
Species Composition Affects the Accuracy of Stand-Level Biomass Models in Hemiboreal Forests


Abstract
:1. Introduction
2. Materials and Methods
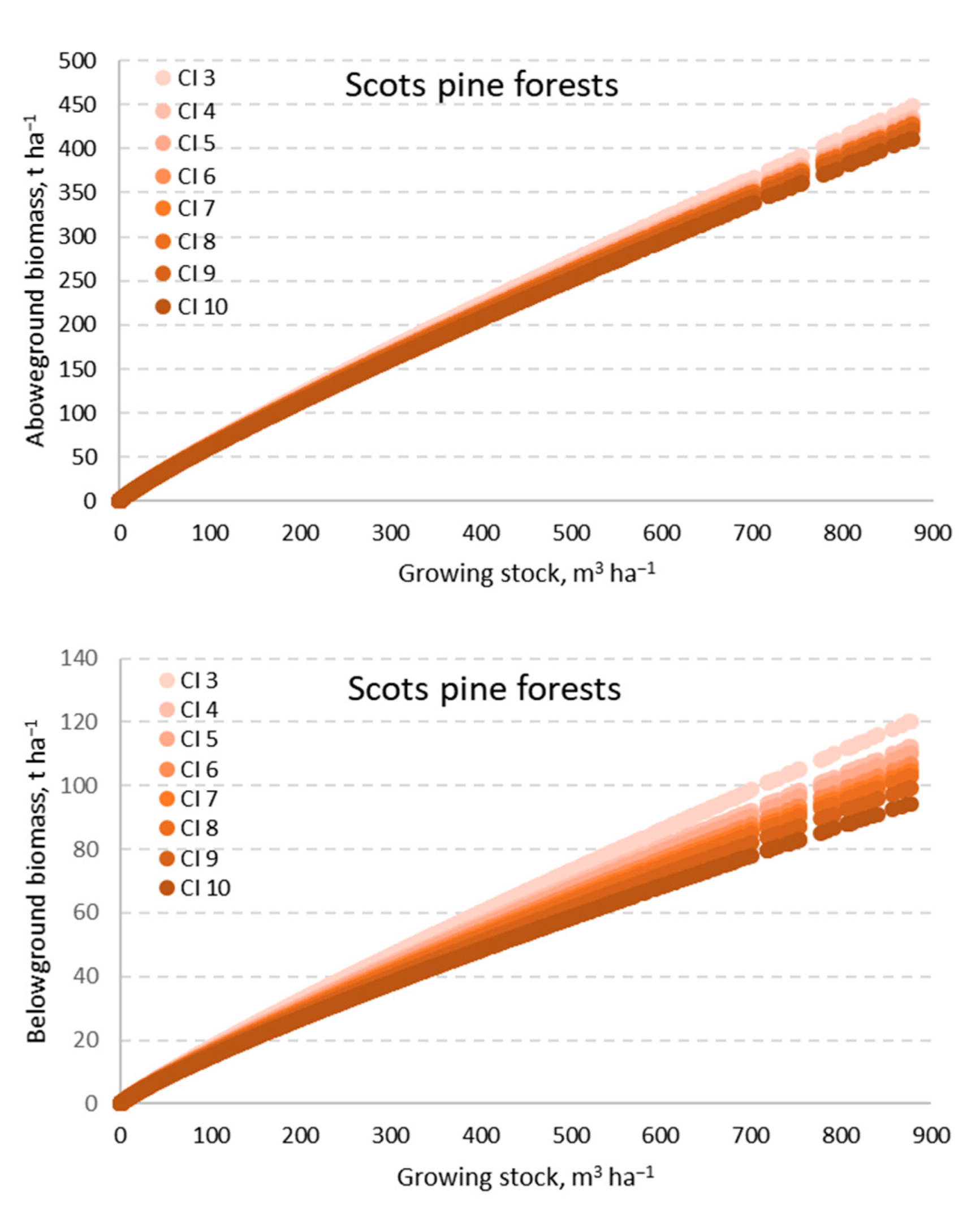
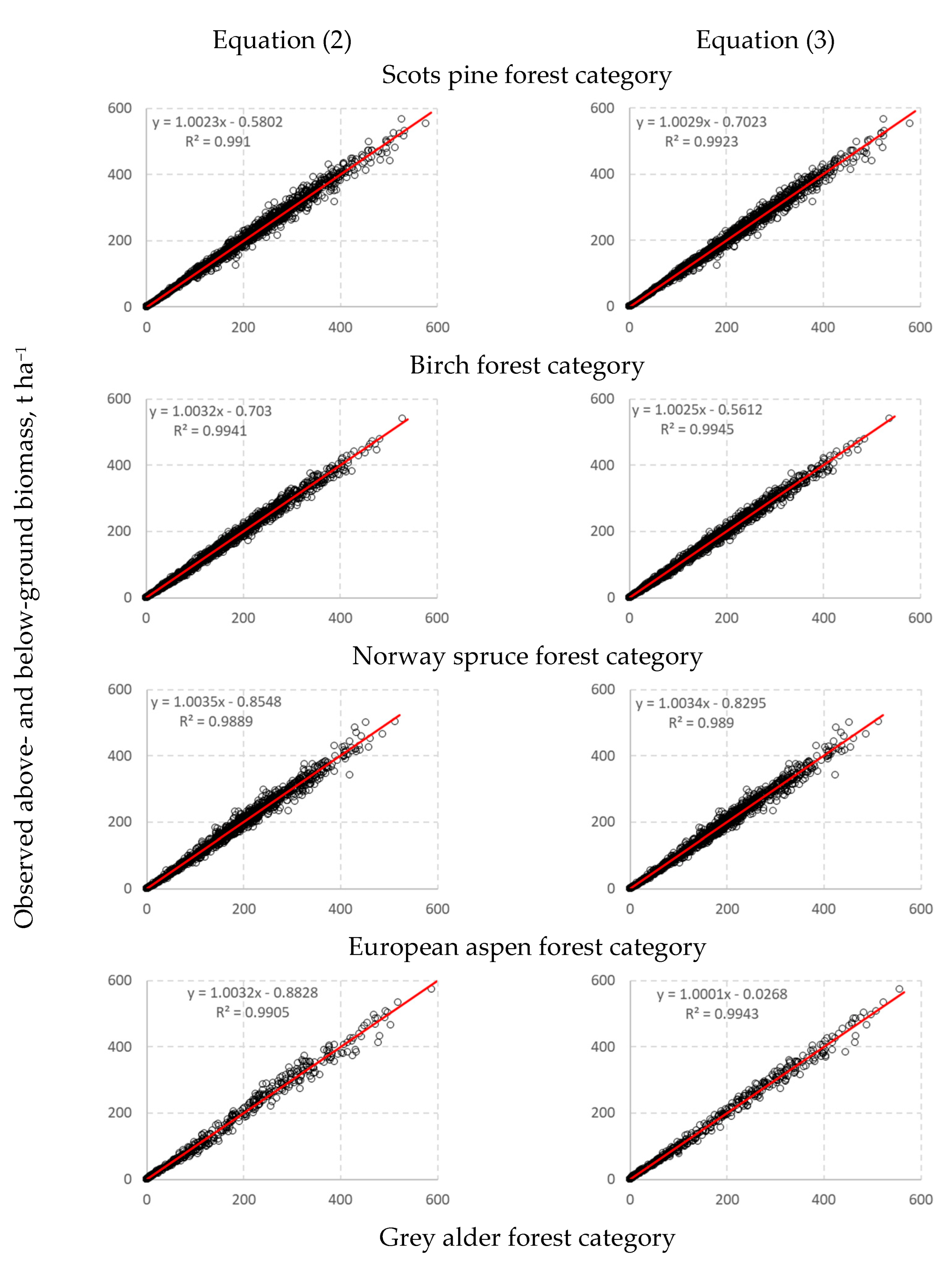
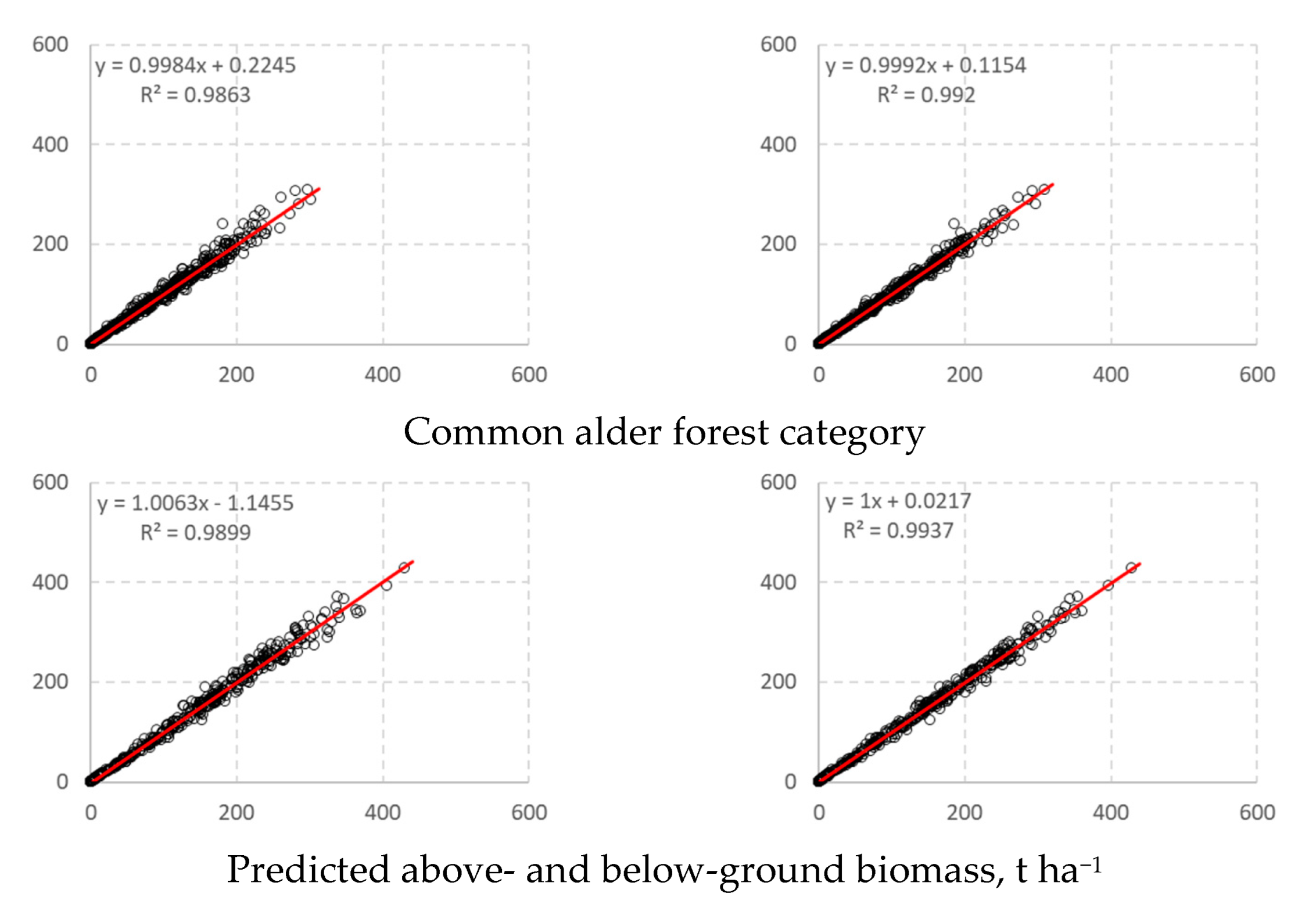
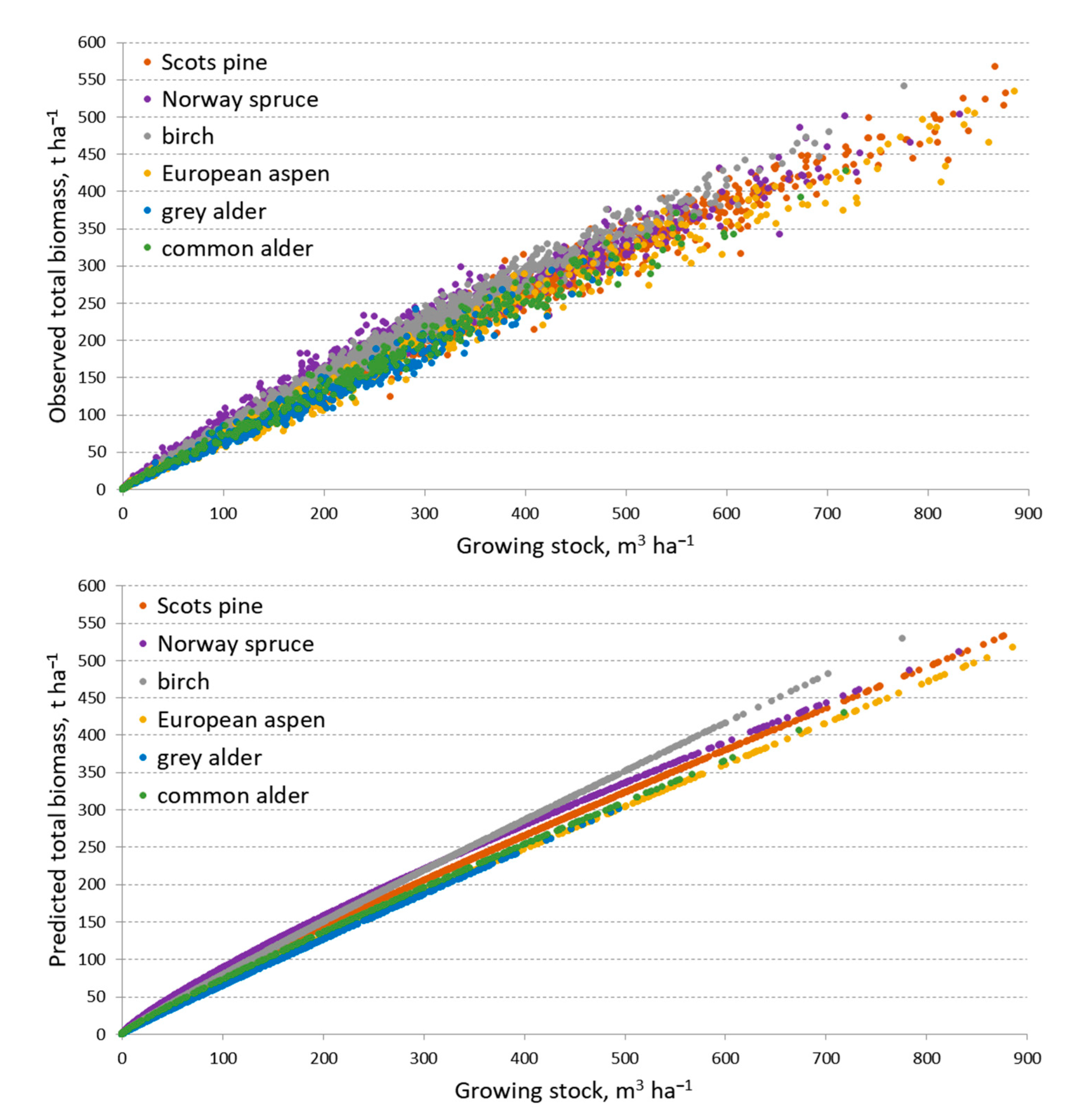
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.J.; Thivolle-Cazat, A.; Bronisz, K.; et al. Comparison of Carbon Estimation Methods for European Forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Martin, A.R.; Doraisami, M.; Thomas, S.C. Global Patterns in Wood Carbon Concentration across the World’s Trees and Forests. Nat. Geosci. 2018, 11, 915–920. [Google Scholar] [CrossRef]
- Lu, J.; Feng, Z.; Zhu, Y. Estimation of Forest Biomass and Carbon Storage in China Based on Forest Resources Inventory Data. Forests 2019, 10, 650. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Eskeland, G.S.; Giske, J.; Hansen, J.P. The Global Potential for Carbon Capture and Storage from Forestry. Carbon Balance Manag. 2016, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The Structure, Distribution, and Biomass of the World’s Forests. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef] [Green Version]
- Bārdule, A.; Petaja, G.; Butlers, A.; Purviņa, D.; Lazdiņš, A. Estimation of Litter Input in Hemiboreal Forests with Drained Organic Soils for Improvement of GHG Inventories. Balt. For. 2021, 29, 232–246. [Google Scholar]
- Gschwantner, T.; Alberdi, I.; Bauwens, S.; Bender, S.; Borota, D.; Bosela, M.; Bouriaud, O.; Breidenbach, J.; Donis, J.; Fischer, C.; et al. Growing Stock Monitoring by European National Forest Inventories: Historical Origins, Current Methods and Harmonisation. For. Ecol. Manag. 2022, 505, 119868. [Google Scholar] [CrossRef]
- Repo, A.; Rajala, T.; Henttonen, H.M.; Lehtonen, A.; Peltoniemi, M.; Heikkinen, J. Age-Dependence of Stand Biomass in Managed Boreal Forests Based on the Finnish National Forest Inventory Data. For. Ecol. Manag. 2021, 498, 119507. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Gómez-García, E.; Diéguez-Aranda, U.; Barrio-Anta, M.; Crecente-Campo, F. Aboveground Stand-Level Biomass Estimation: A Comparison of Two Methods for Major Forest Species in Northwest Spain. Ann. For. Sci. 2012, 69, 735–746. [Google Scholar] [CrossRef] [Green Version]
- Di Cosmo, L.; Gasparini, P.; Tabacchi, G. A National-Scale, Stand-Level Model to Predict Total above-Ground Tree Biomass from Growing Stock Volume. For. Ecol. Manag. 2016, 361, 269–276. [Google Scholar] [CrossRef]
- Fang, J.-Y.; Wang, G.G.; Liu, G.-H.; Xu, S.-L. Forest Biomass of China: An Estimate Based on the Biomass-Volume Relationship. Ecol. Appl. 1998, 8, 1084. [Google Scholar]
- Lehtonen, A.; Mäkipää, R.; Heikkinen, J.; Sievänen, R.; Liski, J. Biomass Expansion Factors (BEFs) for Scots Pine, Norway Spruce and Birch According to Stand Age for Boreal Forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Zasada, M.; Bronisz, K.; Bronisz, A.; Bijak, S. Biomass Conversion and Expansion Factors for a Chronosequence of Young Naturally Regenerated Silver Birch (Betula Pendula Roth) Stands Growing on Post-Agricultural Sites. For. Ecol. Manag. 2017, 384, 208–220. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Šebeň, V. Biomass Functions and Expansion Factors for Young Trees of European Ash and Sycamore Maple in the Inner Western Carpathians. Austrian J. For. Sci. 2015, 132, 1–26. [Google Scholar]
- Khan, M.N.I.; Islam, M.R.; Rahman, A.; Azad, M.S.; Mollick, A.S.; Kamruzzaman, M.; Sadath, M.N.; Feroz, S.M.; Rakkibu, M.G.; Knohl, A. Allometric Relationships of Stand Level Carbon Stocks to Basal Area, Tree Height and Wood Density of Nine Tree Species in Bangladesh. Glob. Ecol. Conserv. 2020, 22, e01025. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized Biomass and Leaf Area Allometric Equations for European Tree Species Incorporating Stand Structure, Tree Age and Climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Smith, A.; Granhus, A.; Astrup, R. Functions for Estimating Belowground and Whole Tree Biomass of Birch in Norway. Scand. J. For. Res. 2016, 31, 568–582. [Google Scholar] [CrossRef]
- Wirth, C.; Schumacher, J.; Schulze, E.D. Generic Biomass Functions for Norway Spruce in Central Europe-a Meta-Analysis Approach toward Prediction and Uncertainty Estimation. Tree Physiol. 2004, 24, 121–139. [Google Scholar] [CrossRef] [Green Version]
- Jalkanen, A.; Makipaa, R.; Stahl, G.; Lehtonen, A.; Petersson, H. Estimation of the Biomass Stock of Trees in Sweden: Comparison of Biomass Equations and Age-Dependent Biomass Expansion Factors. Ann. For. Sci. 2005, 62, 845–851. [Google Scholar] [CrossRef] [Green Version]
- Shi, L. Methods of Estimating Forest Biomass: A Review; Tumuluru, S.L.E.-J.S., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Liepiņš, J.; Liepiņš, K.; Lazdiņš, A. Equations for Estimating the Above- and Belowground Biomass of Grey Alder (Alnus Incana (L.) Moench.) and Common Alder (Alnus Glutinosa L.) in Latvia. Scand. J. For. Res. 2021, 36, 389–400. [Google Scholar] [CrossRef]
- Liepiņš, J.; Lazdiņš, A.; Liepiņš, K. Equations for Estimating Above- and Belowground Biomass of Norway Spruce, Scots Pine, Birch Spp. and European Aspen in Latvia. Scand. J. For. Res. 2018, 33, 58–70. [Google Scholar] [CrossRef]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation Zones and Their Sections in Northwestern Europe. Ann. Bot. Fenn. 1968, 3, 169–211. [Google Scholar]
- EEA. European Forest Types. Categories and Types for Sustainable Forest Management Reporting and Policy; EEA: Copenhagen, Denmark, 2007; Volume 3. [Google Scholar]
- Jansons, J.; Līcīte, I. Latvia. In National Forest Inventories: Pathways for Common Reporting; Erkki, T., Thomas, G., Mark Lawrence, R.E.M., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 341–350. [Google Scholar]
- Liepa, I. Pieauguma Mācība; LLU: Jelgava, Latvia, 1996. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 51. [Google Scholar] [CrossRef]
- Zālītis, P.; Jansons, J. Latvian Forest Typology and Its Origin; Saule: Salaspils, Latvia, 2013. [Google Scholar]
- Motulsky, H.; Arthur, C. Fitting Models to Biological Data Using Linear and Nonlinear Regression. A Practical Guide to Curve Fitting; GraphPad Software: San Diego, CA, USA, 2003. [Google Scholar]
- FAO. Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2020. [Google Scholar]
- Avitabile, V.; Camia, A. An Assessment of Forest Biomass Maps in Europe Using Harmonized National Statistics and Inventory Plots. For. Ecol. Manag. 2018, 409, 489–498. [Google Scholar] [CrossRef]
- Pajtik, J.; Konopka, B.; Lukac, M. Individual Biomass Factors for Beech, Oak and Pine in Slovakia: A Comparative Study in Young Naturally Regenerated Stands. Trees Struct. Funct. 2011, 25, 277–288. [Google Scholar] [CrossRef]
- Petersson, H.; Holm, S.; Ståhl, G.; Alger, D.; Fridman, J.; Lehtonen, A.; Lundström, A.; Mäkipää, R. Individual Tree Biomass Equations or Biomass Expansion Factors for Assessment of Carbon Stock Changes in Living Biomass-A Comparative Study. For. Ecol. Manag. 2012, 270, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Rytter, L.; Ingerslev, M.; Kilpeläinen, A.; Torssonen, P.; Lazdina, D.; Löf, M.; Madsen, P.; Muiste, P.; Stener, L.-G. Increased Forest Biomass Production in the Nordic and Baltic Countries-A Review on Current and Future Opportunities. Silva Fenn. 2016, 50, 1660. [Google Scholar] [CrossRef] [Green Version]
- Liepiņš, J.; Ivanovs, J.; Lazdiņš, A.; Jansons, J.; Liepiņš, K. Mapping of Basic Density within European Aspen Stems in Latvia. Silva Fenn. 2017, 51, 7798. [Google Scholar] [CrossRef] [Green Version]
- Liepiņš, J.; Liepiņš, K. Mean Basic Density and Its Axial Variation in Scots Pine, Norway Spruce and Birch Stems. Res. Rural Dev. 2017, 1, 21–27. [Google Scholar]
- Konôpka, B.; Pajtík, J.; Marušák, R. Canopy Closure Altered Biomass Allocation in Young Spruce Stand. J. For. Sci. 2015, 61, 62–71. [Google Scholar]
- Runģis, D.E.; Lībiete, Z.; Korica, A.; Katrevičs, J.; Jansons, Ā.; Veinberga, I.; Jansons, J. Genetic Diversity and Differentiation of Even-Aged Norway Spruce Stands in Latvia. Balt. For. 2019, 25, 45–51. [Google Scholar] [CrossRef]
- Bārdule, A.; Liepiņš, J.; Liepiņš, K.; Stola, J.; Butlers, A.; Lazdiņš, A. Variation in Carbon Content among the Major Tree Species in Hemiboreal Forests in Latvia. Forests 2021, 12, 1292. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
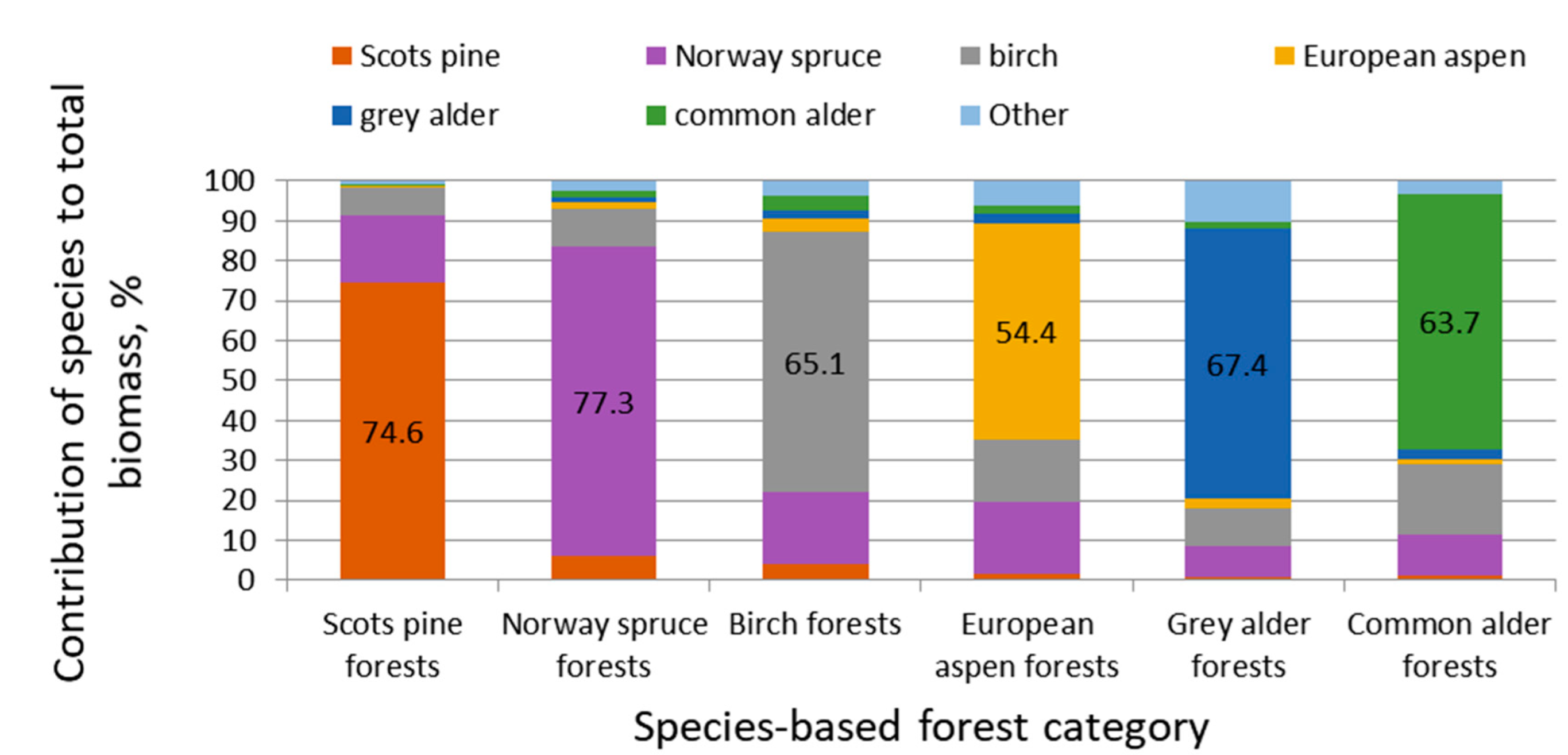{kind=link}
| Species-Based Forest Category | NFI Plots | GS, m3 ha−1 | AGB, t ha−1 | BGB, t ha−1 | SB, t ha−1 | BB, t ha−1 |
|---|---|---|---|---|---|---|
| Average (Min–Max) | Average (Min–Max) | Average (Min–Max) | Average (Min–Max) | Average (Min–Max) | ||
| Scots pine | 1838 | 265.8 (0.1–958.5) | 143.7 (0.1–456.4) | 35.2 (0.1–111.3) | 112.4 (0.1–403.6) | 28 (0.1–124.2) |
| Norway spruce | 1231 | 224.4 (0.1–832.2) | 128 (0.2–398.8) | 35.5 (0.1–112.6) | 89.9 (0.1–321.1) | 34.2 (0.2–96.2) |
| Birch | 1800 | 182.9 (0.1–776.4) | 105.5 (0.1–427.5) | 28.6 (0.1–114) | 84.3 (0.1–352.1) | 21 (0.1–91.9) |
| European aspen | 513 | 231.1 (0.1–1015.2) | 114.6 (0.1–483.9) | 28 (0.1–100.7) | 90.8 (0.1–404.2) | 21.3 (0.1–80.8) |
| Grey alder | 582 | 123.6 (0.1–494.3) | 61.7 (0.1–248.8) | 16.5 (0.1–63.3) | 50.4 (0.1–215.2) | 11.8 (0.1–75.4) |
| Common alder | 396 | 210.3 (0.1–1035.6) | 108.9 (0.1–494.8) | 28.1 (0.1–88) | 91.7 (0.1–484.6) | 17.2 (0.1–53.8) |
| Other | 170 | 173.4 (0.1–731.8) | 90.9 (0.1–319.1) | 30.3 (0.1–182.2) | 68.1 (0.1–238.8) | 32.4 (0.2–136.0) |
| Forest Category | Variable | Age | Diameter at Breast Height | Height | Basal Area | Growing Stock |
|---|---|---|---|---|---|---|
| Scots pine | AGB | 0.56 | 0.75 | 0.86 | 0.94 | 0.99 |
| BGB | 0.55 | 0.76 | 0.86 | 0.93 | 0.99 | |
| SB | 0.55 | 0.75 | 0.89 | 0.93 | 0.99 | |
| BB | 0.53 | 0.64 | 0.62 | 0.82 | 0.82 | |
| Norway spruce | AGB | 0.75 | 0.77 | 0.86 | 0.96 | 0.99 |
| BGB | 0.75 | 0.78 | 0.85 | 0.96 | 0.99 | |
| SB | 0.79 | 0.80 | 0.89 | 0.94 | 0.99 | |
| BB | 0.62 | 0.66 | 0.70 | 0.92 | 0.90 | |
| Birch | AGB | 0.82 | 0.83 | 0.91 | 0.95 | 0.99 |
| BGB | 0.83 | 0.85 | 0.90 | 0.94 | 0.99 | |
| SB | 0.81 | 0.82 | 0.91 | 0.95 | 0.99 | |
| BB | 0.83 | 0.84 | 0.84 | 0.86 | 0.93 | |
| European aspen | AGB | 0.93 | 0.90 | 0.94 | 0.96 | 0.99 |
| BGB | 0.92 | 0.90 | 0.93 | 0.95 | 0.99 | |
| SB | 0.92 | 0.89 | 0.94 | 0.96 | 0.99 | |
| BB | 0.92 | 0.91 | 0.91 | 0.91 | 0.96 | |
| Grey alder | AGB | 0.86 | 0.85 | 0.92 | 0.96 | 0.99 |
| BGB | 0.83 | 0.81 | 0.88 | 0.97 | 0.99 | |
| SB | 0.86 | 0.85 | 0.92 | 0.97 | 0.99 | |
| BB | 0.86 | 0.85 | 0.85 | 0.88 | 0.92 | |
| Common alder | AGB | 0.83 | 0.78 | 0.89 | 0.96 | 0.99 |
| BGB | 0.79 | 0.73 | 0.84 | 0.96 | 0.98 | |
| SB | 0.82 | 0.77 | 0.90 | 0.96 | 0.99 | |
| BB | 0.77 | 0.77 | 0.74 | 0.81 | 0.85 |
| Forest Category | Component * | Parameter Values ± Standard Errors | AIC | RMSE | MAPE | adjR2 | |||
|---|---|---|---|---|---|---|---|---|---|
| a | b1 | ||||||||
| Scots pine | AGB | 1.036 | 0.016 | 0.889 | 0.003 | 12,837.5 | 8.2 | 6.0 | 0.992 |
| BGB | 0.248 | 0.006 | 0.893 | 0.004 | 9473.8 | 3.2 | 8.0 | 0.981 | |
| SB | 0.375 | 0.008 | 1.021 | 0.003 | 13,022.5 | 8.3 | 9.0 | 0.989 | |
| BB | 1.685 | 0.111 | 0.517 | 0.011 | 12,837.5 | 7.9 | 29.7 | 0.703 | |
| Norway spruce | AGB | 1.428 | 0.031 | 0.840 | 0.004 | 8791.9 | 8.6 | 9.48 | 0.991 |
| BGB | 0.553 | 0.018 | 0.782 | 0.006 | 6766.5 | 3.8 | 17.4 | 0.976 | |
| SB | 0.293 | 0.006 | 1.054 | 0.003 | 7709.9 | 5.5 | 8.4 | 0.994 | |
| BB | 2.895 | 0.167 | 0.480 | 0.010 | 8505.2 | 7.6 | 26.7 | 0.845 | |
| Birch | AGB | 0.787 | 0.011 | 0.945 | 0.002 | 11,816.1 | 6.4 | 10.3 | 0.995 |
| BGB | 0.322 | 0.009 | 0.871 | 0.005 | 9537.3 | 3.4 | 19.9 | 0.977 | |
| SB | 0.522 | 0.010 | 0.978 | 0.003 | 12,135.7 | 7.0 | 10.2 | 0.990 | |
| BB | 0.503 | 0.038 | 0.734 | 0.013 | 12,283.0 | 7.3 | 35.9 | 0.802 | |
| European aspen | AGB | 0.644 | 0.026 | 0.957 | 0.006 | 3850.2 | 10.3 | 18.6 | 0.992 |
| BGB | 0.354 | 0.023 | 0.821 | 0.011 | 2984.8 | 4.4 | 21.8 | 0.955 | |
| SB | 0.489 | 0.024 | 0.964 | 0.008 | 3823.7 | 10.0 | 16.1 | 0.988 | |
| BB | 0.396 | 0.052 | 0.758 | 0.021 | 3473.3 | 7.1 | 39.8 | 0.878 | |
| Grey alder | AGB | 0.502 | 0.020 | 0.999 | 0.007 | 3754.8 | 6.1 | 17.0 | 0.989 |
| BGB | 0.351 | 0.025 | 0.816 | 0.013 | 2966.3 | 3.1 | 15.4 | 0.971 | |
| SB | 0.334 | 0.010 | 1.037 | 0.005 | 3161.1 | 3.6 | 16.8 | 0.994 | |
| BB | 0.299 | 0.057 | 0.782 | 0.035 | 3769.6 | 6.1 | 45.8 | 0.709 | |
| Common alder | AGB | 0.701 | 0.024 | 0.947 | 0.006 | 2663.4 | 6.9 | 9.7 | 0.993 |
| BGB | 0.675 | 0.050 | 0.715 | 0.013 | 2247.7 | 4.1 | 24.0 | 0.952 | |
| SB | 0.322 | 0.010 | 1.053 | 0.005 | 2411.7 | 5.0 | 11.1 | 0.996 | |
| BB | 1.081 | 0.218 | 0.543 | 0.035 | 2741.8 | 7.7 | 58.4 | 0.643 | |
| Forest Category | Component * | Parameter Values ± Standard Errors | AIC | RMSE | MAPE | adjR2 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b1 | b2 | |||||||||
| Scots pine | AGB | 1.187 | 0.022 | 0.882 | 0.003 | −0.048 | 0.004 | 12,802.2 | 7.9 | 6.0 | 0.993 |
| BGB | 0.392 | 0.009 | 0.870 | 0.003 | −0.161 | 0.005 | 8578.1 | 2.5 | 7.0 | 0.988 | |
| SB | 0.344 | 0.009 | 1.025 | 0.003 | 0.030 | 0.005 | 12,986.7 | 8.3 | 8.9 | 0.989 | |
| BB | 4.330 | 0.323 | 0.477 | 0.010 | −0.352 | 0.017 | 12,477.5 | 7.2 | 30.1 | 0.756 | |
| Norway spruce | AGB | 1.364 | 0.035 | 0.841 | 0.004 | 0.019 | 0.006 | 8783.7 | 8.5 | 9.4 | 0.991 |
| BGB | 0.477 | 0.019 | 0.786 | 0.006 | 0.062 | 0.010 | 6727.1 | 3.7 | 17.1 | 0.977 | |
| SB | 0.356 | 0.008 | 1.048 | 0.003 | −0.078 | 0.005 | 7476.3 | 5.0 | 7.8 | 0.995 | |
| BB | 1.583 | 0.117 | 0.491 | 0.009 | 0.268 | 0.021 | 8352.5 | 7.2 | 25.1 | 0.863 | |
| Birch | AGB | 0.677 | 0.012 | 0.956 | 0.002 | 0.049 | 0.004 | 11,640.5 | 6.1 | 10.3 | 0.995 |
| BGB | 0.314 | 0.011 | 0.873 | 0.005 | 0.009 | 0.007 | 9537.9 | 3.4 | 19.7 | 0.977 | |
| SB | 0.339 | 0.007 | 1.010 | 0.003 | 0.141 | 0.004 | 11,147.9 | 5.3 | 8.9 | 0.994 | |
| BB | 1.132 | 0.103 | 0.679 | 0.013 | −0.276 | 0.020 | 12,114.8 | 7.0 | 36.9 | 0.820 | |
| European aspen | AGB | 0.710 | 0.025 | 0.971 | 0.006 | −0.104 | 0.008 | 3692.3 | 8.8 | 18.2 | 0.996 |
| BGB | 0.475 | 0.023 | 0.848 | 0.008 | −0.261 | 0.011 | 2587.5 | 3.0 | 19.8 | 0.987 | |
| SB | 0.512 | 0.025 | 0.971 | 0.008 | −0.050 | 0.011 | 3803.7 | 9.8 | 16.2 | 0.988 | |
| BB | 0.634 | 0.080 | 0.780 | 0.020 | −0.344 | 0.029 | 3351.7 | 6.3 | 32.5 | 0.904 | |
| Grey alder | AGB | 0.693 | 0.026 | 0.986 | 0.006 | −0.131 | 0.007 | 3512.9 | 4.9 | 16.0 | 0.994 |
| BGB | 0.641 | 0.042 | 0.798 | 0.010 | −0.262 | 0.015 | 2718.7 | 2.5 | 15.4 | 0.969 | |
| SB | 0.355 | 0.012 | 1.035 | 0.005 | −0.025 | 0.007 | 3150.1 | 3.6 | 16.6 | 0.994 | |
| BB | 1.175 | 0.223 | 0.738 | 0.030 | −0.588 | 0.043 | 3611.8 | 5.3 | 43.2 | 0.778 | |
| Common alder | AGB | 0.748 | 0.021 | 0.972 | 0.005 | −0.111 | 0.007 | 2460.6 | 5.4 | 9.7 | 0.993 |
| BGB | 0.811 | 0.056 | 0.746 | 0.012 | −0.194 | 0.018 | 2146.4 | 3.6 | 16.4 | 0.965 | |
| SB | 0.323 | 0.010 | 1.054 | 0.005 | −0.007 | 0.008 | 2412.2 | 5.0 | 11.3 | 0.996 | |
| BB | 2.226 | 0.441 | 0.605 | 0.034 | −0.580 | 0.050 | 2639.9 | 6.7 | 39.0 | 0.725 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liepiņš, J.; Lazdiņš, A.; Kalēja, S.; Liepiņš, K. Species Composition Affects the Accuracy of Stand-Level Biomass Models in Hemiboreal Forests. Land 2022, 11, 1108. https://doi.org/10.3390/land11071108
Liepiņš J, Lazdiņš A, Kalēja S, Liepiņš K. Species Composition Affects the Accuracy of Stand-Level Biomass Models in Hemiboreal Forests. Land. 2022; 11(7):1108. https://doi.org/10.3390/land11071108
Chicago/Turabian StyleLiepiņš, Jānis, Andis Lazdiņš, Santa Kalēja, and Kaspars Liepiņš. 2022. "Species Composition Affects the Accuracy of Stand-Level Biomass Models in Hemiboreal Forests" Land 11, no. 7: 1108. https://doi.org/10.3390/land11071108
APA StyleLiepiņš, J., Lazdiņš, A., Kalēja, S., & Liepiņš, K. (2022). Species Composition Affects the Accuracy of Stand-Level Biomass Models in Hemiboreal Forests. Land, 11(7), 1108. https://doi.org/10.3390/land11071108