Global and Regional Implications of Biome Evolution on the Hydrologic Cycle and Climate in the NCAR Dynamic Vegetation Model


Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussions
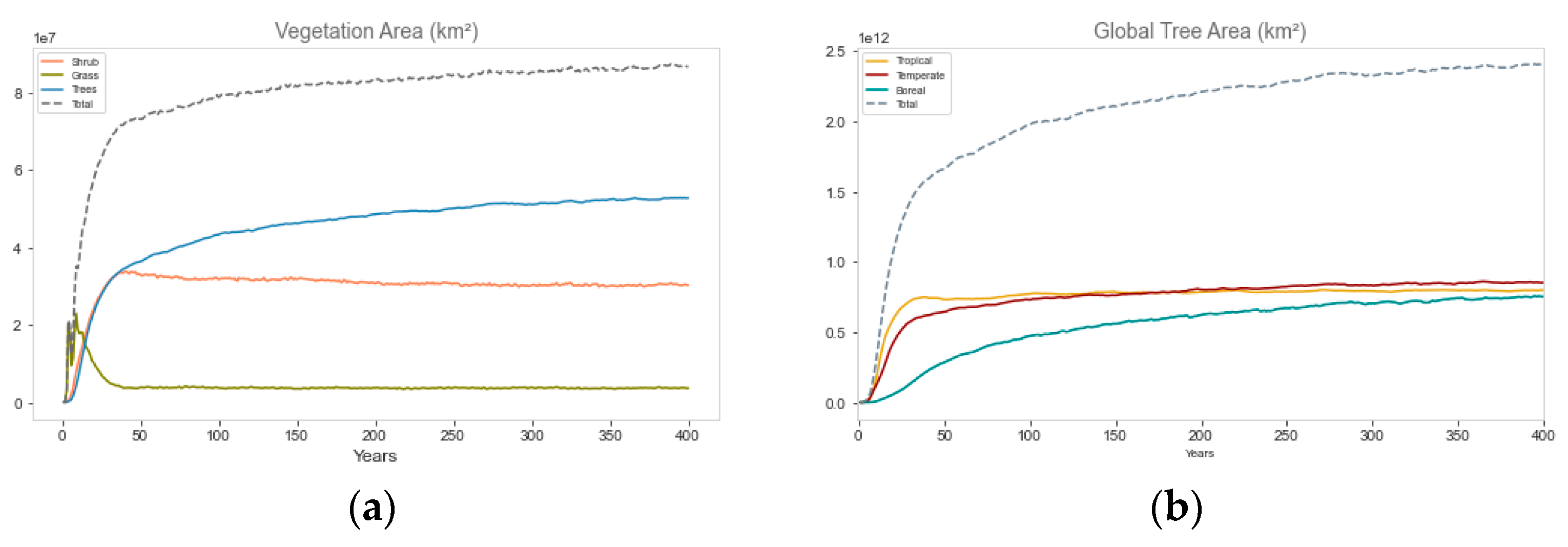
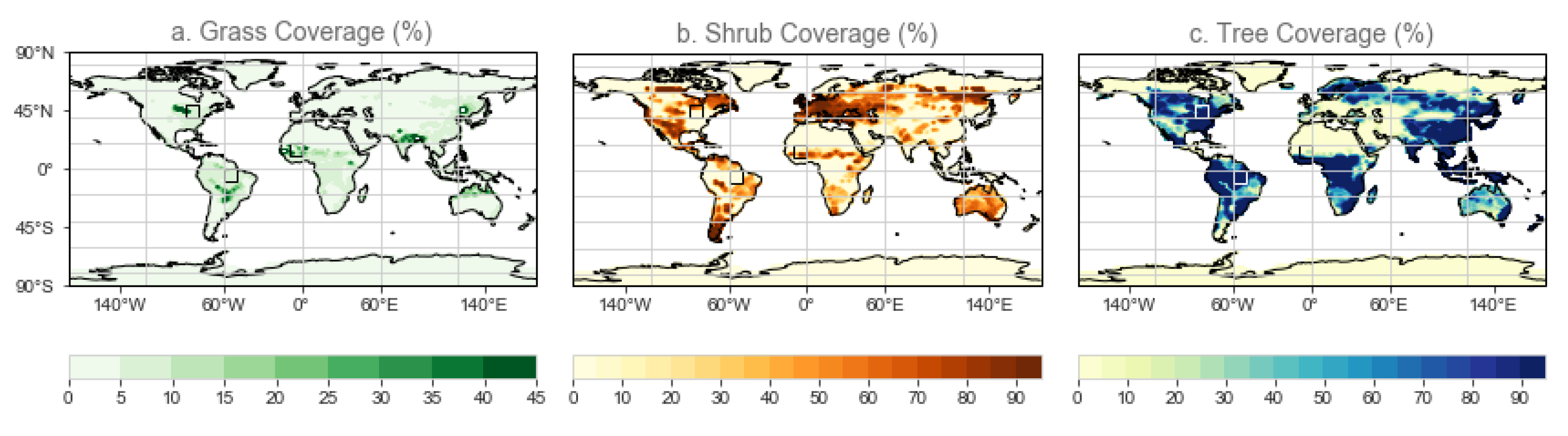
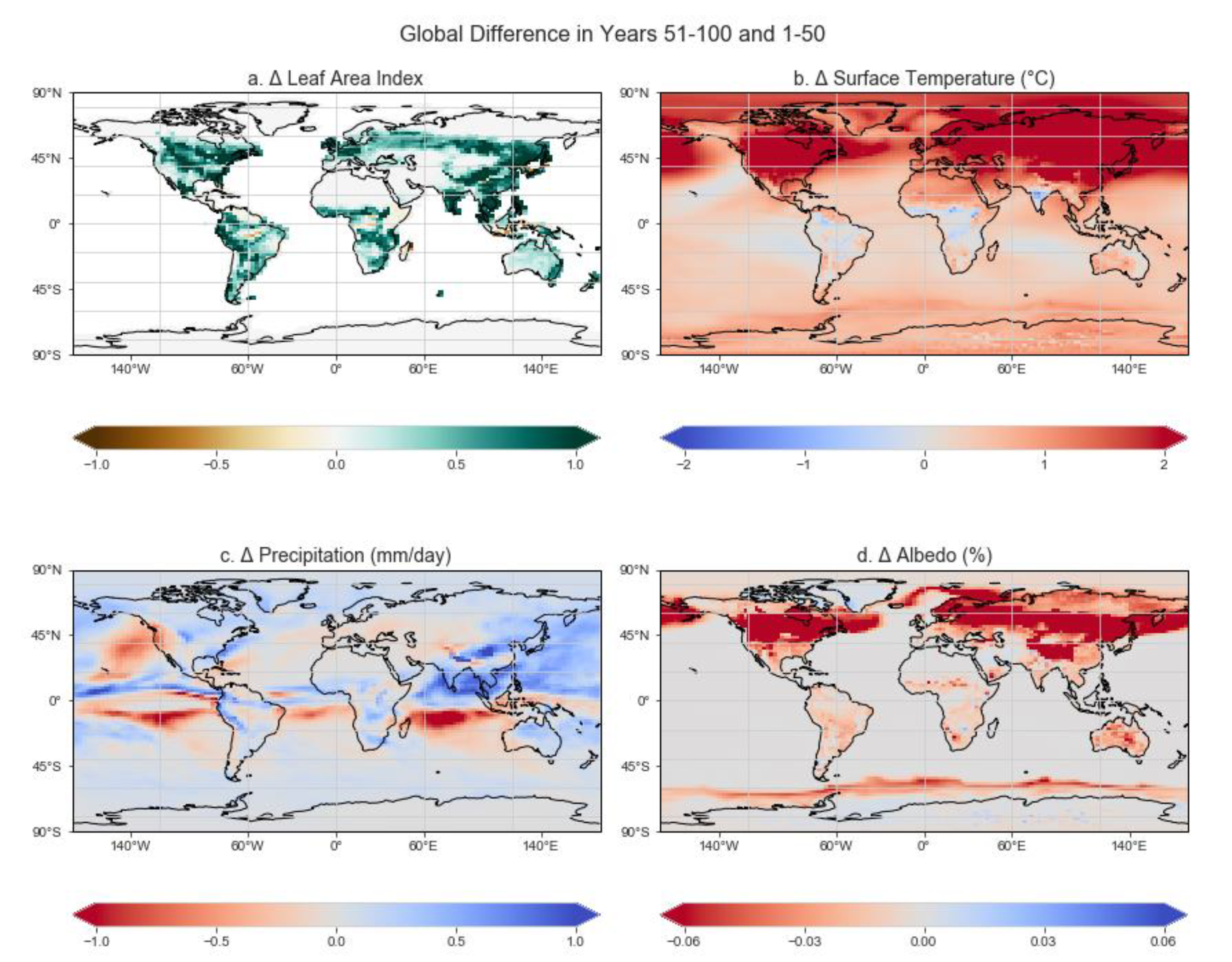
3.1. Global Biome Evolution
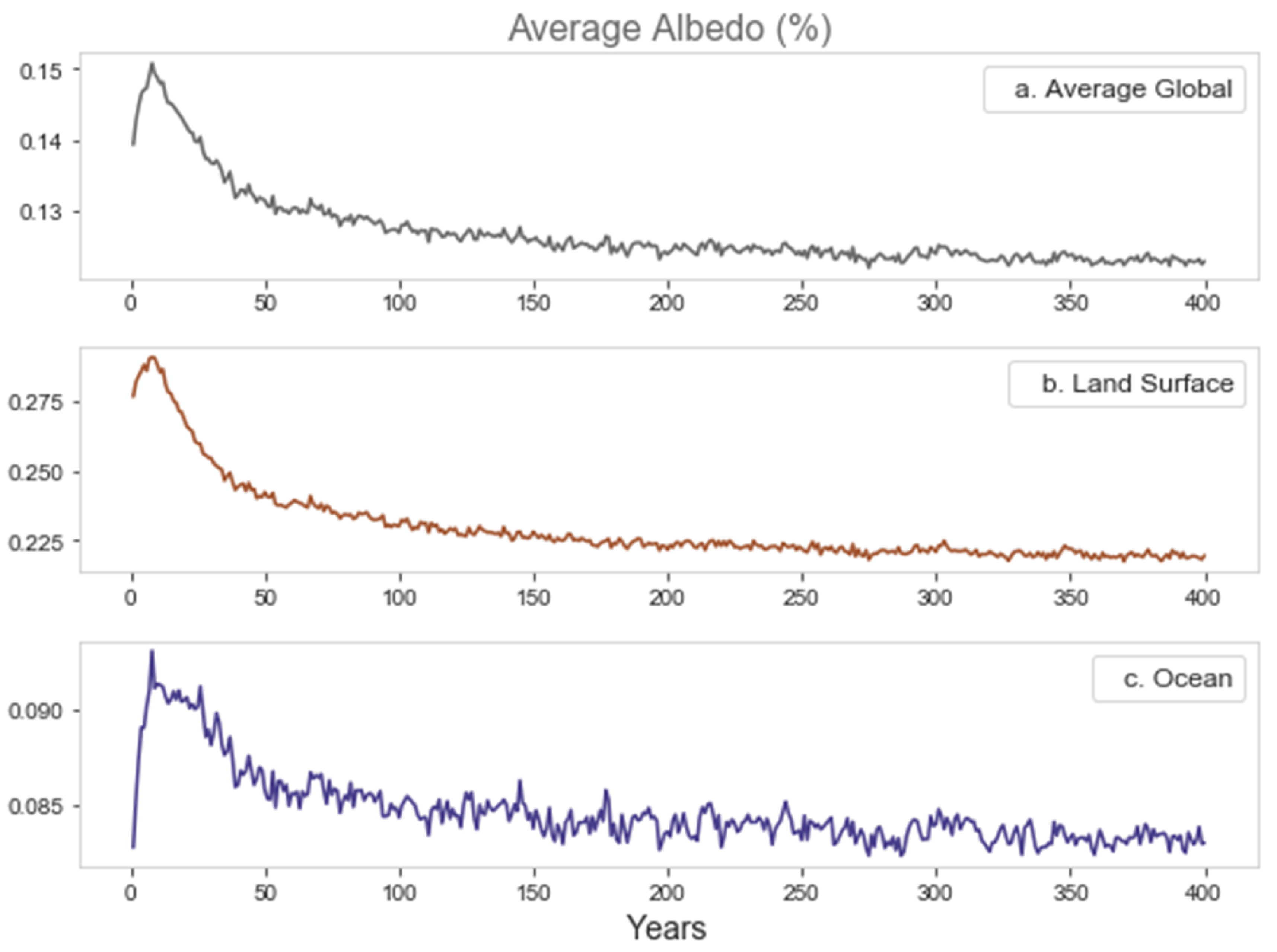
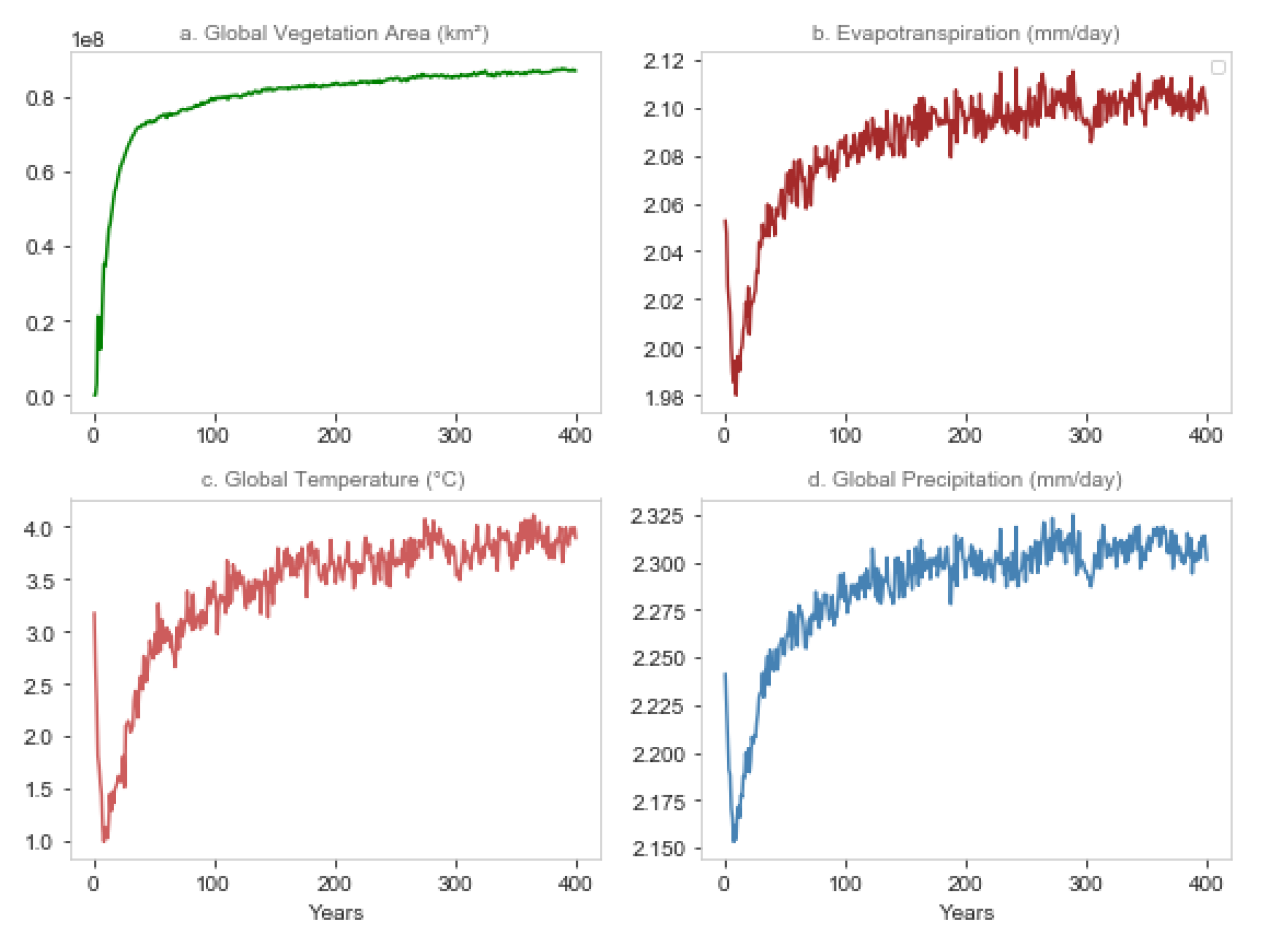
3.2. Global Climate Response
3.3. Latitudinal Responses
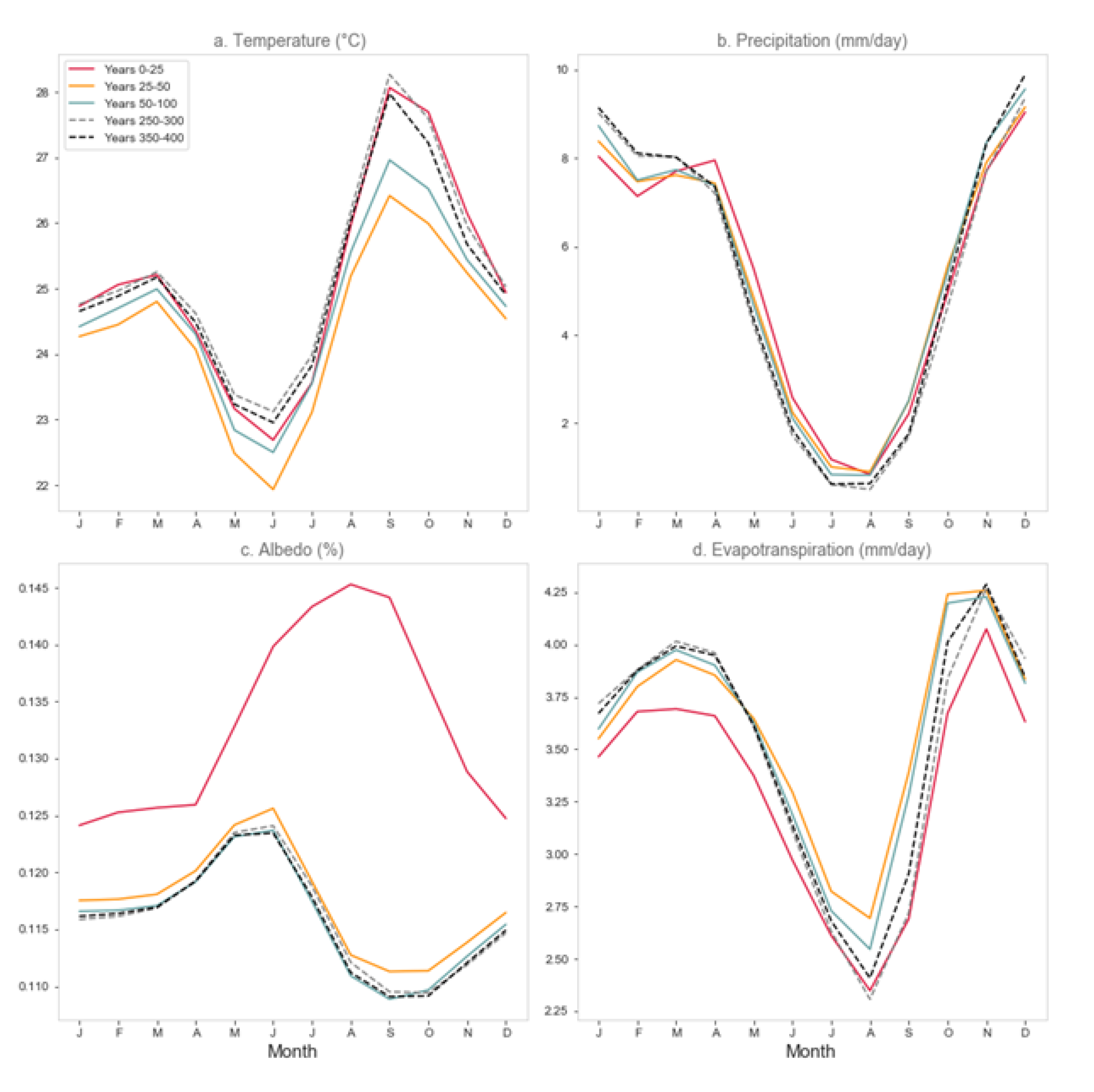
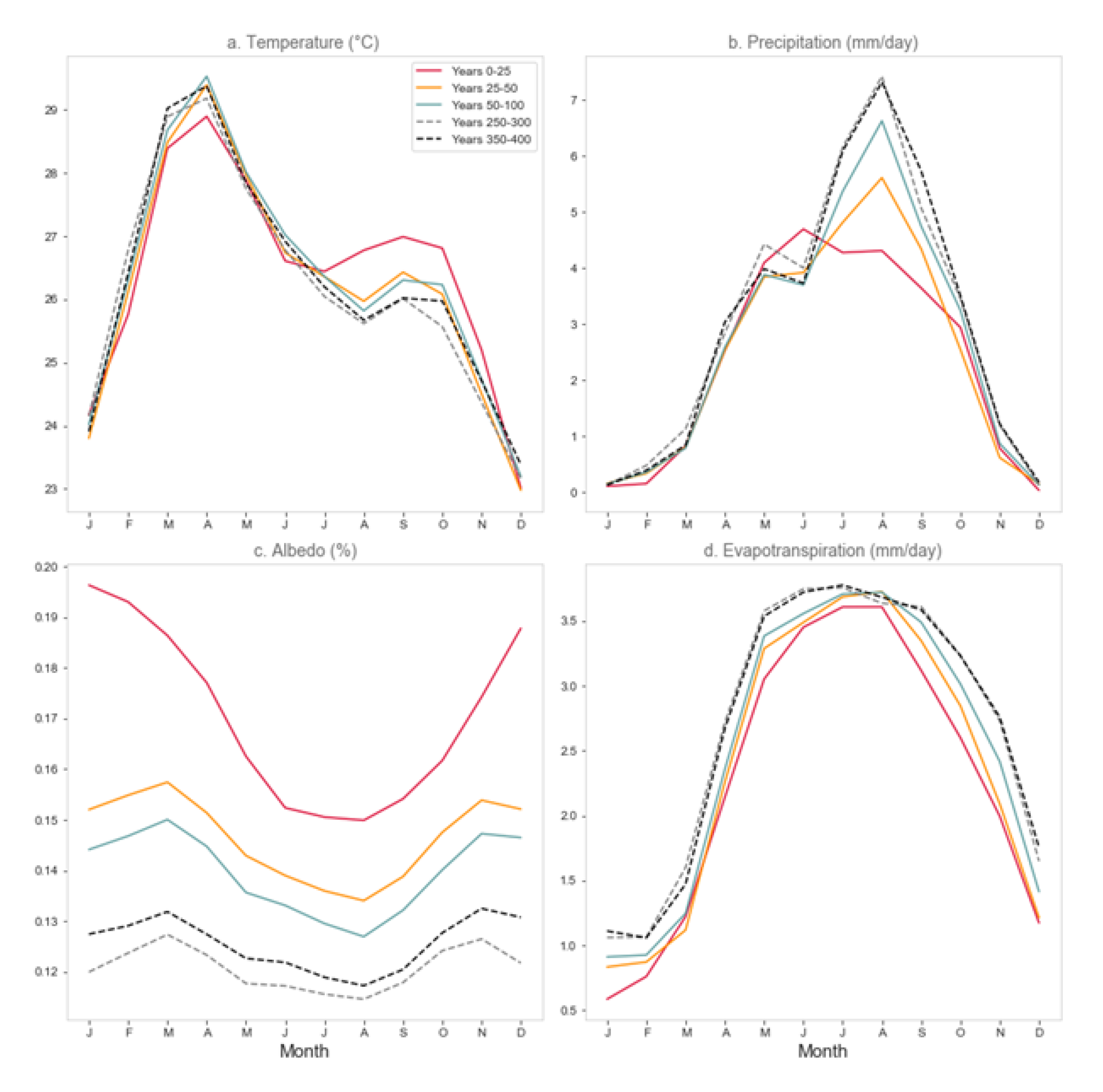
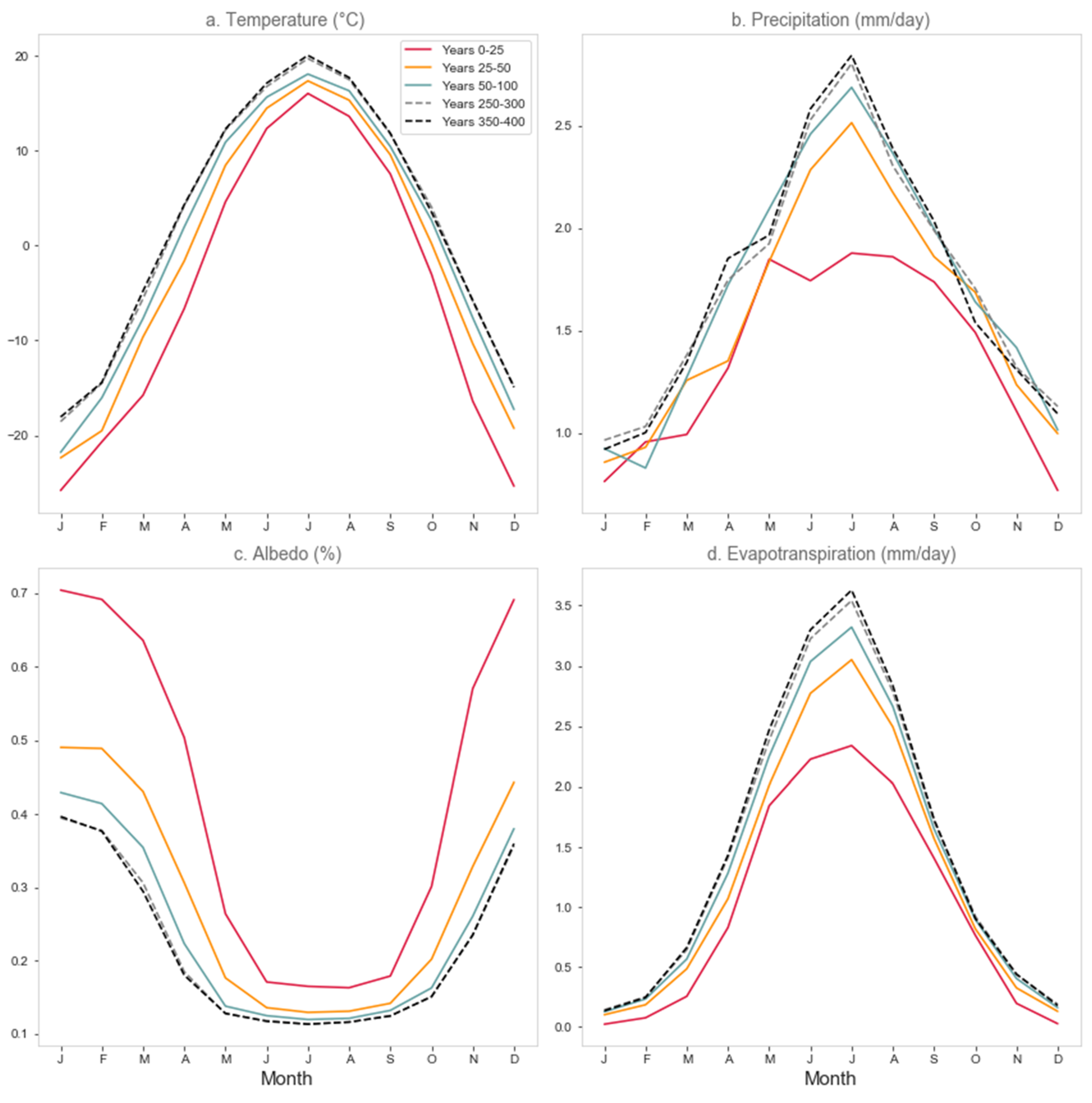
3.4. Regional Climate Response
3.4.1. Northeastern South America
3.4.2. West Africa
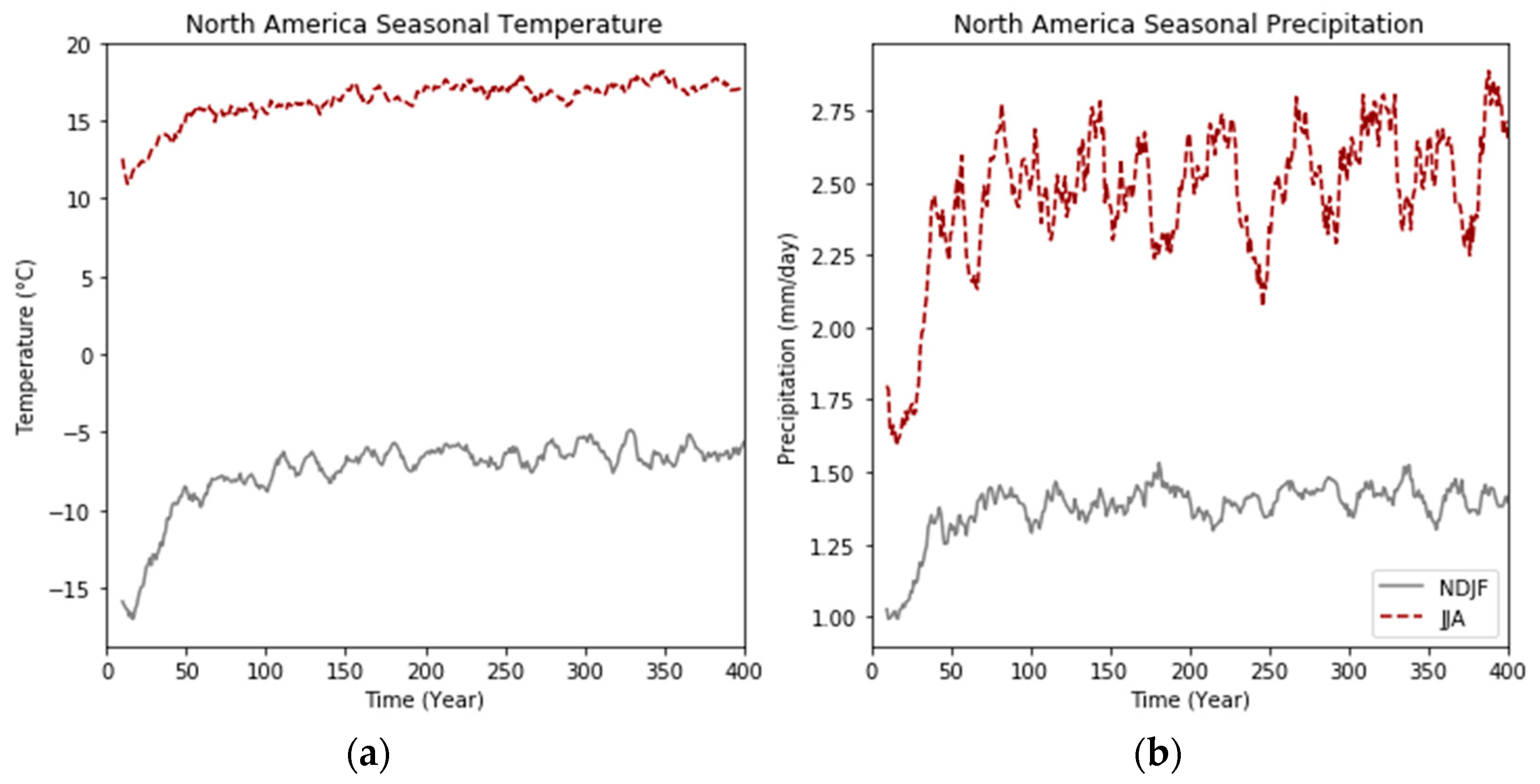
3.4.3. Midwestern North America
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boyce, C.K.; Lee, J.-E. Plant Evolution and Climate over Geological Timescales. Annu. Rev. Earth Planet. Sci. 2017, 45, 61–87. [Google Scholar] [CrossRef]
- Loranty, M.M.; Goetz, S.J.; Beck, P.S.A. Tundra vegetation effects on pan-arctic albedo. Environ. Res. Lett. 2011, 6, 024014. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swann, A.L.S.; Fung, I.Y.; Liu, Y.; Chiang, J.C.H. Remote Vegetation Feedbacks and the Mid-Holocene Green Sahara. J. Clim. 2014, 27, 4857–4870. [Google Scholar] [CrossRef]
- Bonan, G.B. Ecological Climatology: Concepts and Applications, 3rd ed.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Shukla, J.; Mintz, Y. Influence of Land-Surface Evapotranspiration on the Earth’s Climate. Science 1982, 215, 1498–1501. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Cox, P.M.; Betts, R.; Bopp, L.; Von Bloh, W.; Brovkin, V.; Cadule, P.; Doney, S.; Eby, M.; Fung, I.; et al. Climate–Carbon Cycle Feedback Analysis: Results from the C4MIP Model Intercomparison. J. Clim. 2006, 19, 3337–3353. [Google Scholar] [CrossRef]
- McGee, D.; Donohoe, A.; Marshall, J.; Ferreira, D. Changes in ITCZ location and cross-equatorial heat transport at the Last Glacial Maximum, Heinrich Stadial 1, and the mid-Holocene. Earth Planet. Sci. Lett. 2014, 390, 69–79. [Google Scholar] [CrossRef]
- Schneider, T.; Bischoff, T.; Haug, G.H. Migrations and dynamics of the intertropical convergence zone. Nature 2014, 513, 45–53. [Google Scholar] [CrossRef]
- Donohoe, A.; Marshall, J.; Ferreira, D.; Mcgee, D. The relationship between ITCZ location and cross-equatorial atmospheric heat transport: From the seasonal cycle to the Last Glacial Maximum. J. Clim. 2013, 26, 3597–3618. [Google Scholar] [CrossRef]
- Swann, A.L.S.; Fung, I.Y.; Chiang, J.C.H. Mid-latitude afforestation shifts general circulation and tropical precipitation. Proc. Natl. Acad. Sci. USA 2012, 109, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Boyce, C.K.; Brodribb, T.J.; Feild, T.S.; Zwieniecki, M.A. Angiosperm leaf vein evolution was physiologically and environmentally transformative. Proc. R. Soc. 2009, 276, 1771–1776. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B.; Pollard, D.; Thompson, S.L. Effects of boreal forest vegetation on global climate. Nature 1992, 359, 716–718. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Jackson, R.B. Vegetation–climate feedbacks in the conversion of tropical savanna to grassland. J. Clim. 2000, 13, 1593–1602. [Google Scholar] [CrossRef]
- Zhao, M.; Pitman, A.J.; Chase, T. The impact of land cover change on the atmospheric circulation. Clim. Dyn. 2001, 17, 467–477. [Google Scholar] [CrossRef]
- Eltahir, E.A.B. Role of vegetation in sustaining large-scale atmospheric circulations in the tropics. J. Geophys. Res. Space Phys. 1996, 101, 4255–4268. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formation from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- Schimel, D. Climate and Ecosystems. In Primers in Climate; Princeton University Press: Princeton, NJ, USA, in press.
- Oleson, K.W.; Dai, Y.; Bonan, G.; Dickinson, R.E.; Dirmeyer, P.A.; Hoffman, F.; Houser, P.; Levis, S.; Niu, G.-Y.; Thornton, P.; et al. Technical Description of the Community Land Model (CLM); Technical Report NCAR/TN461; National Center for Atmospheric Research: Boulder, CO, USA, 2004; p. 174. [Google Scholar]
- Levis, S.; Bonan, G.B.; Vertenstein, M.; Oleson, K.W. The Community Land Model’s Dynamic Global Vegetation Model (CLM-DGVM): Technical Description and User’s Guide; NCAR Technical Note NCAR/TN-459+IA; National Center for Atmospheric Research: Boulder, CO, USA, 2004; p. 50. [Google Scholar]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation mode. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Neale, R.B.; Richter, J.; Park, S.; Lauritzen, P.H.; Vavrus, S.J.; Rasch, P.J.; Zhang, M. The Mean Climate of the Community Atmosphere Model (CAM4) in Forced SST and Fully Coupled Experiments. J. Clim. 2013, 26, 5150–5168. [Google Scholar] [CrossRef] [Green Version]
- Randall, D. Atmosphere, Clouds, and Climate; Princeton University Press: Princeton, NJ, USA, 2012. [Google Scholar]
- Jeong, S.-J.; Ho, C.-H.; Kim, K.-Y.; Jeong, J.-H. Reduction of spring warming over East Asia associated with vegetation feedback. Geophys. Res. Lett. 2009, 36, L18705. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lintner, B.R.; Boyce, C.K.; Lawrence, P.J. Land use change exacerbates tropical South American drought by sea surface temperature variability. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Swann, A.L.S.; Fung, I.Y.; Levis, S.; Bonan, G.B.; Doney, S.C. Changes in Arctic vegetation amplify high-latitude warming through the greenhouse effect. Proc. Natl. Acad. Sci. USA 2010, 107, 1295–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerten, D.; Schaphoff, S.; Haberlandt, U.; Lucht, W.; Sitch, S. Terrestrial vegetation and water balance—Hydrological evaluation of a dynamic global vegetation model. J. Hydrol. 2004, 286, 249–270. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Midwestern North America | Northeastern South America | West Africa | |
|---|---|---|---|
| Latitude Range | −100° to −85° | 0° to −15° | 7° to 13° |
| Longitude Range | 45° to 55° | −60° to −50° | 2° to −12° |
| Vegetation | Temperate forest | Tropical Rainforest | Dry forest |
| Grasses | Trees | Shrubs |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levey, J.; Lee, J.-E. Global and Regional Implications of Biome Evolution on the Hydrologic Cycle and Climate in the NCAR Dynamic Vegetation Model. Land 2020, 9, 342. https://doi.org/10.3390/land9100342
Levey J, Lee J-E. Global and Regional Implications of Biome Evolution on the Hydrologic Cycle and Climate in the NCAR Dynamic Vegetation Model. Land. 2020; 9(10):342. https://doi.org/10.3390/land9100342
Chicago/Turabian StyleLevey, Jessica, and Jung-Eun Lee. 2020. "Global and Regional Implications of Biome Evolution on the Hydrologic Cycle and Climate in the NCAR Dynamic Vegetation Model" Land 9, no. 10: 342. https://doi.org/10.3390/land9100342
APA StyleLevey, J., & Lee, J. -E. (2020). Global and Regional Implications of Biome Evolution on the Hydrologic Cycle and Climate in the NCAR Dynamic Vegetation Model. Land, 9(10), 342. https://doi.org/10.3390/land9100342