] |
| Hydrodynamic Condition: Low-velocity areas along active channel margins | |||
| Deflection Eddy | Strong topographic deflection of flow laterally by elevated point-bar head, especially in sharp bends, resulting in an elongated zone of flow separation along inner bank [147,263]. | Intermediate bed elevation at margin of active channel along interface of point-bar face and platform; inundated with flow. | Separated, recirculating flow [136,147,150] |
| Expansion Eddy | Abrupt expansions of channel width at local re-entrants associated with remnant channels (floodplain sloughs) [137] or early stages of concave bank bench development in meandering channels [134], local bank ledge failures or other large structures that create local lateral channel constrictions [263,264]. | Local topographic low along channel bank, producing intermediate bed elevation within active channel; inundated above baseflow [270,273]. | Separated, recirculating flow within expansion zone [136,147,150]. |
| Local Hydraulic Dead Zone at Margins | Channel margins adjacent to thalweg in straight and meandering channels with high hydraulic roughness consisting of local bank irregularities and dense vegetation. | Vertical areas along bank at margins of active channel; wetted inundation slightly above baseflow [262,272]. | Stagnant or low-velocity areas produced by high hydraulic roughness [261,262,265]. |
| Hydrodynamic Condition: Low-velocity areas at channel-floodplain transition | |||
| Remnant Point-bar Platforms, Vegetated | Grassy, vegetated zones on surface of point-bar platform along inner (convex) bank of meander bends [71]. | Elevated surface above active channel; inundated at bankfull or overbank flows [273]. | Low velocities or recirculating flow from high hydraulic roughness during bankflow flow [264]. |
| Concave-bank Bench | Crescent-shaped depositional area at location corresponding to upstream limb of concave bank in the next bend downstream [134] bench becomes vegetated with grass. | Elevated surface about active channel; flood inundation only during bankfull or overbank flows [134,135]. | Separated, recirculating flow resulting from enlargement of expansion eddy during bankflow flow [134,270]. |
| Remnant channels; floodplain slough mouth | Remnant channels lateral to active channel; gradually slope towards at re-entrants locations at the mouth of floodplain sloughs [137,270,271]. | Elevated surface about active channel; flood inundation only during bankfull or overbank flows [135,137]. | Separated, recirculating flow resulting from enlargement of expansion eddy during bankflow flow [270]. |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwartz, J.S. Use of Ecohydraulic-Based Mesohabitat Classification and Fish Species Traits for Stream Restoration Design. Water 2016, 8, 520. https://doi.org/10.3390/w8110520
Schwartz JS. Use of Ecohydraulic-Based Mesohabitat Classification and Fish Species Traits for Stream Restoration Design. Water. 2016; 8(11):520. https://doi.org/10.3390/w8110520
Chicago/Turabian StyleSchwartz, John S. 2016. "Use of Ecohydraulic-Based Mesohabitat Classification and Fish Species Traits for Stream Restoration Design" Water 8, no. 11: 520. https://doi.org/10.3390/w8110520
APA StyleSchwartz, J. S. (2016). Use of Ecohydraulic-Based Mesohabitat Classification and Fish Species Traits for Stream Restoration Design. Water, 8(11), 520. https://doi.org/10.3390/w8110520