
Could Purposefully Engineered Native Grassland Gardens Enhance Urban Insect Biodiversity?

 , and
, and 
Abstract
:1. Introduction
1.1. Literature Review
1.2. Local Perspective on Urban Biodiversity
1.3. Objectives
- i.
- What is the plant species richness, plant species composition, and site heterogeneity (measured through beta diversity) of the intervention sites (young and native) in comparison to the control (young and non-native) and reference sites (old and native, and old and non-native)?
- ii.
- What differences in ground-dwelling insect species assemblages can be observed between these different sites?
- iii.
- What correlations could be drawn between plant species characteristics and observed insect species diversity?
2. Method
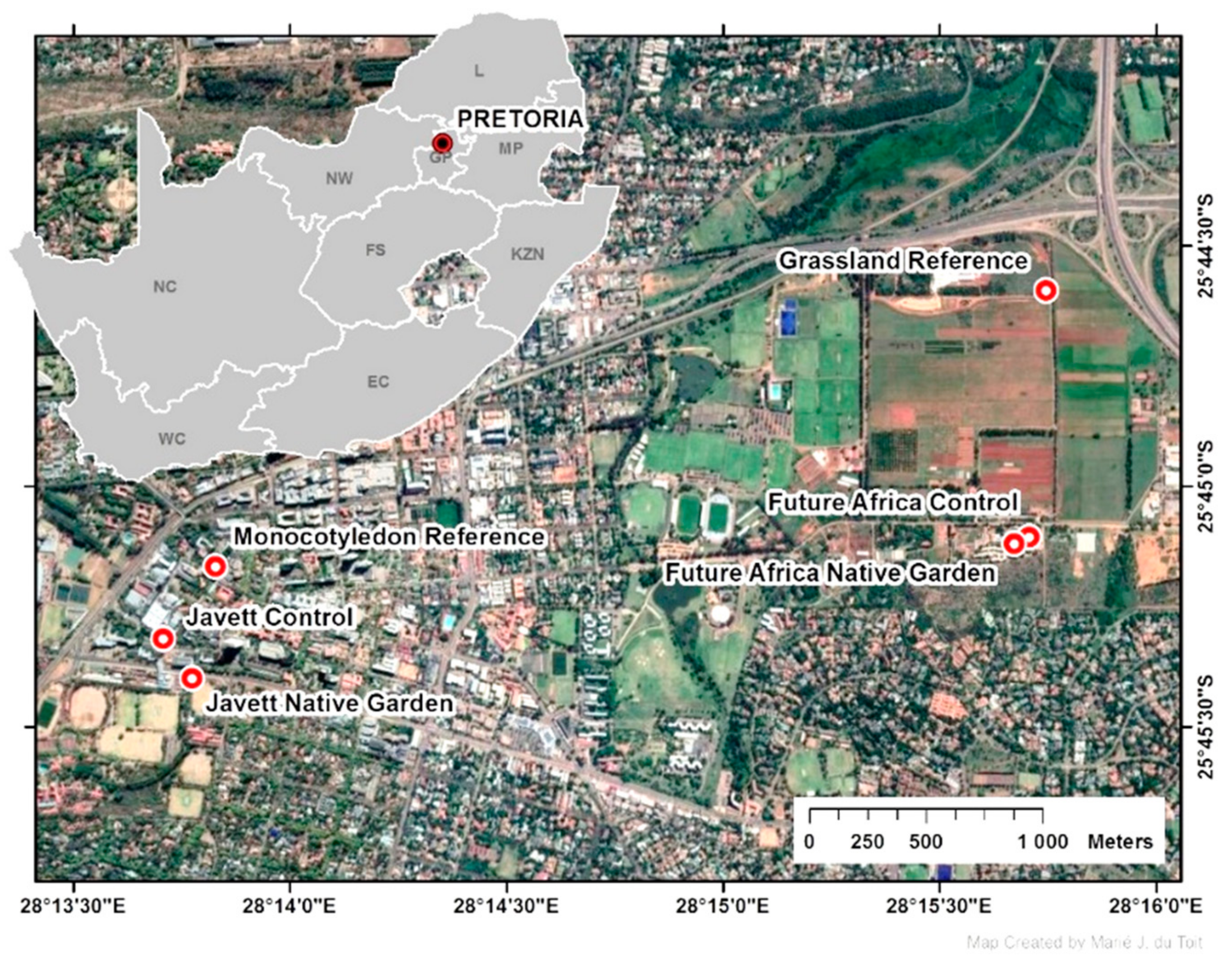
2.1. Study Area and Study Sites
2.2. Plant Species Composition and Species Diversity
2.3. Insect Diversity and Sampling
2.4. Plant and Insect Sampling Correlations
3. Results
3.1. Plant Assemblages and Dominant Garden Type Qualities
3.1.1. Plant Species, Family and Functional Type Richness
3.1.2. Plant Species Composition and Cover
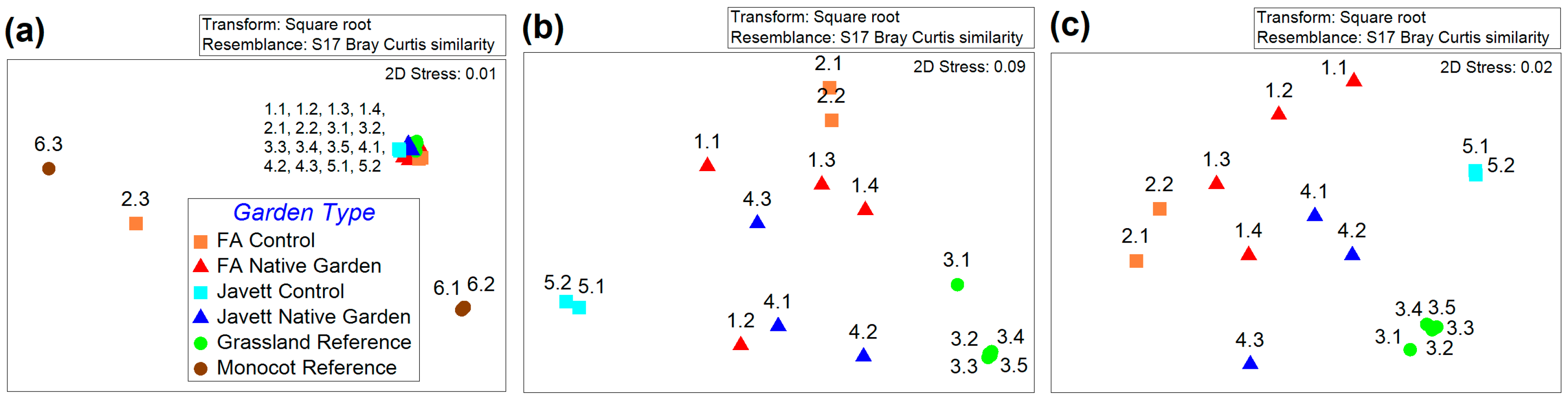
3.1.3. Plant Beta Diversity (Species Turnover)
3.2. Insect Diversity and Abundance
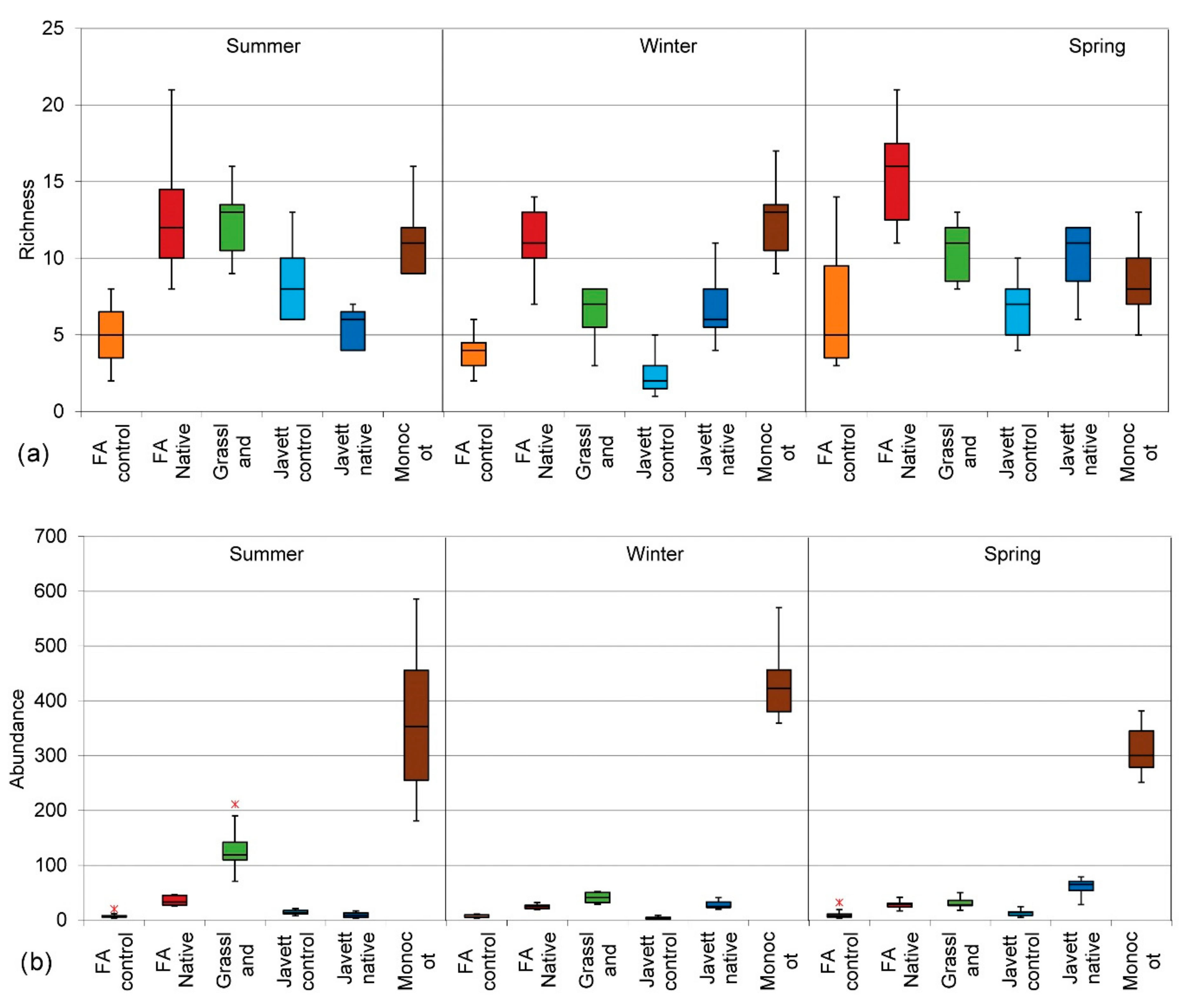
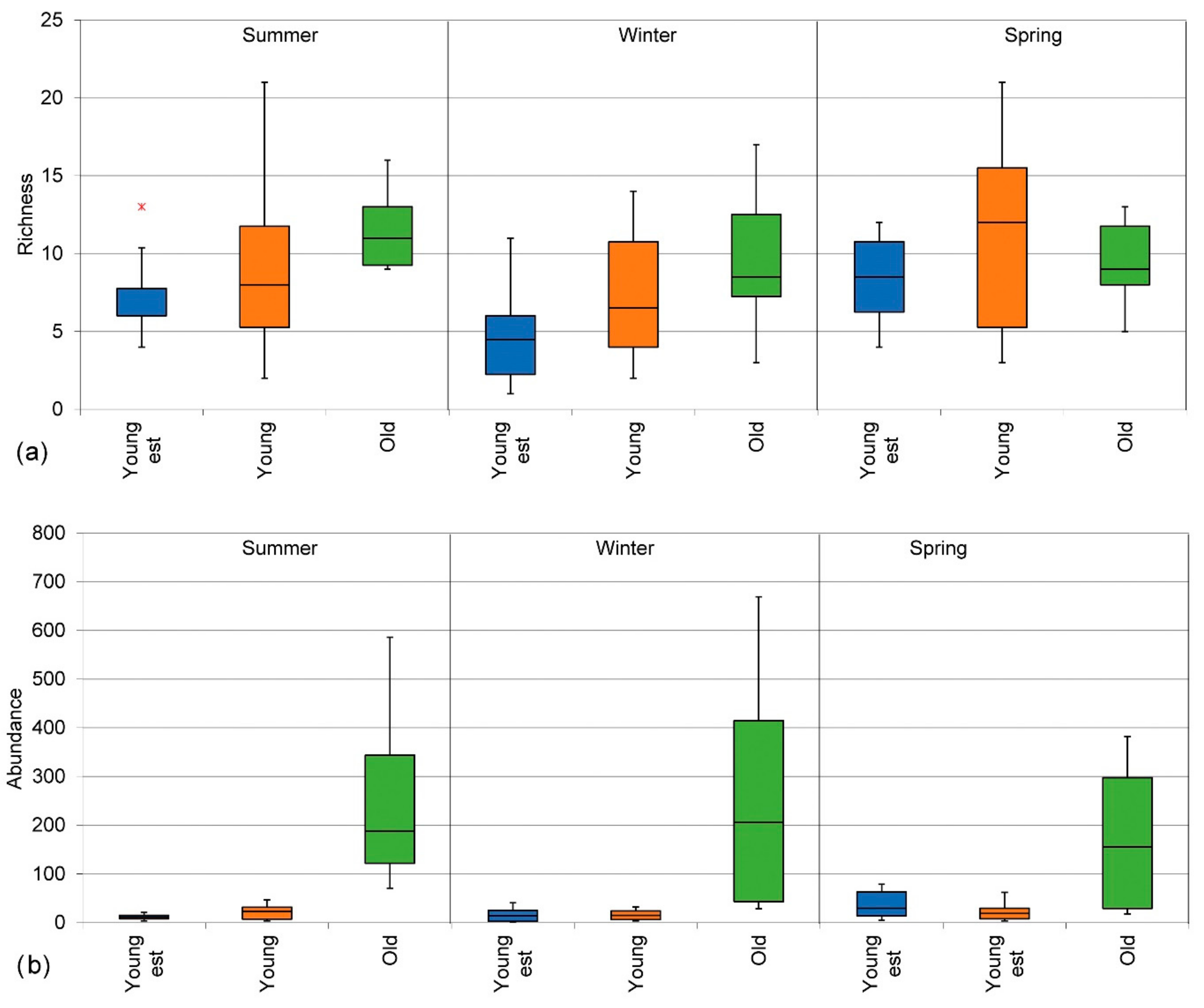
3.2.1. Insect Species Richness
3.2.2. Insect Species Abundance
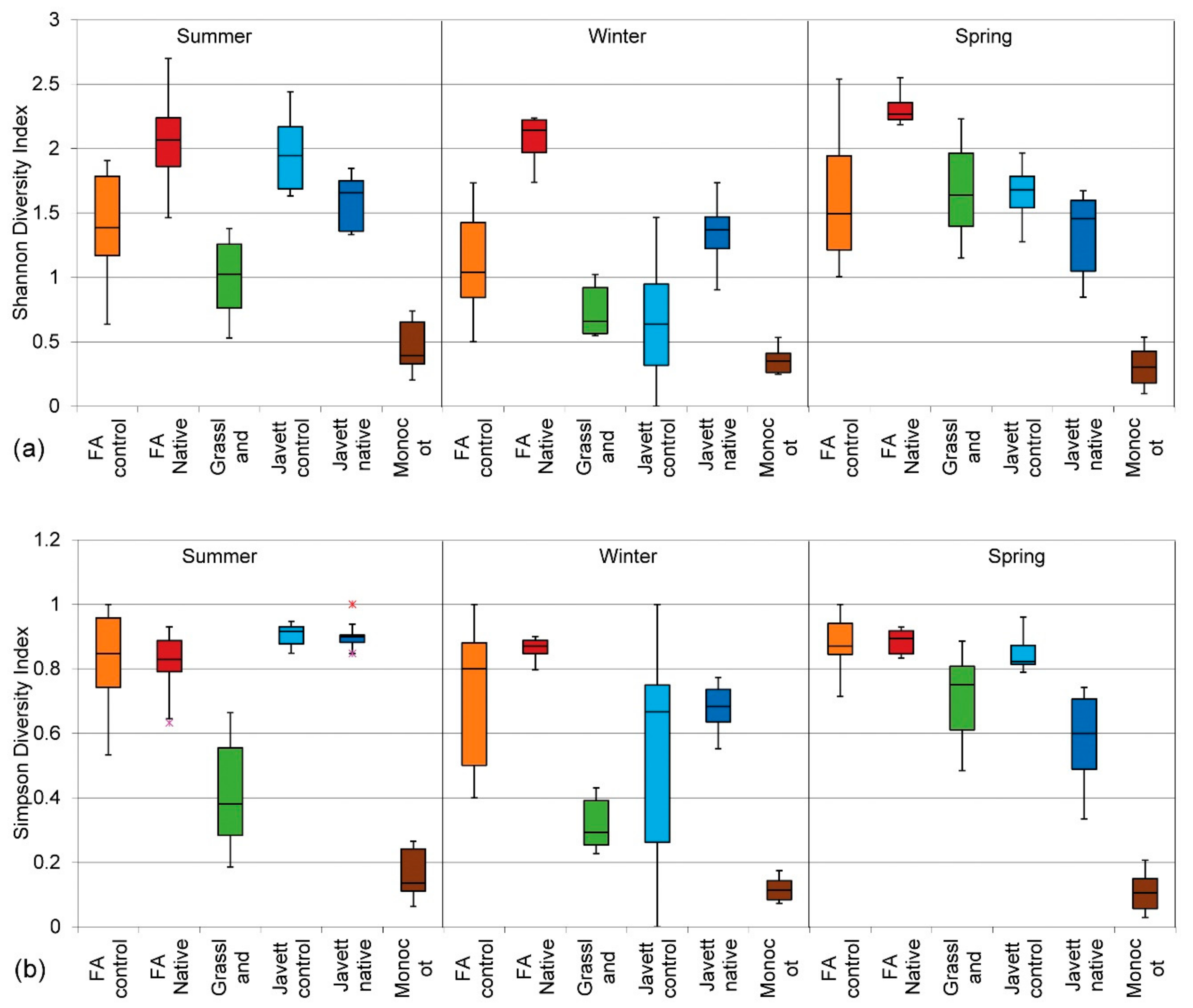
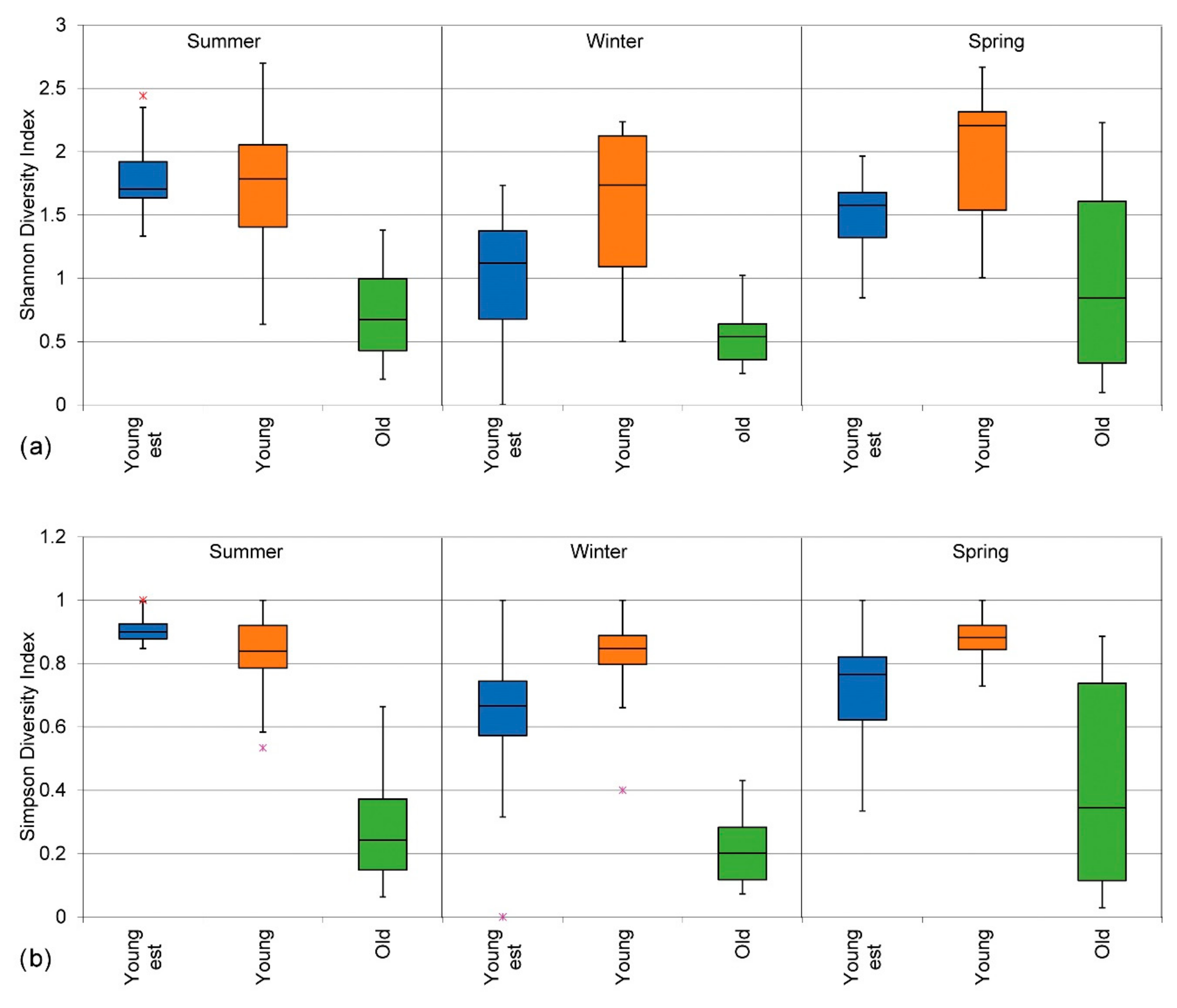
3.2.3. Shannon Diversity Index (H)
3.2.4. Simpson Diversity Index (D)
3.2.5. Effect of Patch Age
3.3. Plant and Insect Sampling Correlations
4. Discussion
4.1. Plant Beta Diversity (Species Turnover)
4.2. Plant Species Composition and Functional Types
4.3. Garden Microclimate and Age
4.4. Garden Size and Connectivity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ warning to humanity on insect extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Müller, N.; Werner, P.; Kelcey, J.G. (Eds.) Urban Biodiversity and Design; Wiley and Blackwell: Oxford, UK, 2010. [Google Scholar]
- Ahern, J.; Cilliers, S.S.; Niemelä, J. The concept of ecosystem services in adaptive urban planning and design: A framework for supporting innovation. Landsc. Urban Plan. 2014, 125, 254–259. [Google Scholar] [CrossRef] [Green Version]
- McEwan, K.; Ferguson, F.J.; Richardson, M.; Cameron, R. The good things in urban nature: A thematic framework for optimising urban planning for nature connectedness. Landsc. Urban Plan. 2020, 194, 103687. [Google Scholar] [CrossRef]
- Samways, M.J.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Solutions for humanity on how to conserve insects. Biol. Conserv. 2020, 242, 108427. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2006. [Google Scholar]
- Derby Lewis, A.; Bouman, M.J.; Winter, A.M.; Hasle, E.A.; Stotz, D.F.; Johnston, M.K.; Klinger, K.R.; Rosenthal, A.; Czarnecki, C.A. Does nature need cities? Pollinators reveal a role for cities in wildlife conservation. Front. Ecol. Evol. 2019, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shwartz, A.; Turbé, A.; Julliard, R.; Simon, L.; Prévot, A.C. Outstanding challenges for urban conservation research and action. Glob. Environ. Chang. 2014, 28, 39–49. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Hahs, A.K. The future of urban biodiversity research: Moving beyond the ‘low-hanging fruit’. Urban Ecosyst. 2013, 16, 397–409. [Google Scholar] [CrossRef]
- McGeoch, M.A. The selection, testing and application of terrestrial insects as bioindi-cators. Biol. Rev. Camb. Philos. Soc. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Helden, A.J.; Stamp, G.C.; Leather, S.R. Urban biodiversity: Comparison of insect assemblages on native and non-native trees. Urban Ecosyst. 2012, 15, 611–624. [Google Scholar] [CrossRef]
- Weisser, W.W.; Siemann, E. Insects and Ecosystem Function; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Felson, A.J.; Bradford, M.A.; Terway, T.M. Promoting earth stewardship through urban design experiments. Front. Ecol. Environ. 2013, 11, 362–367. [Google Scholar] [CrossRef]
- Brunbjerg, A.K.; Hale, J.D.; Bates, A.J.; Fowler, R.E.; Rosenfeld, E.J.; Sadler, J.P. Can patterns of urban biodiversity be predicted using simple measures of green infrastructure? Urban For. Urban Green. 2018, 32, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Clark, T.E.; Samways, M.J. Sampling arthropod diversity for urban ecological landscaping in a species-rich southern hemisphere botanic garden. J. Insect Conserv. 1997, 1, 221–234. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Standish, R.J.; Hobbs, R.J.; Miller, J.R. Improving city life: Options for ecological restoration in urban landscapes and how these might influence interactions between people and nature. Landsc. Ecol. 2013, 28, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Millennium Ecosystem Assessment (MEA). Ecosystems and Human Wellbeing, Synthesis; Island Press: Washington, DC, USA, 2005.
- Knuff, A.K.; Staab, M.; Frey, J.; Helbach, J.; Klein, A.-M. Plant composition, not richness, drives occurrence of specialist herbivores. Ecol. Entomol. 2019, 44, 833–843. [Google Scholar] [CrossRef]
- Botha, M.; Siebert, S.J.; Van Den Berg, J. Do arthropod assemblages fit the grassland and savanna biomes of South Africa? S. Afr. J. Sci. 2016, 112, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Symstad, A.J.; Siemann, E.; Haarstad, J. An experimental test of the effect of plant functional group diversity on arthropod diversity. Oikos 2000, 89, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Pryke, J.S.; Roets, F.; Samways, M.J. Importance of habitat heterogeneity in remnant patches for conserving dung beetles. Biodivers. Conserv. 2013, 22, 2857–2873. [Google Scholar] [CrossRef]
- Collinge, S.K. Effects of grassland fragmentation on insect species loss, colonization, and movement patterns. Ecology 2000, 81, 2211–2226. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Norton, B.A.; Evans, K.L.; Warren, P.H. Urban biodiversity and landscape ecology: Patterns, processes and planning. Curr. Landsc. Ecol. Rep. 2016, 1, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Sattler, T.; Duelli, P.; Obrist, M.K.; Arlettaz, R.; Moretti, M. Response of arthropod species richness and functional groups to urban habitat structure and management. Landsc. Ecol. 2010, 25, 941–954. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef]
- Jaganmohan, M.; Vailshery, L.S.; Nagendra, H. Patterns of insect abundance and distribution in urban domestic gardens in Bangalore, India. Diversity 2013, 5, 767–778. [Google Scholar] [CrossRef]
- Salisbury, A.; Armitage, J.; Bostock, H.; Perry, J.; Tatchell, M.; Thompson, K. Enhancing gardens as habitats for flower-visiting aerial insects (pollinators): Should we plant native or exotic species? J. Appl. Ecol. 2015, 52, 1156–1164. [Google Scholar] [CrossRef]
- Conway, T.M.; Almas, A.D.; Coore, D. Ecosystem services, ecological integrity, and native species planting: How to balance these ideas in urban forest management? Urban For. Urban Green. 2019, 41, 1–5. [Google Scholar] [CrossRef]
- McHarg, I.L. Design with Nature; Wiley: New York, NY, USA, 1969. [Google Scholar]
- Kendle, A.D.; Rose, J.E. The aliens have landed! What are the justifications for ‘native only’ policies in landscape plantings? Landsc. Urban Plan. 2000, 47, 19–31. [Google Scholar] [CrossRef]
- Özgüner, H.; Kendle, A.D.; Bisgrove, R.J. Attitudes of landscape professionals towards naturalistic versus formal urban landscapes in the UK. Landsc. Urban Plan. 2007, 81, 34–45. [Google Scholar] [CrossRef]
- Zeunert, J. Challenging assumptions in urban restoration ecology. Landsc. J. 2013, 32, 231–242. [Google Scholar] [CrossRef]
- Lennon, M. Moral-material ontologies of nature conservation: Exploring the discord between ecological restoration and novel ecosystems. Environ. Values 2017, 26, 5–29. [Google Scholar] [CrossRef] [Green Version]
- Salisbury, A.; Al-Beidh, S.; Armitage, J.; Bird, S.; Bostock, H.; Platoni, A.; Tatchell, M.; Thompson, K.; Perry, J. Enhancing gardens as habitats for plant-associated invertebrates: Should we plant native or exotic species? Biodivers. Conserv. 2017, 26, 2657–2673. [Google Scholar] [CrossRef]
- Tallamy, D.W.; Shropshire, K.J. Ranking lepidopteran use of native versus introduced plants. Conserv. Biol. 2009, 23, 941–947. [Google Scholar] [CrossRef]
- Hoyle, H.; Hitchmough, J.; Jorgensen, A. Attractive, climate-adapted and sustainable? Public perception of non-native planting in the designed urban landscape. Landsc. Urban Plan. 2017, 164, 49–63. [Google Scholar] [CrossRef]
- Threlfall, C.; Kendal, D. The distinct ecological and social roles that wild spaces play in urban ecosystems. Urban For. Urban Green. 2018, 29, 348–356. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; Da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.; Pilgrim, J.D.; Rodrigues, A.S.L. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Pfab, M.F.; Compaan, P.C.; Whittington-Jones, C.A.; Engelbrecht, I.; Dumalisile, L.; Mills, L.; West, S.D.; Muller, P.; Masterson, G.P.R.; Nevhutalu, L.S.; et al. The Gauteng Conservation Plan: Planning for biodiversity in a rapidly urbanising province. Bothalia Afr. Biodivers. Conserv. 2017, 47, 1–16. [Google Scholar] [CrossRef] [Green Version]
- United Nations. World urbanization prospects: The 2018 Revision. United Nations, Department of Economic and Social Affairs, Population Division (UN/DESA/PD). 2018. Available online: https://population.un.org/wup/Download/ (accessed on 10 October 2020).
- Pauleit, S.; Lindley, S.; Cilliers, S.; Shackleton, C. Urbanisation and ecosystem services in sub-Saharan Africa: Current status and scenarios. Landsc. Urban Plan. 2018, 180, 247–248. [Google Scholar] [CrossRef]
- Breed, C.; Mehrtens, H. Using “live” public sector projects in design teaching to transform urban green infrastructure in South Africa. Land 2022, 11, 45. [Google Scholar] [CrossRef]
- Shackleton, C.M.; Gwedla, N. The legacy effects of colonial and apartheid imprints on urban greening in South Africa: Spaces, species and suitability. Front. Ecol. Evol. 2021, 8, 579813. [Google Scholar] [CrossRef]
- Shackleton, S.; Chinyimba, A.; Hebinck, P.; Shackleton, C.; Kaoma, H. Multiple benefits and values of trees in urban landscapes in two towns in northern South Africa. Landsc. Urban Plan. 2015, 136, 76–86. [Google Scholar] [CrossRef]
- Cilliers, S.; Siebert, S.J.; Du Toit, M.J.; Barthel, S.; Mishra, S.; Cornelius, A.; Davoren, E. Health clinic gardens as nodes of social-ecological innovation to promote garden ecosystem services in sub-Saharan Africa. Landsc. Urban Plan. 2018, 180, 294–307. [Google Scholar] [CrossRef]
- Cocks, M.; Alexander, J.; Mogano, L.; Vetter, S. Ways of belonging: Meanings of “nature” among Xhosa-speaking township residents in South Africa. J. Ethnobiol. 2016, 36, 820–841. [Google Scholar] [CrossRef]
- Makakavhule, K.; Landman, K. Towards deliberative democracy through democratic governance and design of public spaces in the South African capital city, Tshwane. Urban Des. Int. 2020, 25, 280–292. [Google Scholar] [CrossRef]
- Breed, C.A. Value negotiation and professional self-regulation—environmental concern in the design of the built environment. Urban For. Urban Green. 2022, 74, 127626. [Google Scholar] [CrossRef]
- Norton, B.A.; Thomson, L.J.; Williams, N.S.G.; McDonnell, M.J. The effect of urban ground covers on arthropods: An experiment. Urban Ecosyst. 2014, 17, 77–99. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Mata, L.; Mackie, J.A.; Hahs, A.K.; Stork, N.E.; Williams, N.S.G.; Livesley, S.J. Increasing biodiversity in urban green spaces through simple vegetation interventions. J. Appl. Ecol. 2017, 54, 1874–1883. [Google Scholar] [CrossRef] [Green Version]
- Van Schalkwyk, J.; Pryke, J.S.; Samways, M.J.; Gaigher, R. Congruence between arthropod and plant diversity in a biodiversity hotspot largely driven by underlying abiotic factors. Ecol. Appl. 2019, 29, e01883. [Google Scholar] [CrossRef]
- Mucina, L.; Hoare, D.B.; Lötter, M.C.; Du Preez, P.J.; Rutherford, M.C.; Scott-Shaw, C.R.; Kose, L. Grassland biome. In The Vegetation of South Africa, Lesotho and Swaziland; Mucina, L., Rutherford, M.C., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2006; Volume 19, pp. 348–437. [Google Scholar]
- Grobler, C.H.; Bredenkamp, G.J.; Brown, L.R. Primary grassland communities of urban open spaces in Gauteng, South Africa. S. Afr. J. Bot. 2006, 72, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Carbutt, C.; Tau, M.; Stephens, A.; Escott, B. The conservation status of temperate grasslands in southern Africa. Grassroots 2011, 1, 17–23. [Google Scholar]
- Koricheva, J.; Mulder, C.P.; Schmid, B.; Joshi, J.; Huss-Danell, K. Numerical responses of different trophic groups of invertebrates to manipulations of plant diversity in grasslands. Oecologia 2000, 125, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.R. Using ecological theory to guide urban planting design: An adaptation strategy for climate change. Landsc. J. 2011, 30, 2–11. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Vellend, M. Do commonly used indices of β-diversity measure species turnover? J. Veg. Sci. 2001, 12, 545–552. [Google Scholar] [CrossRef]
- Buschke, F.; Kemp, M.; Seaman, M.; Louw, S. Intra-annual variation of arthropod-plant interactions and arthropod trophic structure in an endangered grassland in the Free State province, South Africa. Afr. J. Range Forage Sci. 2011, 28, 57–63. [Google Scholar] [CrossRef]
- Vrdoljak, S.M.; Samways, M.J. Optimizing colored pan traps to survey flower visiting insects. J. Insect Conserv. 2012, 16, 345–354. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon-Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Kessler, M.; Abrahamczyk, S.; Bos, M.; Buchori, D.; Putra, D.D.; Gradstein, S.R.; Höhn, P.; Kluge, J.; Orend, F.; Pitopang, R.; et al. Alpha and beta diversity of plants and animals along a tropical land-use gradient. Ecol. Appl. 2009, 19, 2142–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moolman, J.; Van den Berg, J.; Conlong, D.; Cugala, D.; Siebert, S.; Le Ru, B. Species diversity and distribution of lepidopteran stem borers in South Africa and Mozambique. J. Appl. Entomol. 2013, 137, 641–720. [Google Scholar] [CrossRef]
- Davies-Colley, R.J.; Payne, G.; Van Elswijk, M. Microclimate gradients across a forest edge. N. Z. J. Ecol. 2000, 24, 111–121. [Google Scholar]
- Willmer, P.G. Microclimate and the environmental physiology of insects. In Advances in Insect Physiology; Berridge, M.J., Treherne, J.E., Wigglesworth, V.B., Eds.; Academic Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Cook, W.M.; Faeth, S.H. Irrigation and land-use drive ground arthropod community patterns in an urban desert. Env. Entomol. 2006, 35, 1532–1540. [Google Scholar] [CrossRef]
- Niemelä, J. Is there a need for a theory of urban ecology? Urban Ecosyst. 1999, 3, 57–65. [Google Scholar] [CrossRef]
- Rabitsch, W. The hitchhiker’s guide to alien ant invasions. BioControl 2011, 56, 551–572. [Google Scholar] [CrossRef]
- Sugihara, G.; Bersier, L.-F.; Southwood, T.R.E.; Primm, S.L.; May, R.M. Predicted correspondence between species abundances and dendrograms of niche similarities. Proc. Natl. Acad. Sci. USA 2003, 100, 5246–5251. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | Size (m2) | Perimeter (m) | Project Start | Intervention | Surroundings |
|---|---|---|---|---|---|
| Moot Plain Grassland (old and native) (reference area) | ±100 m2 sample area (part of a 5 ha grassland area) | Not applicable for sample area (buffer area 1100 m perimeter) | Land purchased by the University of Pretoria in 1938 | “Moot Plains Grassland” Little past disturbance—possibly grazing. More than 100 different species previously recorded. Recent rapid survey recorded 53 species. | Hillcrest Campus: 290 ha, of which 255 ha (92%) is green/pervious. Large motorway 500 m to the north and wetland area 1 km to the west. |
| Monocotyledon Garden (old and non-native) (reference area) | ±100 m2 (part of 0.25 ha green patch) | Not applicable for sample area (greater patch 203 m perimeter) | Established as a didactic garden in the late 1980s | Garden section with 14 mostly monocotyledon species. | Hatfield Campus: 570 ha, of which 188 ha (32%) is green/pervious. Larger garden patch includes lawn. |
| Future Africa Native Garden (young and native)(intervention) | 198 m2 | 60 m | October 2018 | Garden section with more than 70 different native species planted. | Hillcrest Campus: 290 ha, of which 255 ha (92%) is green/pervious. Natural rocky outcrop area 500 m to the south. |
| Future Africa Non-native Garden (young and non-native) (control areas) | 147 m2 + 37 m2 | 49 m + 30 m | October 2018 | Two garden sections with 12 species. | Same as above. |
| Javett Native Garden (young and native) (intervention) | 73 m2 | 55 m | February 2019 | Garden section with more than 40 different species planted. | Hatfield Campus: 570 ha, of which 188 ha (32%) is green/pervious. Large motorway 3 m to the north, a dominance of impervious surfaces. |
| Javett Non-native Garden (young and non-native) (control area) | 75 m2 | 40 m | February 2019 | Garden sections with 20 succulent species planted. | Hatfield Campus: 570 ha, of which 188 ha (32%) is green/pervious. A dominance of impervious surfaces, road and trees nearby. |
| Grassland Reference (May) | Grassland Reference (July) | Monocotyledon Reference (May) | Monocotyledon Reference (July) | Future Africa Native Garden (May) | Future Africa Native Garden (July) | Future Africa Control (May) | Future Africa Control (July) | Javett Native Garden (May) | Javett Native Garden (July) | Javett Control (May) | Javett Control (July) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 11 | 12 | 8 | 8 | 18 | 18 | 6 | 6 | 10 | 10 | 7 | 7 |
| Poaceae 79% Asteraceae 9% | Poaceae 78% | Strelitziaceae 33% Iridaceae 15% Agapanthaceae 11% | Strelitziaceae 25% Iridaceae 10% | Asteraceae 23% Poaceae 19% Hypoxidaceae 12% | Poaceae 19% Asteracea 11% | Asphodelaceae 31% Poaceae 21% Hypoxidaceae 13% | Asphodelaceae 29% Poaceae 19% Lamiaceae 12% | Asphodelaceae 19% Acanthaceae 15% Poaceae 13% | Asphodelaceae 19% Acanthaceae 16% Poaceae 16% | Aizoaceae 41% Asphodelaceae 20% Alliaceae 13% | Aizoaceae 40% Asphodelaceae 20% Alliaceae 11% |
| 25 | 27 | 9 | 9 | 32 | 32 | 8 | 8 | 19 | 19 | 14 | 14 |
| −0.60 | −0.32 | −0.72 (−0.17) ʸ | −0.73 (−0.20) ʸ | −0.87 | −0.85 | −0.81 (−0.43) ʸ | −0.87 (−0.60) ʸ | −0.76 | −0.76 | −0.27 | −0.15 |
| Themeda triandra 51% Eragrostis chloromelas 19% | Themeda triandra 50% Eragrostis chloromelas 19% | Strelitzia juncea 17% Strelitzia reginae 17% Aristea ecklonii 15% | Strelitzia juncea 13% Strelitzia reginae 13% Aristea ecklonii 10% | Haplocarpha lyrata 19% Melinis nerviglumis 18% Stachys natalensis var. natalensis 12% | Melinis nerviglumis 19% Haplocarpha lyrata 9% | Melinis nerviglumis 21% Aloiampelos tenuior 14% Hypoxis hemerocallidea 13% | Melinis nerviglumis 19% Aloe vera 13% Melissa officinalis 12% | Polygala virgata var. virgata 10% Dicliptrea eenii 9% | Dyschoriste setigera 9% Aloe merlothii 9% | Carprobrotus edulis 39% Nothoscordum gracile 13% Aloe arborescens (hybrid) 10% | Carprobrotus edulis 37% Nothoscordum gracile 11% Aloe arborescens (hybrid) 10% |
| 100% | 78% | 95% | 56% | 89% | 58% | 89% | 80% | 87% | 71% | 100% | 96.5% |
| 0% | 0% | 0% | 0% | 7% | 7% | 0% | 0% | 11% | 12% | 0% | 0% |
| 0% | 1% | 5% | 32% | 4% | 30% | 11% | 20% | 2% | 11% | 0% | 3.5% |
| 0% | 21% | 0% | 12% | 0% | 6% | 0% | 0% | 0% | 6% | 0% | 0% |
| 4% | 0% | 80% | 52% | 39% | 30% | 34% | 32% | 55% | 50% | 100% | 96.5% |
| 96% | 78% | 15% | 4% | 52% | 29% | 55% | 49% | 32% | 25% | 0% | 0% |
| 4 | 4 | 4 | 4 | 6 | 6 | 5 | 5 | 5 | 5 | 2 | 2 |
| Perennial graminoid (7) | Perennial graminoid (8) | Perennial forb (6) | Perennial forb (6) | Perennial forb (6) | Perennial graminoid (4) | Succulent (3) | Succulent (3) | Succulent (5) | Perennial graminoid (5) | Succulent (12) | Succulent (12) |
| 100% | 78% | 8% | 0% | 87.75% | 60% | 43% | 37% | 87% | 71% | 14% | 14% |
| 0% | 0% | 59% | 40% | 1.25% | 1% | 22% | 19% | 0% | 0% | 54% | 51% |
| 0% | 0% | 18% | 11% | 0% | 0% | 24% | 24% | 0% | 0% | 23% | 21% |
| Variables | Insect | Plant | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Abundance | Richness | Order | Ave_ Shannon Index | Ave_ Simpson Index | Ave_Beta Diversity_All | Ave_Beta Diversity_Removed | Family Richness (ƴ) | Species Richness (ƴ) | Functional Types (ƴ) | ||
| Summer | |||||||||||
| Plants summer | Ave_beta diversity_all | 0.3143 (0.5639) | −0.0286 (1.0000) | −0.3381 (0.4972) | −0.3143 (0.5639) | 0.1429 (0.8028) | - | - | - | - | - |
| Ave_beta diversity_ removed | 0.2571 (0.6583) | −0.3143 (0.5639) | −0.1690 (0.7139) | −0.6000 (0.2417) | −0.1429 (0.8028) | 0.6000 (0.2417) | - | - | - | - | |
| Family richness(ƴ) | 0.4857 (0.3556) | 0.8286 (0.0583) | −0.1690 (0.7139) | 0.2571 (0.6583) | −0.3714 (0.4972) | −0.2571 (0.6583) | −0.7143 (0.1361) | - | - | - | |
| Species richness(ƴ) | 0.3143 (0.5639) | 0.7714 (0.1028) | −0.1690 (0.7139) | 0.4857 (0.3556) | −0.0857 (0.9194) | −0.1429 (0.8028) | −0.7714 (0.1028) | 0.9429 * (0.0333) | - | - | |
| Functional types(ƴ) | −0.3237 (0.4972) | 0.0883 (0.9194) | 0.3482 (0.5639) | 0.3825 (0.4972) | −0.0883 (0.8028) | −0.9710 * (0.0028) | −0.7650 (0.1028) | 0.4119 (0.4972) | 0.324 (0.564) | - | |
| Ave% native cover | −0.0286 (1.0000) | 0.4286 (0.4194) | −0.3381 (0.4972) | 0.2571 (0.6583) | −0.0857 (0.9194) | −0.2571 (0.6583) | −0.8286 (0.0583) | 0.7143 (0.1361) | 0.771 (0.103) | 0.441 (0.419) | |
| Winter | |||||||||||
| Plants winter | Ave_beta diversity_all | 0.0857 (0.9194) | −0.2571 (0.6583) | 0.3395 (0.5639) | −0.6571 (0.1750) | −0.7143 (0.1361) | |||||
| Ave_beta diversity_ removed | −0.0286 (1.000) | −0.4857 (0.3556) | 0.1234 (0.9194) | −0.9429 * (0.0167) | −0.7714 (0.1028) | 0.7714 (0.1028) | |||||
| Family richness(ƴ) | 0.4286 (0.4194) | 0.7714 (0.1028) | 0.6172 (0.2417) | 0.4857 (0.3556) | 0.1429 (0.8028) | 0.0286 (1.0000) | −0.5429 (0.2972) | ||||
| Species richness(ƴ) | 0.1429 (0.8028) | 0.5429 (0.2972) | 0.4938 (0.3556) | 0.5429 (0.2972) | 0.2571 (0.6583) | 0.1429 (0.8028) | −0.4857 (0.3556) | 0.9429 * (0.0333) | |||
| Functional types(ƴ) | 0.0000 (1.0000) | 0.5002 (0.3556) | −0.1271 (0.8028) | 0.8827 (0.0583) | 0.7945 (0.1028) | −0.8827 * (0.0333) | −0.9710 * (0.0028) | 0.4119 (0.4972) | 0.3237 (0.5639) | ||
| Ave% native cover | 0.1429 (0.8028) | 0.1429 (0.8028) | −0.0309 (0.9194) | 0.6000 (0.2417) | 0.2571 (0.6583) | −0.0857 (0.9194) | −0.5429 (0.2972) | 0.6000 (0.2417) | 0.6571 (0.1750) | 0.3531 (0.5639) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breed, C.A.; Morelli, A.; Pirk, C.W.W.; Sole, C.L.; Du Toit, M.J.; Cilliers, S.S. Could Purposefully Engineered Native Grassland Gardens Enhance Urban Insect Biodiversity? Land 2022, 11, 1171. https://doi.org/10.3390/land11081171
Breed CA, Morelli A, Pirk CWW, Sole CL, Du Toit MJ, Cilliers SS. Could Purposefully Engineered Native Grassland Gardens Enhance Urban Insect Biodiversity? Land. 2022; 11(8):1171. https://doi.org/10.3390/land11081171
Chicago/Turabian StyleBreed, Christina A., Agata Morelli, Christian W. W. Pirk, Catherine L. Sole, Marié J. Du Toit, and Sarel S. Cilliers. 2022. "Could Purposefully Engineered Native Grassland Gardens Enhance Urban Insect Biodiversity?" Land 11, no. 8: 1171. https://doi.org/10.3390/land11081171
APA StyleBreed, C. A., Morelli, A., Pirk, C. W. W., Sole, C. L., Du Toit, M. J., & Cilliers, S. S. (2022). Could Purposefully Engineered Native Grassland Gardens Enhance Urban Insect Biodiversity? Land, 11(8), 1171. https://doi.org/10.3390/land11081171