
Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis

 ,
, 

Abstract
:1. Introduction
1.1. Stream Processes Driving Nutrient Cycling
1.2. Stream Impairment in Human Dominated Watersheds
2. Review of Empirical Nutrient Studies
2.1. Selection Criteria and Typology Development
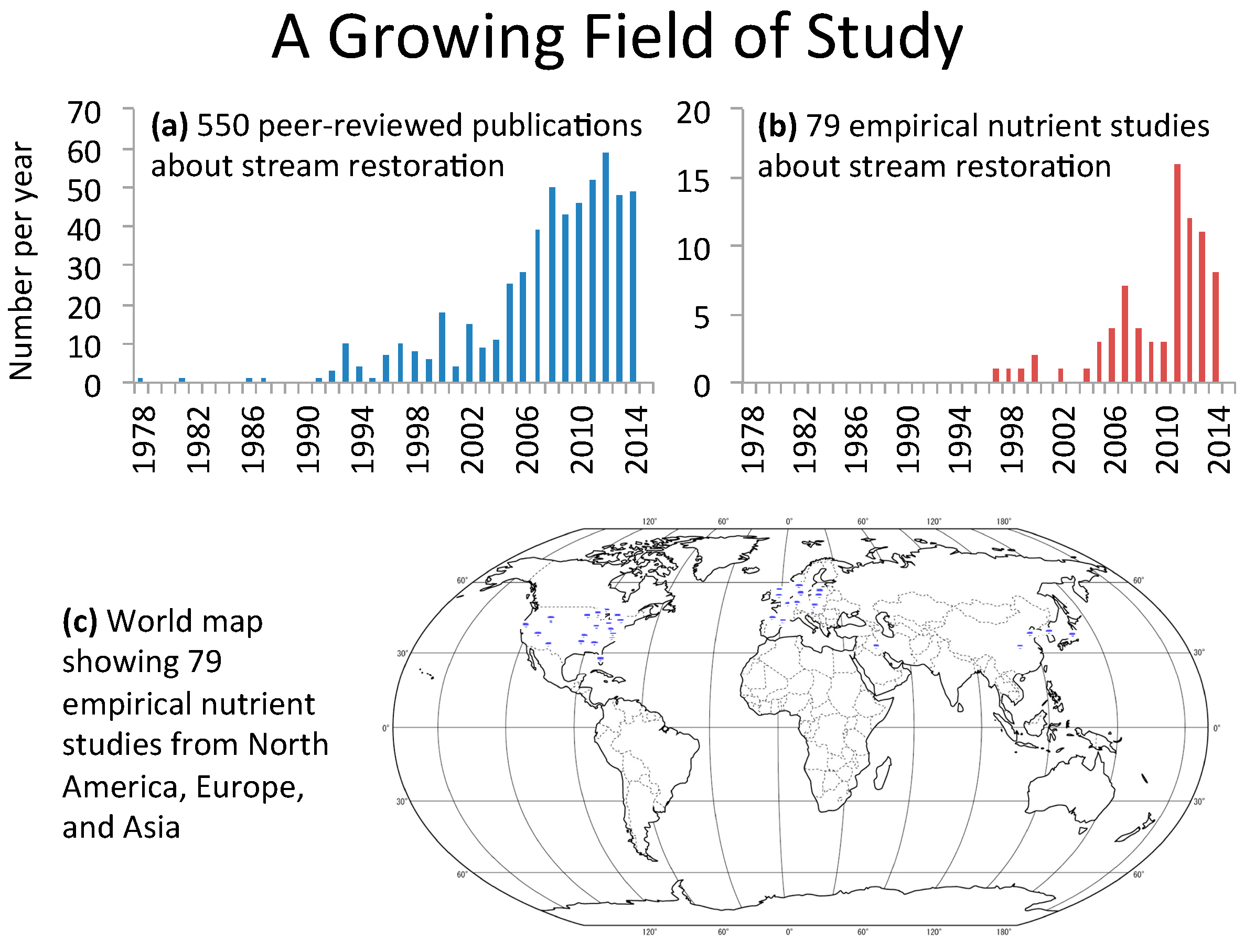
2.2. Growth in Stream Restoration Studies over Time
2.3. Restoration Typologies That Increase Hydrologic Connectivity
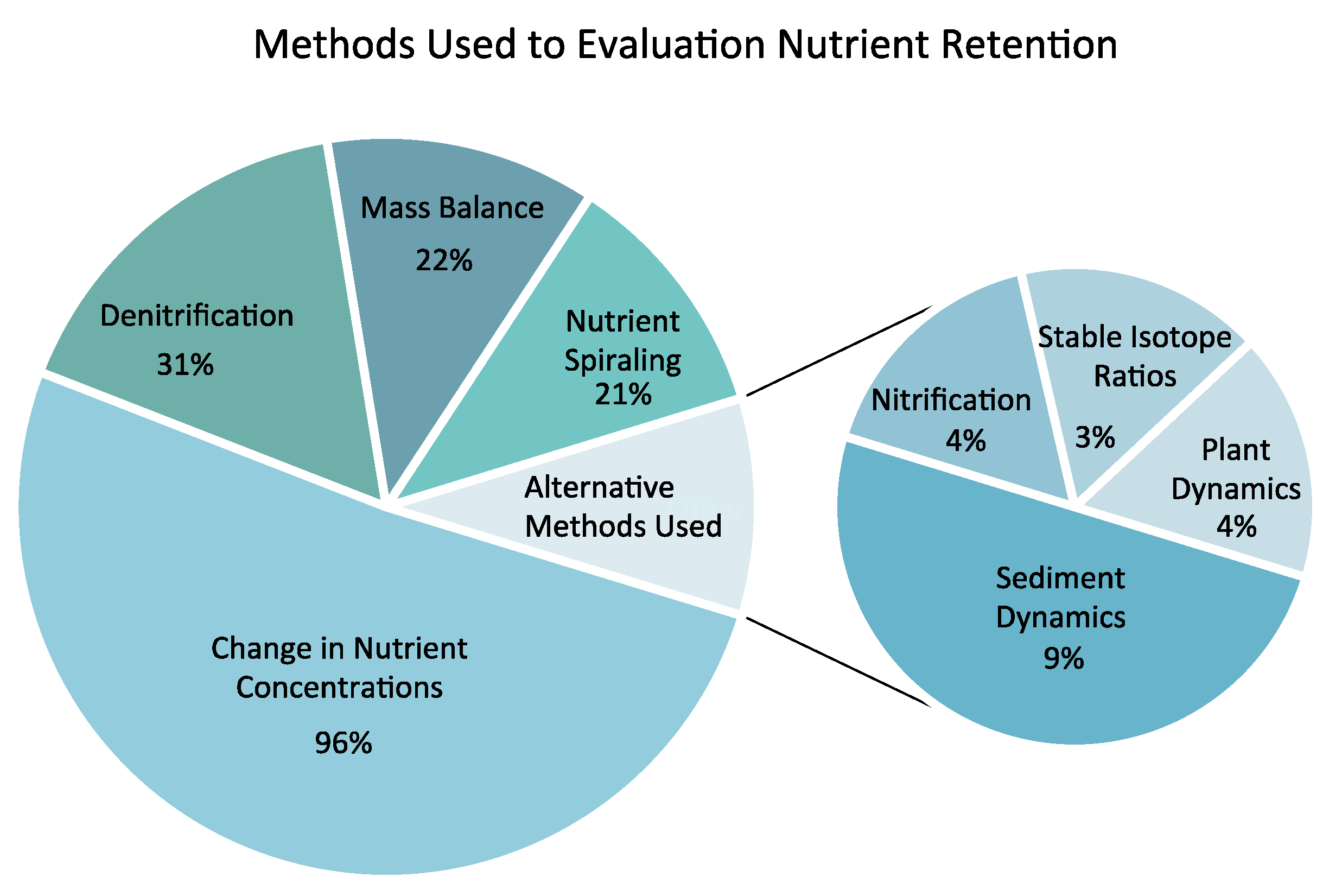
2.4. Comparison of Methods Used for Evaluating Stream Restoration Effectiveness
3. Nutrient Spiraling Meta-Data Analysis
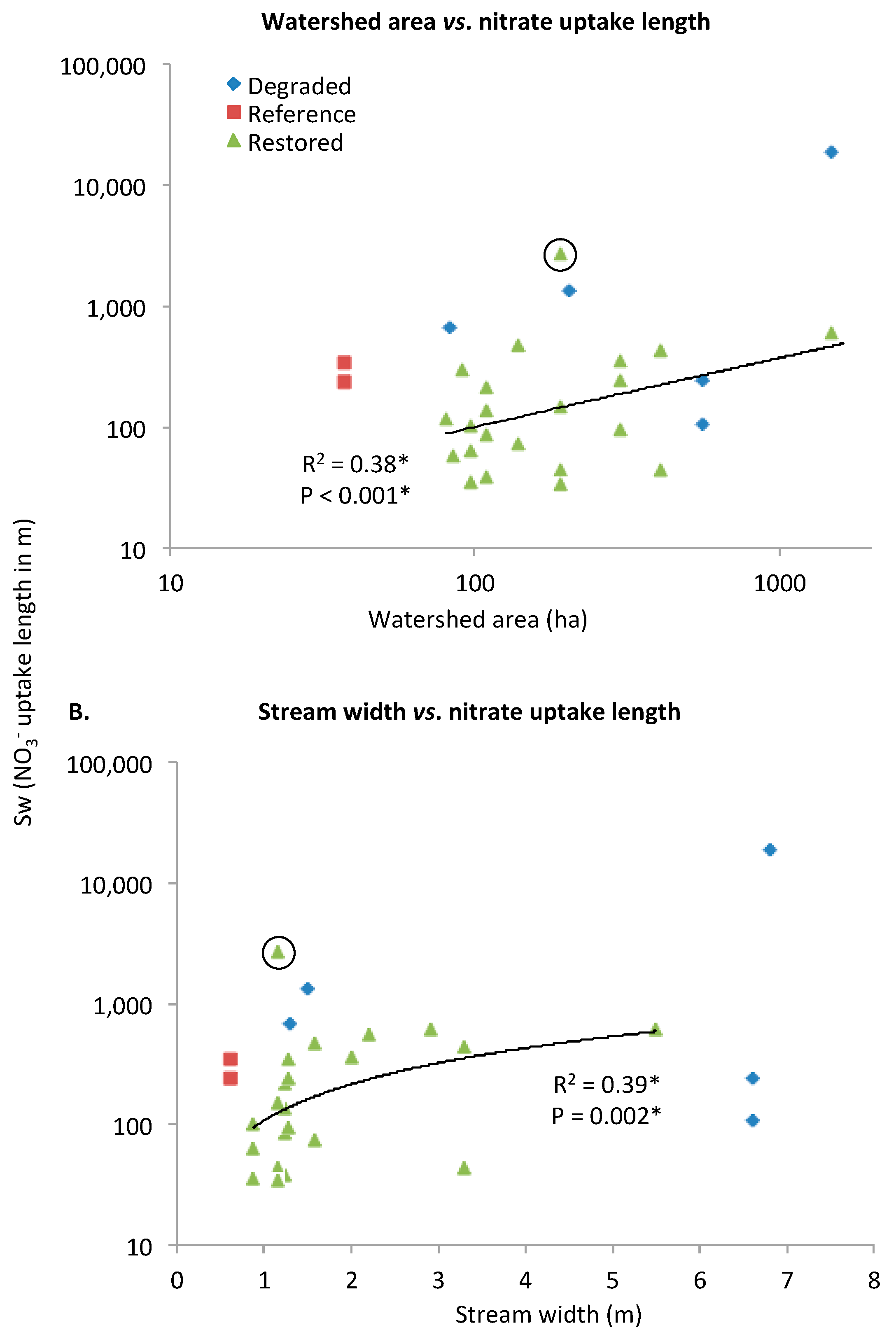
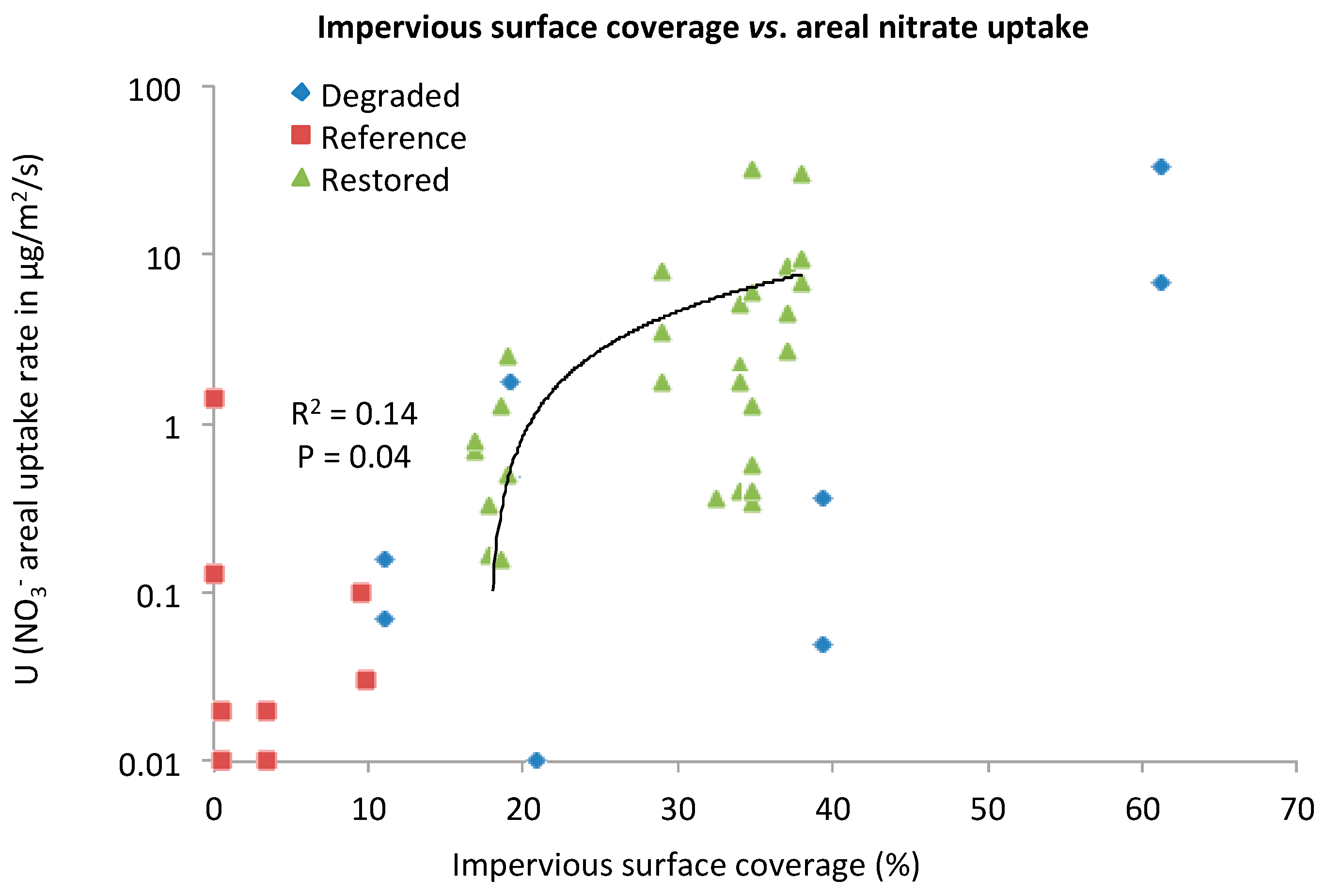
3.1. Evaluating Potential Controlling Factors of Nutrient Uptake
3.2. Nutrient Spiraling Results
3.2.1. Nitrate
Nitrate Uptake Length (SW)
Nitrate Areal Uptake Rate (U)
Nitrate Uptake Velocity (Vf)
3.2.2. Ammonium
Ammonium Uptake Length (SW)
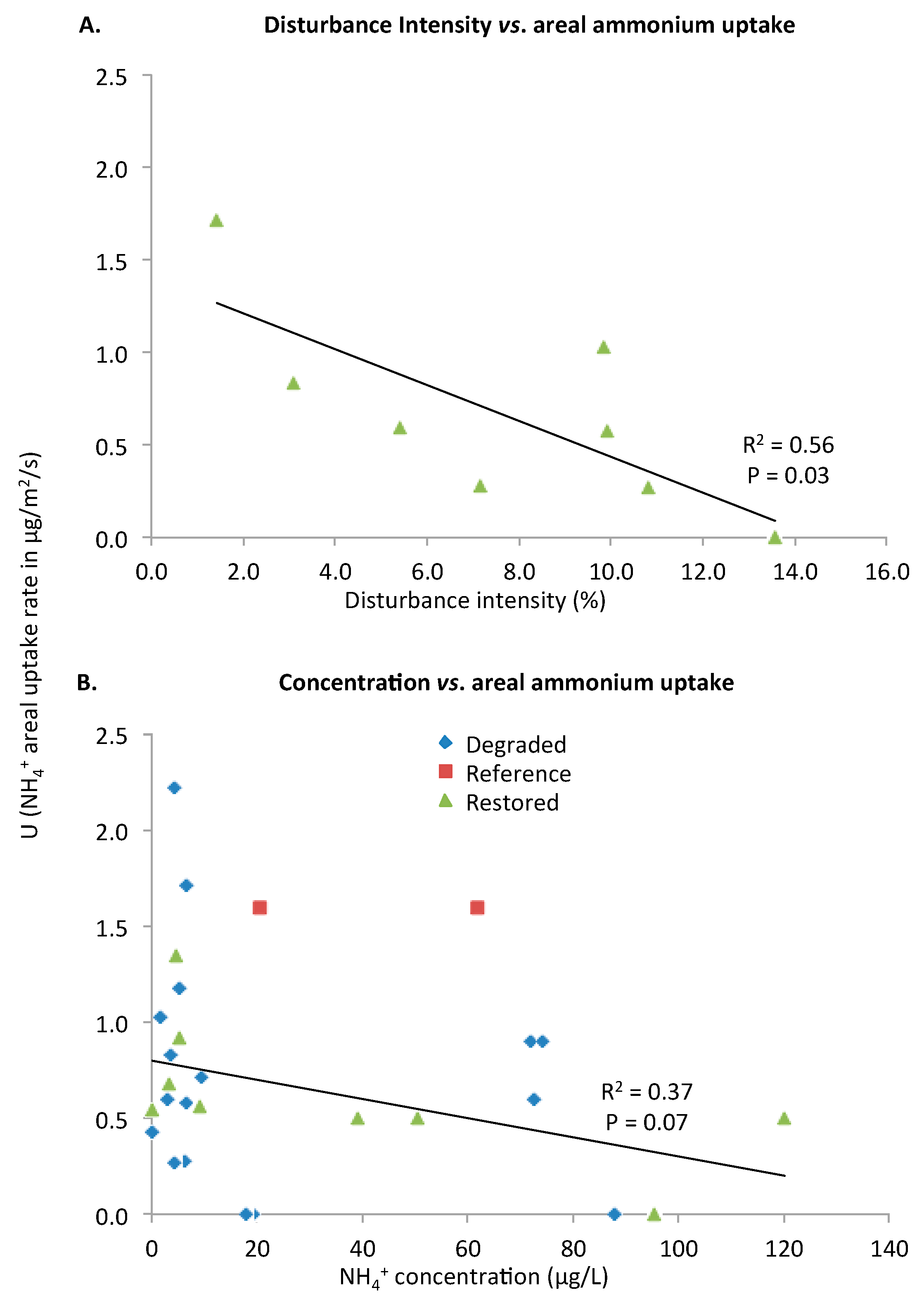
Ammonium Areal Uptake Rate (U)
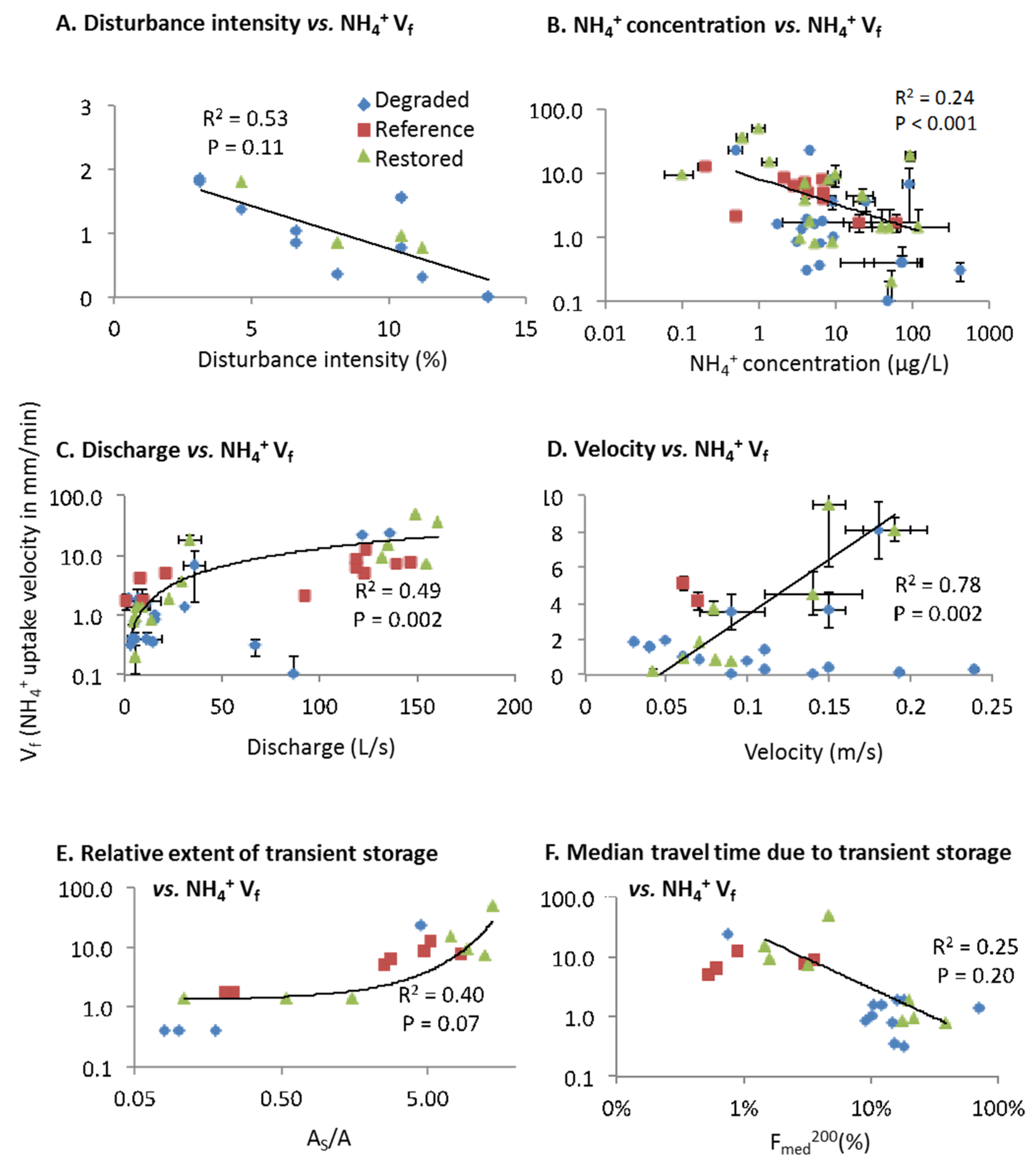
Ammonium Uptake Velocity (Vf)
3.2.3. Soluble Reactive Phosphorus (SRP)
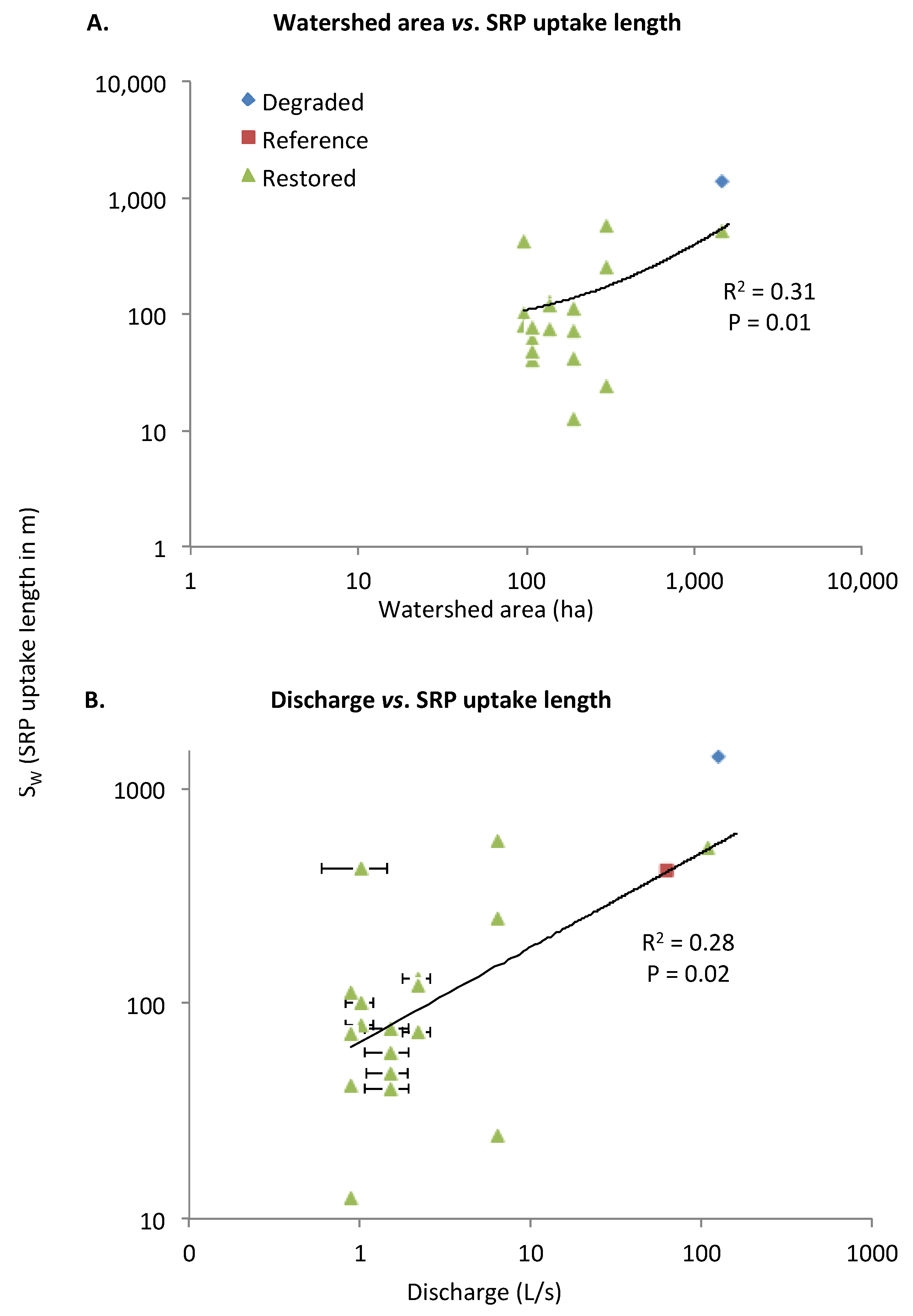
SRP Uptake Length (SW) and Areal Uptake Rate (U)
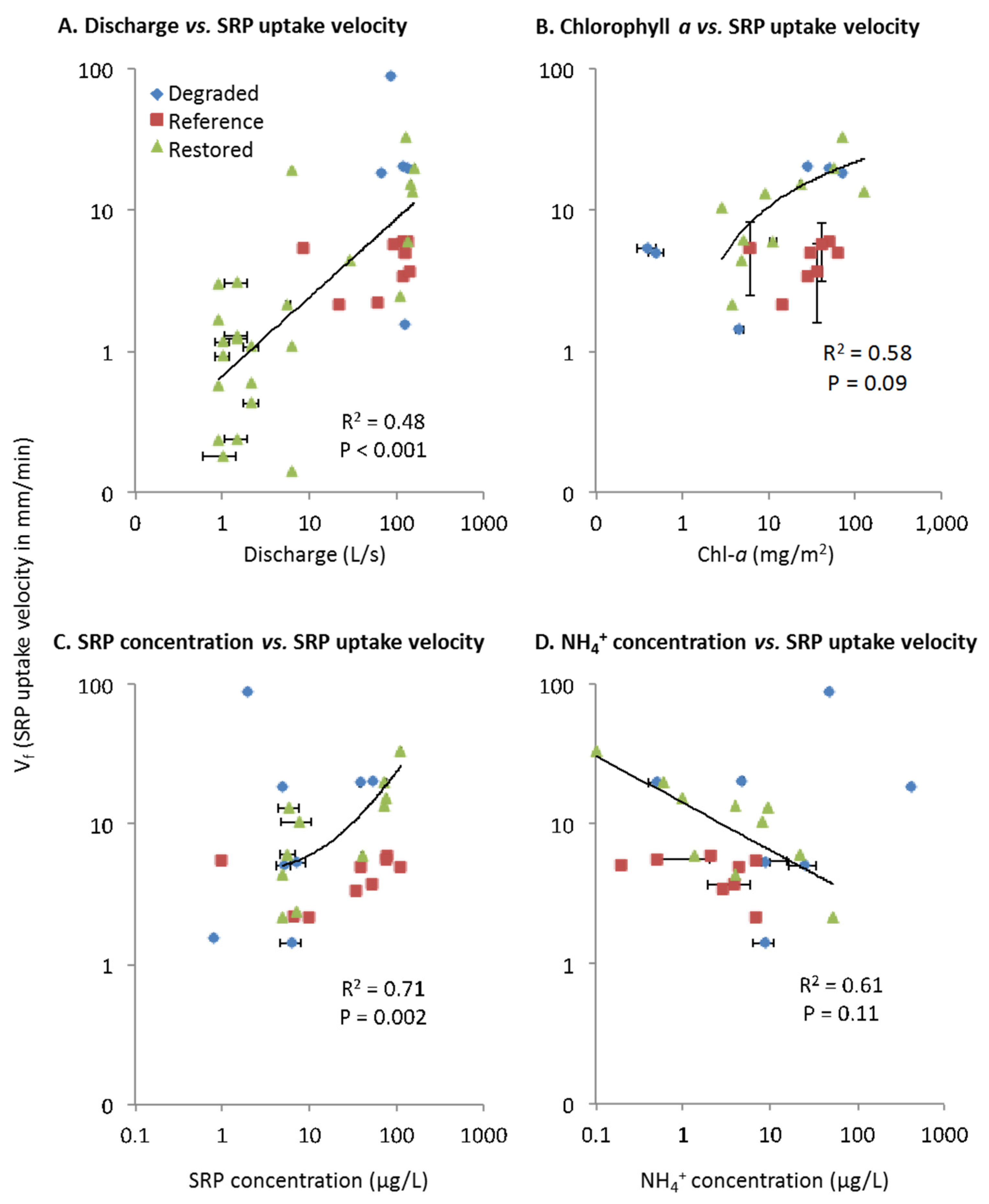
SRP Uptake Velocity (Vf)
4. Discussion
4.1. Nutrient Retention in Restored Streams Over Time: Urban Succession
4.2. Low Nutrient Uptake Rates in AMD Remediated Streams
4.3. Size Matters: Optimizing Reactive Sediment Volume and Transient Storage
4.4. Restored Riffles, Substrate, and Coarse Woody Debris
5. Conclusions and Management Implications
Knowledge Gaps and Future Directions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- National Academy of Engineering Manage the nitrogen Cycle—Engineering Challenges. Available online: http://www.engineeringchallenges.org/cms/8996/9132.aspx (accessed on 25 October 2014).
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Ecology: Controlling Eutrophication: Nitrogen and Phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Green, P.A.; Vörösmarty, C.J.; Meybeck, M.; Galloway, J.N.; Peterson, B.J.; Boyer, E.W. Pre-industrial and contemporary fluxes of nitrogen through rivers: A global assessment based on typology. Biogeochemistry 2004, 68, 71–105. [Google Scholar] [CrossRef]
- Caraco, N.F. Disturbance of the phosphorus cycle: A case of indirect effects of human activity. Trends Ecol. Evol. 1993, 8, 51–54. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human Alteration of the Global Nitrogen Cycle: Sources and Consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Galloway, J.N.; Aber, J.D.; Erisman, J.W.; Seitzinger, S.P.; Howarth, R.W.; Cowling, E.B.; Cosby, B.J. The nitrogen cascade. BioScience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Conley, D.J. Biogeochemical nutrient cycles and nutrient management strategies. Hydrobiologia 1999, 410, 87–96. [Google Scholar] [CrossRef]
- Bennett, E.M.; Carpenter, S.R.; Caraco, N.F. Human impact on erodable phosphorus and eutrophication: A global perspective increasing accumulation of phosphorus in soil threatens rivers, lakes, and coastal oceans with eutrophication. BioScience 2001, 51, 227–234. [Google Scholar] [CrossRef]
- Howarth, R.W.; Sharpley, A.; Walker, D. Nutrient Pollution to Coastal Waters in the United States: Implications for Achieving Coastal Water Quality Goals. Estuaries 2002, 25, 656–676. [Google Scholar] [CrossRef]
- Kemp, W.M.; Boynton, W.R.; Adoli, J.E.; Boesch, D.F.; Boicourt, W.C.; Brush, G.S.; Cornwell, J.C.; Fisher, T.R.; Glibert, P.M.; Hagy, J.D.; et al. Eutrophication of Chesapeake Bay: Historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005, 303, 1–29. [Google Scholar] [CrossRef]
- Boesch, D.F.; Brinsfield, R.B.; Magnien, R.E. Chesapeake Bay eutrophication: Scientific understanding, ecosystem restoration, and challenges for agriculture. J. Environ. Qual. 2001, 30, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S.; Groffman, P.M.; Band, L.E.; Shields, C.A.; Morgan, R.P.; Palmer, M.A.; Belt, K.T.; Swan, C.M.; Findlay, S.E.G.; Fisher, G.T. Interaction between Urbanization and Climate Variability Amplifies Watershed Nitrate Export in Maryland. Environ. Sci. Technol. 2008, 42, 5872–5878. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.J.; Rosenberg, R. Spreading Dead Zones and Consequences for Marine Ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, X.; Liu, L.; Zhang, W.; Guo, P. Algae community and trophic state of subtropical reservoirs in southeast Fujian, China. Environ. Sci. Pollut. Res. 2012, 19, 1432–1442. [Google Scholar] [CrossRef] [PubMed]
- Howarth, R.W.; Swaney, D.P.; Boyer, E.W.; Marino, R.; Jaworski, N.; Goodale, C. The influence of climate on average nitrogen export from large watersheds in the Northeastern United States. Biogeochemistry 2006, 79, 163–186. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Mayer, P.M.; Vidon, P.G.; Smith, R.M.; Pennino, M.J.; Newcomer, T.A.; Duan, S.; Welty, C.; Belt, K.T. Land Use and Climate Variability Amplify Carbon, Nutrient, and Contaminant Pulses: A Review with Management Implications. J. Am. Water Resour. Assoc. 2014, 50, 585–614. [Google Scholar] [CrossRef]
- Collins, K.A.; Lawrence, T.J.; Stander, E.K.; Jontos, R.J.; Kaushal, S.S.; Newcomer, T.A.; Grimm, N.B.; Cole Ekberg, M.L. Opportunities and challenges for managing nitrogen in urban stormwater: A review and synthesis. Ecol. Eng. 2010, 36, 1507–1519. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Groffman, P.M.; Band, L.E.; Elliott, E.M.; Shields, C.A.; Kendall, C. Tracking nonpoint source nitrogen pollution in human-impacted watersheds. Environ. Sci. Technol. 2011, 45, 8225–8232. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Kaushal, S.S.; Groffman, P.M.; Band, L.E.; Belt, K.T. Phosphorus export across an urban to rural gradient in the Chesapeake Bay watershed. J. Geophys. Res. 2012, 117, G01025. [Google Scholar] [CrossRef]
- Stets, E.G.; Kelly, V.J.; Crawford, C.G. Regional and Temporal Differences in Nitrate Trends Discerned from Long-Term Water Quality Monitoring Data. J. Am. Water Resour. Assoc. 2015, 51, 1394–1407. [Google Scholar] [CrossRef]
- Moomaw, W.R. Energy, industry and nitrogen: Strategies for decreasing reactive nitrogen emissions. AMBIO J. Hum. Environ. 2002, 31, 184–189. [Google Scholar] [CrossRef]
- Dinnes, D.L.; Karlen, D.L.; Jaynes, D.B.; Kaspar, T.C.; Hatfield, J.L.; Colvin, T.S.; Cambardella, C.A. Nitrogen management strategies to reduce nitrate leaching in tile-drained midwestern soils. Agron. J. 2002, 94, 153–171. [Google Scholar] [CrossRef]
- Maxted, J.T.; Diebel, M.W.; Vander Zanden, M.J. Landscape Planning for Agricultural Non-Point Source Pollution Reduction. II. Balancing Watershed Size, Number of Watersheds, and Implementation Effort. Environ. Manag. 2009, 43, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.F.; Jones, R.A. Detergent phosphate bans and eutrophication. Environ. Sci. Technol. 1986, 20, 330–331. [Google Scholar] [CrossRef] [PubMed]
- Howarth, R.W. Coastal nitrogen pollution: A review of sources and trends globally and regionally. Harmful Algae 2008, 8, 14–20. [Google Scholar] [CrossRef]
- Søvik, A.K.; Syversen, N. Retention of particles and nutrients in the root zone of a vegetative buffer zone: Effect of vegetation and season. Boreal Environ. Res. 2008, 13, 223–230. [Google Scholar]
- Weigelhofer, G.; Fuchsberger, J.; Teufl, B.; Welti, N.; Hein, T. Effects of Riparian Forest Buffers on In-Stream Nutrient Retention in Agricultural Catchments. J. Environ. Qual. 2012, 41, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Bukaveckas, P.A. Effects of channel restoration on water velocity, transient storage, and nutrient uptake in a channelized stream. Environ. Sci. Technol. 2007, 41, 1570–1576. [Google Scholar] [CrossRef] [PubMed]
- Klocker, C.A.; Kaushal, S.S.; Groffman, P.M.; Mayer, P.M.; Morgan, R.P. Nitrogen uptake and denitrification in restored and unrestored streams in urban Maryland, USA. Aquat. Sci. 2009, 71, 411–424. [Google Scholar] [CrossRef]
- Pennino, M.P.; Kaushal, S.S.; Beaulieu, J.J.; Mayer, P.M.; Arango, C.P. Effects of urban stream burial on nitrogen uptake and ecosystem metabolism: Implications for watershed nitrogen and carbon fluxes. Biogeochemistry 2014, 121, 247–269. [Google Scholar] [CrossRef]
- Melzer, A.; Exler, D. Nitrate and Nitrite Reductase Activities in Aquatic Macrophytes. Stud. Aquat. Vasc. Plants Royal Botanic Society of Belgium, Brussels. 1982, 128–135. [Google Scholar]
- Froelich, P. Kinetic Control of Dissolved Phosphate in Natural Rivers and Estuaries—A Primer on the Phosphate Buffer Mechanism. Limnol. Oceanogr. 1988, 33, 649–668. [Google Scholar] [CrossRef]
- House, W.A.; Denison, F.H. Exchange of inorganic phosphate between river waters and bed-sediments. Environ. Sci. Technol. 2002, 36, 4295–4301. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Kaushal, S.S. Warming increases carbon and nutrient fluxes from sediments in streams across land use. Biogeosciences 2013, 10, 1193–1207. [Google Scholar] [CrossRef]
- Davidson, E.A.; Schimel, J.P. Microbial processes of production and consumption of nitric oxide, nitrous oxide and methane. In Biogenic Trace Gases: Measuring Emissions from Sediment and Water; Matson, P.A., Harriss, R.C., Eds.; Blackwell Science: Cambridge, MA, USA, 1995; pp. 327–357. [Google Scholar]
- Groffman, P.M.; Dorsey, A.M.; Mayer, P.M. N processing within geomorphic structures in urban streams. J. N. Am. Benthol. Soc. 2005, 24, 613–625. [Google Scholar] [CrossRef]
- Boyer, E.W.; Alexander, R.B.; Parton, W.J.; Li, C.; Butterbach-Bahl, K.; Donner, S.D.; Skaggs, R.W.; Grosso, S.J.D. Modeling denitrification in terrestrial and aquatic ecosystems at regional scales. Ecol. Appl. 2006, 16, 2123–2142. [Google Scholar] [CrossRef]
- Hedin, L.O.; von Fischer, J.C.; Ostrom, N.E.; Kennedy, B.P.; Brown, M.G.; Robertson, G.P. Thermodynamic constraints on nitrogen transformations and other biogeochemical processes at soil-stream interfaces. Ecology 1998, 79, 684–703. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Groffman, P.M.; Mayer, P.M.; Striz, E.; Gold, A.J. Effects of stream restoration on denitrification in an urbanizing watershed. Ecol. Appl. 2008, 18, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Mayer, P.M.; Groffman, P.M.; Striz, E.A.; Kaushal, S.S. Nitrogen dynamics at the groundwater-surface water interface of a degraded urban stream. J. Environ. Qual. 2010, 39, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Forshay, K.J.; Dodson, S.I. Macrophyte presence is an indicator of enhanced denitrification and nitrification in sediments of a temperate restored agricultural stream. Hydrobiologia 2011, 668, 21–34. [Google Scholar] [CrossRef]
- Roley, S.S.; Tank, J.L.; Williams, M.A. Hydrologic connectivity increases denitrification in the hyporheic zone and restored floodplains of an agricultural stream. J. Geophys. Res. 2012, 117, G00N04. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Belt, K.T. The urban watershed continuum: Evolving spatial and temporal dimensions. Urban Ecosyst. 2012, 15, 409–435. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Helton, A.M.; Poole, G.C.; Hall, R.O.; Hamilton, S.K.; Peterson, B.J.; Tank, J.L.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Stream denitrification across biomes and its response to anthropogenic nitrate loading. Nature 2008, 452, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.E. Effects of agriculture on erosion and sedimentation in the Piedmont Province, Maryland. Geol. Soc. Am. Bull. 1975, 86, 1281–1286. [Google Scholar] [CrossRef]
- Magilligan, F.J. Historical floodplain sedimentation in the Galena River basin, Wisconsin and Illinois. Ann. Assoc. Am. Geogr. 1985, 75, 583–594. [Google Scholar] [CrossRef]
- Knox, J.C. Historical valley floor sedimentation in the Upper Mississippi Valley. Ann. Assoc. Am. Geogr. 1987, 77, 224–244. [Google Scholar] [CrossRef]
- Parola, A.C.; Vesely, W.S.; Croasdaile, M.A.; Hansen, C.; Jones, M.S. Geomorphic Characteristics of Streams in the Bluegrass Physiographic Region of Kentucky; Technical Report for Nonpoint Source Implementation Program of University of Louisville: Louisville, KY, USA, 2007. [Google Scholar]
- Walter, R.C.; Merritts, D.J. Natural streams and the legacy of water-powered mills. Science 2008, 319, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Nogaro, G.; Datry, T.; Mermillod-Blondin, F.; Descloux, S.; Montuelle, B. Influence of streambed sediment clogging on microbial processes in the hyporheic zone. Freshw. Biol. 2010, 55, 1288–1302. [Google Scholar] [CrossRef]
- Kasahara, T.; Hill, A. Modeling the effects of lowland stream restoration projects on stream–subsurface water exchange. Ecol. Eng. 2008, 32, 310–319. [Google Scholar] [CrossRef]
- Verhoeven, J.T. A.; Arheimer, B.; Yin, C.Q.; Hefting, M.M. Regional and global concerns over wetlands and water quality. Trends Ecol. Evol. 2006, 21, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P., II. The urban stream syndrome: Current knowledge and the search for a cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Elmore, A.J.; Kaushal, S.S. Disappearing headwaters: Patterns of stream burial due to urbanization. Front. Ecol. Environ. 2008, 6, 308–312. [Google Scholar] [CrossRef]
- Roley, S.S.; Tank, J.L.; Stephen, M.L.; Johnson, L.T.; Beaulieu, J.J.; Witter, J.D. Floodplain restoration enhances denitrification and reach-scale nitrogen removal in an agricultural stream. Ecol. Appl. 2012, 22, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, E.S.; Palmer, M.A. River restoration: The fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecol. Appl. 2011, 21, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Mayer, P.M.; Reynolds, S.K.; McCutchen, M.D.; Canfield, T.J. Meta-Analysis of Nitrogen Removal in Riparian Buffers. J. Environ. Qual. 2007, 36, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.J.; Mulholland, P.J.; Houser, J.N. Effects of upland disturbance and instream restoration on hydrodynamics and ammonium uptake in headwater streams. J. N. Am. Benthol. Soc. 2007, 26, 38–53. [Google Scholar] [CrossRef]
- Theriot, J.M.; Conkle, J.L.; Reza Pezeshki, S.; DeLaune, R.D.; White, J.R. Will hydrologic restoration of Mississippi River riparian wetlands improve their critical biogeochemical functions? Ecol. Eng. 2013, 60, 192–198. [Google Scholar] [CrossRef]
- Garcia-Linares, C.; Martinez-Santos, M.; Martinez-Bilbao, V.; Sanchez-Perez, J.M.; Antiguedad, I. Wetland restoration and nitrate reduction: The example of the peri-urban wetland of Vitoria-Gasteiz (Basque Country, North Spain). Hydrol. Earth Syst. Sci. 2003, 7, 109–121. [Google Scholar] [CrossRef]
- Wolf, K.L.; Noe, G.B.; Ahn, C. Hydrologic Connectivity to Streams Increases Nitrogen and Phosphorus Inputs and Cycling in Soils of Created and Natural Floodplain Wetlands. J. Environ. Qual. 2013, 42, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Audet, J.; Hoffmann, C.C.; Jensen, H.S. Low nitrogen and phosphorus release from sediment deposited on a Danish restored floodplain. Ann. Limnol. Int. J. Limnol. 2011, 47, 231–238. [Google Scholar] [CrossRef]
- Troxler Gann, T.G.; Childers, D.L.; Rondeau, D.N. Ecosystem structure, nutrient dynamics, and hydrologic relationships in tree islands of the southern Everglades, Florida, USA. For. Ecol. Manag. 2005, 214, 11–27. [Google Scholar] [CrossRef]
- Akamatsu, F.; Shimano, K.; Denda, M.; Ide, K.; Ishihara, M.; Toda, H. Effects of sediment removal on nitrogen uptake by riparian plants in the higher floodplain of the Chikuma River, Japan. Landsc. Ecol. Eng. 2008, 4, 91–96. [Google Scholar] [CrossRef]
- Aspetsberger, F.; Huber, F.; Kargl, S.; Scharinger, B.; Peduzzi, P.; Hein, T. Particulate organic matter dynamics in a river floodplain system: Impact of hydrological connectivity. Arch. Hydrobiol. 2002, 156, 23–42. [Google Scholar] [CrossRef]
- Newcomer, T.A.; Kaushal, S.S.; Mayer, P.M.; Shields, A.R.; Canuel, E.A.; Groffman, P.M.; Gold, A.J. Influence of natural and novel organic carbon sources on denitrification in forest, degraded urban, and restored streams. Ecol. Monogr. 2012, 82, 449–466. [Google Scholar] [CrossRef]
- Rohatgi, A. WebPlotDigitizer: Web Based Tool to Extract Data from Plots, Images, and Maps. Available online: http://arohatgi.info/WebPlotDigitizer/userManual.pdf (accessed on 15 November 2015).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Stream Solute Workshop. Concepts and methods for assessing solute dynamics in stream ecosystems. J. N. Am. Benthol. Soc. 1990, 9, 95–119. [Google Scholar]
- Gomez-Velez, J.D.; Harvey, J.W. A hydrogeomorphic river network model predicts where and why hyporheic exchange is important in large basins. Geophys. Res. Lett. 2014, 41, 6403–6412. [Google Scholar] [CrossRef]
- Weigelhofer, G.; Welti, N.; Hein, T. Limitations of stream restoration for nitrogen retention in agricultural headwater streams. Ecol. Eng. 2013, 60, 224–234. [Google Scholar]
- Northington, R.M.; Benfield, E.F.; Schoenholtz, S.H.; Timpano, A.J.; Webster, J.R.; Zipper, C. An assessment of structural attributes and ecosystem function in restored Virginia coalfield streams. Hydrobiologia 2011, 671, 51–63. [Google Scholar] [CrossRef]
- Hines, S.L.; Hershey, A.E. Do channel restoration structures promote ammonium uptake and improve macroinvertebrate-based water quality classification in urban streams? Inland Waters 2011, 1, 133–145. [Google Scholar] [CrossRef]
- Kasahara, T.; Hill, A.R. Effects of riffle-step restoration on hyporheic zone chemistry in N-rich lowland streams. Can. J. Fish. Aquat. Sci. 2006, 63, 120–133. [Google Scholar] [CrossRef]
- Filoso, S.; Palmer, M.A. Assessing stream restoration effectiveness at reducing nitrogen export to downstream waters. Ecol. Appl. 2011, 21, 1989–2006. [Google Scholar] [CrossRef] [PubMed]
- Newcomer Johnson, T.A.; Kaushal, S.S.; Mayer, P.M.; Grese, M.M. Effects of stormwater management and stream restoration on watershed nitrogen retention. Biogeochemistry 2014, 121, 81–106. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Hassett, B.A.; Cada, P.; Bernhardt, E.S. Testing the field of dreams hypothesis: Functional responses to urbanization and restoration in stream ecosystems. Ecol. Appl. 2011, 21, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Argerich, A.; Martí, E.; Sabater, F.; Haggerty, R.; Ribot, M. Influence of transient storage on stream nutrient uptake based on substrata manipulation. Aquat. Sci. 2011, 73, 365–376. [Google Scholar] [CrossRef]
- Arango, C.P.; James, P.W.; Hatch, K.B. Rapid ecosystem response to restoration in an urban stream. Hydrobiologia 2015, 749, 197–211. [Google Scholar] [CrossRef]
- McMillan, S.K.; Tuttle, A.K.; Jennings, G.D.; Gardner, A. Influence of Restoration Age and Riparian Vegetation on Reach-Scale Nutrient Retention in Restored Urban Streams. JAWRA J. Am. Water Resour. Assoc. 2014, 50, 626–638. [Google Scholar] [CrossRef]
- Hoellein, T.J.; Tank, J.L.; Entrekin, S.A.; Rosi-Marshall, E.J.; Stephen, M.L.; Lamberti, G.A. Effects of benthic habitat restoration on nutrient uptake and ecosystem metabolism in three headwater streams: Stream restoration and ecosystem function. River Res. Appl. 2012, 28, 1451–1461. [Google Scholar] [CrossRef]
- Bott, T.L.; Jackson, J.K.; McTammany, M.E.; Newbold, D.; Rier, S.T.; Sweeney, B.W.; Battle, J.M. Abandoned coal mine drainage and its remediation: Impacts on stream ecosystem structure and function. Ecol. Appl. 2012, 22, 2144–2163. [Google Scholar] [CrossRef] [PubMed]
- Ensign, S.H.; Doyle, M.W. Nutrient spiraling in streams and river networks. J. Geophys. Res. Biogeosci. 2006, 111, G04009. [Google Scholar] [CrossRef]
- Schueler, T.R.; Fraley-McNeal, L.; Cappiella, K. Is Impervious Cover Still Important? Review of Recent Research. J. Hydrol. Eng. 2009, 14, 309–315. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]
- Kaushal, S.S.; McDowell, W.H.; Wollheim, W.M.; Johnson, T.A. N.; Mayer, P.M.; Belt, K.T.; Pennino, M.J. Urban Evolution: The Role of Water. Water 2015, 7, 4063–4087. [Google Scholar] [CrossRef]
- Kaushal, S.S.; McDowell, W.H.; Wollheim, W.M. Tracking evolution of urban biogeochemical cycles: Past, present, and future. Biogeochemistry 2014, 121, 1–21. [Google Scholar] [CrossRef]
- Doyle, M.W.; Singh, J.; Lave, R.; Robertson, M.M. The morphology of streams restored for market and nonmarket purposes: Insights from a mixed natural-social science approach. Water Resour. Res. 2015, 51, 5603–5622. [Google Scholar] [CrossRef]
- Grimm, N.B. Nitrogen dynamics during succession in a desert stream. Ecology 1987, 68, 1157–1170. [Google Scholar] [CrossRef]
- Ensign, S.H.; Doyle, M.W. In-channel transient storage and associated nutrient retention: Evidence from experimental manipulations. Limnol. Oceanogr. 2005, 50, 1740–1751. [Google Scholar] [CrossRef]
- Smith, J.J.; Lake, P.S. The breakdown of buried and surface-placed leaf litter in an upland stream. Hydrobiologia 1993, 271, 141–148. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; Mayer, P.M.; Kaushal, S.S.; Pennino, M.P.; Arango, C.P.; Balz, D.A.; Fritz, K.M.; Hill, B.H.; Elonen, C.M.; Santo Domingo, J.W.; et al. Effects of urban stream burial on organic matter dynamics and reach scale nitrate retention. Biogeochemistry 2014, 121, 107–126. [Google Scholar] [CrossRef]
- Valett, H.M.; Morrice, J.A.; Dahm, C.N.; Campana, M.E. Parent lithology, surface–groundwater exchange, and nitrate retention in headwater streams. Limnol. Oceanogr. 1996, 41, 333–345. [Google Scholar] [CrossRef]
- Thomas, S.A.; Valett, H.M.; Webster, J.R.; Mulholland, P.J. A regression approach to estimating reactive solute uptake in advective and transient storage zones of stream ecosystems. Adv. Water Resour. 2003, 26, 965–976. [Google Scholar] [CrossRef]
- Jordan, T.E.; Andrews, M.P.; Szuch, R.P.; Whigham, D.F.; Weller, D.E.; Jacobs, A.D. Comparing functional assessments of wetlands to measurements of soil characteristics and nitrogen processing. Wetlands 2007, 27, 479–497. [Google Scholar] [CrossRef]
- Lautz, L.K.; Siegel, D.I. The effect of transient storage on nitrate uptake lengths in streams: An inter-site comparison. Hydrol. Process. 2007, 21, 3533–3548. [Google Scholar] [CrossRef]
- Hall, R.O., Jr.; Bernhardt, E.S.; Likens, G.E. Relating nutrient uptake with transient storage in forested mountain streams. Limnol. Oceanogr. 2002, 47, 255–265. [Google Scholar] [CrossRef]
- Webster, J.R.; Mulholland, P.J.; Tank, J.L.; Valett, H.M.; Dodds, W.K.; Peterson, B.J.; Bowden, W.B.; Dahm, C.N.; Findlay, S.; Gregory, S.V.; et al. Factors affecting ammonium uptake in streams—An inter-biome perspective. Freshw. Biol. 2003, 48, 1329–1352. [Google Scholar] [CrossRef]
- Dodds, W.K.; Martí, E.; Tank, J.L.; Pontius, J.; Hamilton, S.K.; Grimm, N.B.; Bowden, W.B.; McDowell, W.H.; Peterson, B.J.; Valett, H.M.; et al. Carbon and nitrogen stoichiometry and nitrogen cycling rates in streams. Oecologia 2004, 140, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Goodale, C.L.; Aber, J.D.; Vitousek, P.M.; McDowell, W.H. Long-term decreases in stream nitrate: Successional causes unlikely; possible links to DOC? Ecosystems 2005, 8, 334–337. [Google Scholar] [CrossRef]
- Harrison, M.D.; Groffman, P.M.; Mayer, P.M.; Kaushal, S.S. Microbial biomass and activity in geomorphic features in forested and urban restored and degraded streams. Ecol. Eng. 2012, 38, 1–10. [Google Scholar] [CrossRef]
- Powers, S.M.; Johnson, R.A.; Stanley, E.H. Nutrient Retention and the Problem of Hydrologic Disconnection in Streams and Wetlands. Ecosystems 2012, 15, 435–449. [Google Scholar] [CrossRef]
- Merwade, V.M.; Maidment, D.R.; Hodges, B.R. Geospatial representation of river channels. J. Hydrol. Eng. 2005, 10, 243–251. [Google Scholar] [CrossRef]
- Abdelnour, A.; McKane, B.R.; Stieglitz, M.; Pan, F.; Cheng, Y. Effects of harvest on carbon and nitrogen dynamics in a Pacific Northwest forest catchment. Water Resour. Res. 2013, 49, 1292–1313. [Google Scholar] [CrossRef]
- Abdelnour, A.; Stieglitz, M.; Pan, F.; McKane, R. Catchment hydrological responses to forest harvest amount and spatial pattern. Water Resour. Res. 2011, 47, W09521. [Google Scholar] [CrossRef]
- McKane, R.B. US EPA Sustainable and Healthy Communities Research: A Pacific Northwest Demonstration Study; United States Environmental Protection Agency: Washington, DC, USA, 2014.
- McKane, R.B. US EPA Enhanced Version of Velma Eco-Hydrological Modeling and Decision Support Framework to Address Engineered and Natural Applications of Green Infrastructure for Reducing Nonpoint Inputs of Nutrients and Contaminants; United States Environmental Protection Agency: Washington, DC, USA, 2014.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strategies Used to Increase Hydrologic Connectivity | Typologies Included from Figure 2 | Number of Results from 79 Studies | Positive Results (%) | Neutral Results (%) | Negative Results (%) |
|---|---|---|---|---|---|
| Floodplain Reconnection | ABCD | 62 | 60% | 28% | 12% |
| Streambed Reconnection | EF | 9 | 70% | 20% | 10% |
| Increased Stream Surface Area | G | 19 | 65% | 22% | 13% |
| Increased Wetland Surface Area | HI | 24 | 75% | 14% | 11% |
| Total | 114 | 62% | 26% | 12% |
| Citation | Tracer Used | Uptake Metrics Recorded | Description of Study Streams | N | (Typology from Figure 2: Rating)—Summary of Nutrient Spiraling Results | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NH4+ | NO3− | SRP | k | SW | U | Vf | % | Other | ||||
| [72] | X | X | X | X | X | Pristine, restored, broadened, and incised streams | 31 | (ABG: Positive and Negative)—Restored and pristine reaches had significantly shorter SW and larger Vf than channelized reaches, and NH4+ uptake was positively correlated with transient storage. | ||||
| [59] | X | X | X | X | Coarse woody debris treatment and control | 16 | (A: Positive)—Coarse woody debris treatments had significantly higher uptake than the control (Vf increased by 23%–154% and U by 61%–235%). | |||||
| [73] | X | X | Acid mine drainage (AMD) | 9 | (AC: Neutral)—All streams were net heterotrophic with varying levels of NH4+ uptake. No site differences were found. | |||||||
| [74] | X | X | X | X | Restored and unrestored reference streams | 12 | (AB: Positive)—Significantly shorter NH4+ SW was observed in restored compared to unrestored sites 2 years post-restoration likely due to greater biofilm development on larger substrates with less canopy cover. There was not a significant change to U or Vf. | |||||
| [79] | X | X | X | λ λs | Control stream plus 4 treatments | 24 | (A: Neutral)—Substrate treatment increased transient storage zone and decreased velocity in 20 m reaches but did not significantly affect larger reach. | |||||
| [80] | X | X | X | Pre-restoration, restored, and reference | 14 | (GI: Positive and Neutral)—After stream restoration, nutrient demand spiked to levels that have rarely been reported, but demand recovered within 35 days. | ||||||
| [75] | X | X | g/day | Man-made riffles/step vs. natural riffle | 4 | (A: Positive)—Natural riffle had greater NO3− % removal than constructed riffle, but constructed riffle removed 3 times more due to larger hyporheic exchange flux. | ||||||
| [76] | X | X | X | X | 3 restored streams | 6 | (AC: Positive and Neutral)—Doubling tracer N concentration increased SW and decreased U & Vf | |||||
| [30] | X | X | X | X | X | X | 2 degraded and 2 restored streams | 5 | (ABEG: Positive)—SW increased with velocity | |||
| [77] | X | X | X | X | Restored, urban, and forest streams | 6 | (ABEH: Positive)—Vf and U were greater in stream reaches than adjacent stormwater control measures | |||||
| [78] | X | X | X | 3 restored, 3 urban degraded, & 3 forest streams | 24 | (A: Positive and Neutral)—In summer, restored reaches had higher uptake rates than unrestored/forested reaches; Temperature and % canopy cover explained 80% of the variation in uptake. | ||||||
| [29] | X | X | X | X | X | Channelized, restored, and reference reach | 44 | (AGI: Positive)—Lowering velocity and raising transient storage in restored stream increased uptake but difference was not statistically significant. | ||||
| [81] | X | X | X | X | X | 5 streams restored from 2002–2010 | 15 | (CG: Positive and Neutral)—P uptake was greater in newly restored sites (attributed to assimilation by algal biofilms), whereas NO3− uptake was highest in older sites potentially due to greater channel stability and establishment of microbial communities. | ||||
| [82] | X | X | X | X | Restored and reference | 24 | (A: Positive and Neutral)—Increases in gravel, cobble and boulder habitat in the restoration reaches were correlated with higher rates of nutrient uptake and metabolism. | |||||
| [83] | X | X | X | X | Acid mine drainage (AMD) degraded, restored, and reference | 6 | (HI: Positive and Negative)—Acid Mine Drainage (AMD) remediation restored NH4+ uptake, reduced NO3− uptake to undetectable level, and restored SRP uptake to near normal rates. | |||||
| Sum | 8 | 9 | 6 | 4 | 7 | 9 | 12 | 3 | 2 | |||
| Stream Type | NO3− | NH4+ | SRP | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SW | U | Vf | SW | U | Vf | SW | U | Vf | ||
| (m) | (μg/m2/s) | (mm/min) | (m) | (μg/m2/s) | (mm/min) | (m) | (μg/m2/s) | (mm/min) | ||
| Restored | Mean | 316 | 5.2 | 2.2 | 245.6 | 0.6 | 9.4 | 153.2 | 13.8 | 5.7 |
| Median | 136 | 1.8 | 1.1 | 0.5 | 4.1 | 77.8 | 3.4 | 1.9 | ||
| Range | 34–2668 | 0.15–32 | 0.0–8.9 | 70–421 | 0.0–1.4 | 0.2–49 | 12–572 | 0.3–117 | 0.1–33 | |
| Number | 25 | 32 | 36 | 2 | 9 | 18 | 18 | 17 | 28 | |
| Degraded | Mean | 3107 | 5.3 | 3.0 | 609.5 | 0.7 | 3.5 | 19.9 | ||
| Median | 1341 | 0.42 | 1.0 | 789.5 | 0.6 | 1.0 | 1403 | 11.8 | ||
| Range | 108–18,632 | 0.01–33.6 | 0.02–38.2 | 197–842 | 0.0–2.2 | 0.0–22.8 | 1.4–87.4 | |||
| Number | 13 | 12 | 24 | 3 | 17 | 23 | 1 | 0 | 8 | |
| Reference | Mean | 2714 | 0.19 | 3.3 | 3.9 | 4.2 | ||||
| Median | 345 | 0.03 | 0.4 | 210.5 | 1.6 | 0.4 | 413 | 4.9 | ||
| Range | 238–7558 | 0.00–1.43 | 0.03–35 | 0.03–35 | 2.2–5.9 | |||||
| Number | 3 | 10 | 13 | 1 | 1 | 11 | 1 | 0 | 9 | |
| ANOVA Comparisons Between Stream Types (p Value < 0.12) | ||||||||||
| Restored vs. Degraded | p = 0.03 | n.s. | n.s. | n.s. | p = 0.07 | p = 0.11 | n.s. | n.s. | p = 0.03 | |
| Degraded vs. Reference | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | p = 0.05 | |
| Reference vs. Restored | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Tracer Used | Uptake Metrics Recorded | Watershed | Reach | Concentration | Transient Storage | Metabolism | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Watershed Area | % Impervious | % Disturbance | Width | Discharge | Velocity | NH4+ | SRP | AS/A | Fmed200(%) | Chl-a | ||
| NO3− | SW | <0.001 * (+) | 0.002 * (+) | |||||||||
| U | 0.04 (+) | |||||||||||
| Vf | ||||||||||||
| NH4+ | SW | |||||||||||
| U | 0.03 (−) | 0.07 (−) | ||||||||||
| Vf | 0.11 (−) | 0.002 (+) | 0.002 (+) | <0.001 (−) | 0.07 (+) | 0.20 (−) | ||||||
| SRP | SW | 0.01 (+) | 0.02 (+) | |||||||||
| U | ||||||||||||
| Vf | <0.001 (+) | 0.11 (−) | 0.002 (+) | 0.04 (+) | ||||||||
| No. of Correlations: | 2 | 1 | 2 | 1 | 3 | 1 | 3 | 1 | 1 | 1 | 1 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Newcomer Johnson, T.A.; Kaushal, S.S.; Mayer, P.M.; Smith, R.M.; Sivirichi, G.M. Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis. Water 2016, 8, 116. https://doi.org/10.3390/w8040116
Newcomer Johnson TA, Kaushal SS, Mayer PM, Smith RM, Sivirichi GM. Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis. Water. 2016; 8(4):116. https://doi.org/10.3390/w8040116
Chicago/Turabian StyleNewcomer Johnson, Tamara A., Sujay S. Kaushal, Paul M. Mayer, Rose M. Smith, and Gwen M. Sivirichi. 2016. "Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis" Water 8, no. 4: 116. https://doi.org/10.3390/w8040116
APA StyleNewcomer Johnson, T. A., Kaushal, S. S., Mayer, P. M., Smith, R. M., & Sivirichi, G. M. (2016). Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis. Water, 8(4), 116. https://doi.org/10.3390/w8040116