Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
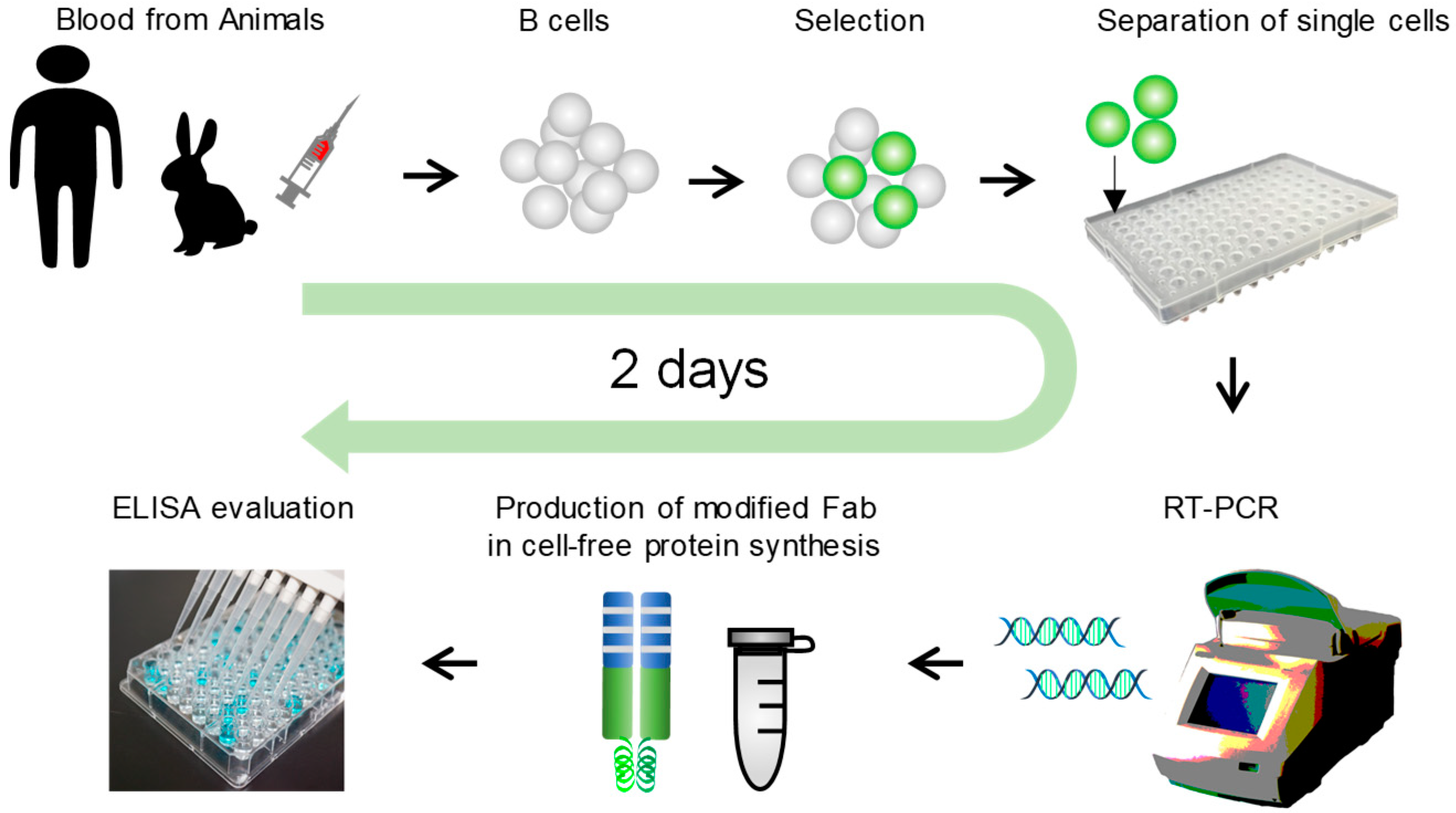
2.1. Overview of Ecobody Technology
2.2. Preparation of Antigens and Immunization of Rabbits
2.3. Preparation of EBV Infected Human Cells
2.4. Selection of B Cells
2.4.1. Selection of Rabbit B Cells Binding to Bacterial Cells
2.4.2. Selection of Rabbit B Cells Producing mAbs to Nontoxic VT2
2.4.3. Selection of Influenza Vaccine Reactive EBV Infected Human Cells
2.5. RT-PCR
2.6. Cell-Free Protein Synthesis (CFPS)
2.7. ELISA
2.8. mAb Production in E. coli
3. Results
3.1. Rabbit mAbs Screening
3.1.1. V. parahaemolyticus
3.1.2. E. coli O26
3.1.3. Nontoxic VT2
3.2. Expression in E. coli
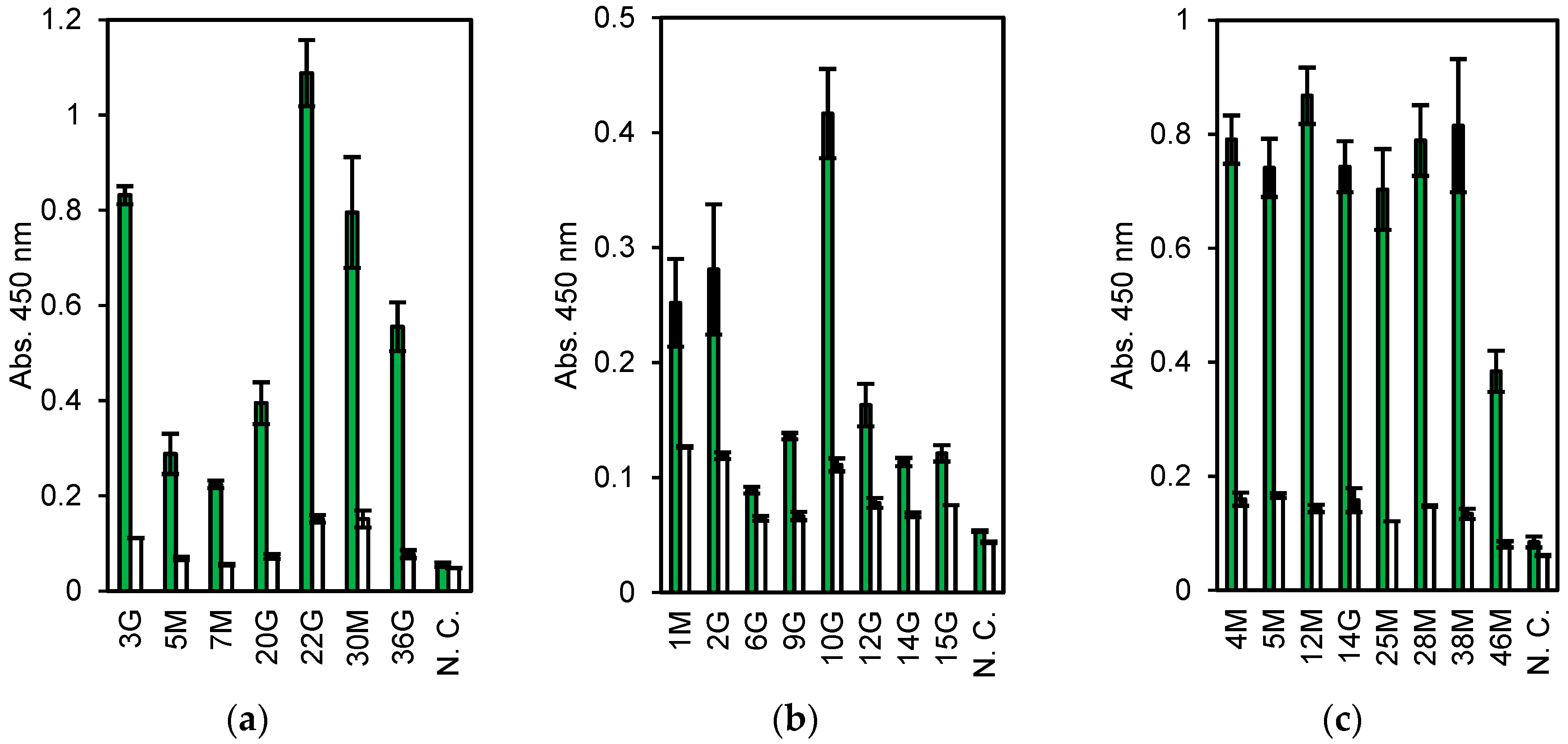
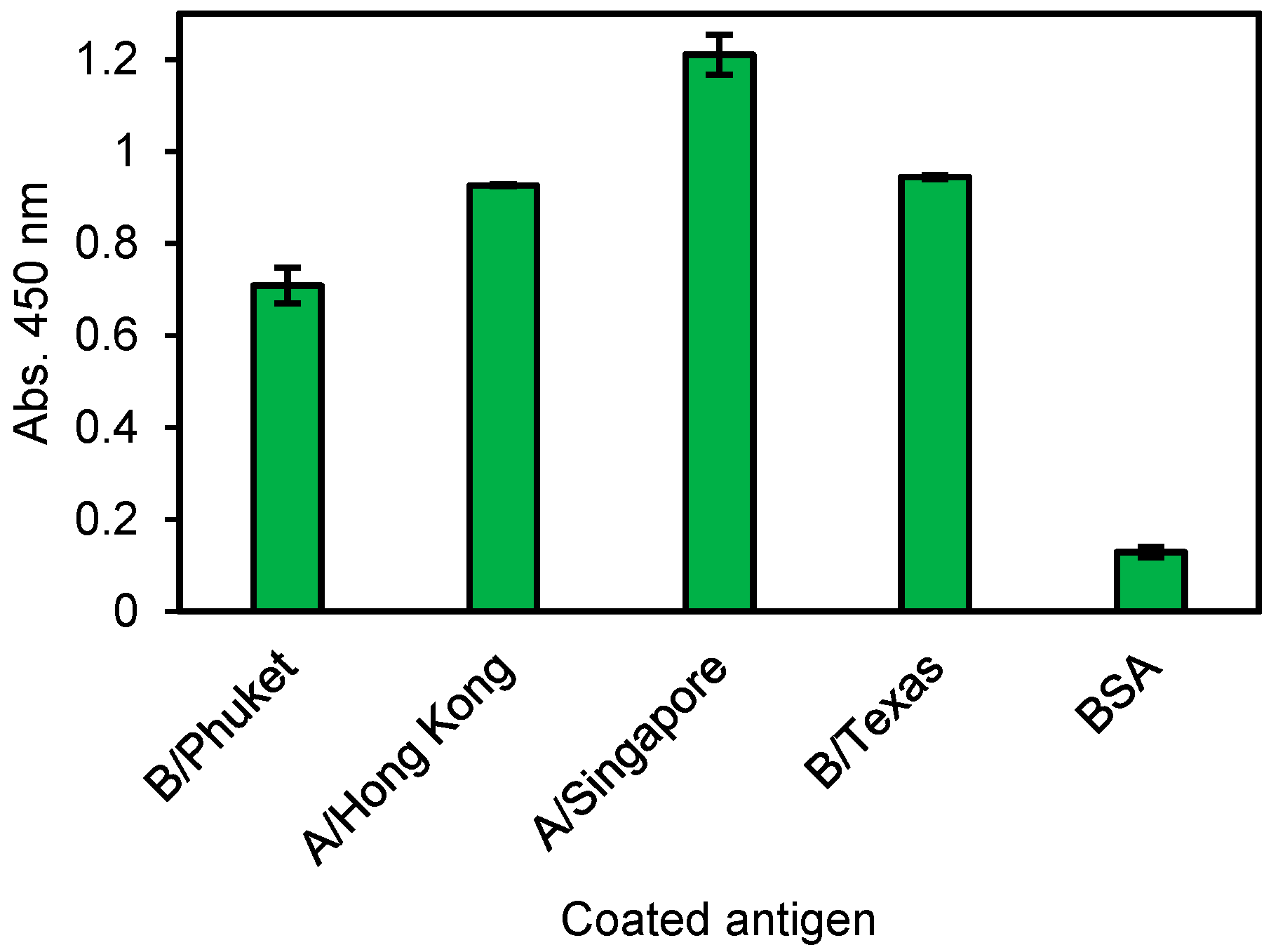
3.3. Human mAbs Screening
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tiller, T. Single B cell antibody technologies. New Biotechnol. 2011, 28, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, N.; Yoshioka, M.; Fujimoto, R.; Yamagishi, F.; Isobe, M. Rapid production of antigen-specific monoclonal antibodies from a variety of animals. BMC Biol. 2012, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Seeber, S.; Ros, F.; Thorey, I.; Tiefenthaler, G.; Kaluza, K.; Lifke, V.; Fischer, J.A.; Klostermann, S.; Endl, J.; Kopetzki, E.; et al. A robust high throughput platform to generate functional recombinant monoclonal antibodies using rabbit B cells from peripheral blood. PLoS ONE 2014, 9, e86184. [Google Scholar] [CrossRef] [PubMed]
- Starkie, D.O.; Compson, J.E.; Rapecki, S.; Lightwood, D.J. Generation of Recombinant Monoclonal Antibodies from Immunised Mice and Rabbits via Flow Cytometry and Sorting of Antigen-Specific IgG+ Memory B Cells. PLoS ONE 2016, 11, e0152282. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.S.; Tang, A.; Chen, Z.; Horton, M.S.; Yan, H.; Wang, X.M.; Dubey, S.A.; DiStefano, D.J.; Ettenger, A.; Fong, R.H.; et al. Rapid isolation of dengue-neutralizing antibodies from single cell-sorted human antigen-specific memory B-cell cultures. MAbs 2016, 8, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, H.; Kanamori, T.; Shimizu, Y.; Ueda, T. A highly controllable reconstituted cell-free system—A breakthrough in protein synthesis research. Curr. Pharm. Biotechnol. 2010, 11, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.D.; Gan, R.; Hodgman, C.E.; Jewett, M.C. Cell-free protein synthesis: Applications come of age. Biotechnol. Adv. 2012, 30, 1185–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemella, A.; Thoring, L.; Hoffmeister, C.; Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. Chembiochem. 2015, 16, 2420–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Suzuki, H.; Hanai, Y.; Wada, F.; Hitomi, K.; Yamane, T.; Nakano, H. A novel strategy for generation of monoclonal antibodies from single B cells using rt-PCR technique and in vitro expression. Biotechnol. Prog. 2006, 22, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Sabrina, Y.; Ali, M.; Nakano, H. In vitro generation of anti-hepatitis B monoclonal antibodies from a single plasma cell using single-cell RT-PCR and cell-free protein synthesis. J. Biosci. Bioeng. 2010, 109, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Hashimura, D.; Kojima, T.; Minabe, S.; Nakano, H. In vitro generation of rabbit anti-Listeria monocytogenes monoclonal antibody using single cell based RT-PCR linked cell-free expression systems. J. Immunol. Methods 2015, 427, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Glabe, C.; Luecke, H. Crystal structure of a conformation-dependent rabbit IgG Fab specific for amyloid prefibrillar oligomers. Biochim. Biophys. Acta 2012, 1820, 1908–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groff, D.; Armstrong, S.; Rivers, P.J.; Zhang, J.; Yang, J.; Green, E.; Rozzelle, J.; Liang, S.; Kittle, J.D.; Steiner, A.R.; et al. Engineering toward a bacterial ‘endoplasmic reticulum’ for the rapid expression of immunoglobulin proteins. Mabs 2014, 6, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Hanson, J.A.; Steiner, A.R.; Tran, C.; Masikat, M.R.; Chen, R.; Zawada, J.F.; Sato, A.K.; Hallam, T.J.; Yin, G. A simplified and robust protocol for immunoglobulin expression in Escherichia coli cell-free protein synthesis systems. Biotechnol. Prog. 2015, 31, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Fukui, K.; Yamamoto, H.; Hashimura, D.; Miyake, S.; Hirakawa, Y.; Yamasaki, T.; Kojima, T.; Nakano, H. ‘Zipbody’ leucine zipper-fused Fab in E. coli in vitro and in vivo expression systems. Protein Eng. Des. Sel. 2016, 29, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Nagai, S.; Nakano, H. N-terminal SKIK peptide tag markedly improves expression of difficult-to-express proteins in Escherichia coli and Saccharomyces cerevisiae. J. Biosci. Bioeng. 2017, 123, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Ojima-Kato, T.; Nagai, S.; Nakano, H. Ecobody technology: Rapid monoclonal antibody screening method from single B cells using cell-free protein synthesis for antigen-binding fragment formation. Sci. Rep. 2017, 7, 13979. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McMahon, S.; Skinner, C.; Merrill, P.; Scotcher, M.C.; Stanker, L.H. Development and characterization of monoclonal antibodies against Shiga toxin 2 and their application for toxin detection in milk. J. Immunol. Methods 2013, 389, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, M.; Goto, M.; Noda, S.; Sagara, M.; Yamada, S.; Yamamoto, N.; Kawakami, Y.; Matsui, Y.; Miyazawa, Y.; Yamazaki, H.; et al. DNA repair capacity measured by high throughput alkaline comet assays in EBV-transformed cell lines and peripheral blood cells from cancer patients and healthy volunteers. Mutat. Res. 2005, 588, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tsumoto, K.; Shinoki, K.; Kondo, H.; Uchikawa, M.; Juji, T.; Kumagai, I. Highly efficient recovery of functional single-chain Fv fragments from inclusion bodies overexpressed in Escherichia coli by controlled introduction of oxidizing reagent—Application to a human single-chain Fv fragment. J. Immunol. Methods 1998, 219, 119–129. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Concentration (mg/mL) | Total Volume (mL) | Amount of Protein (mg) | Recovery Ratio (%) | |
|---|---|---|---|---|
| Solubilization by 6 M GuHCl | 1.62 | 0.95 | 1.54 | |
| Refolding | 0.98 | 1.3 | 1.28 | 82.8 |
| His tag purification | 0.094 | 4.5 | 0.423 | 27.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojima-Kato, T.; Morishita, S.; Uchida, Y.; Nagai, S.; Kojima, T.; Nakano, H. Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology. Antibodies 2018, 7, 38. https://doi.org/10.3390/antib7040038
Ojima-Kato T, Morishita S, Uchida Y, Nagai S, Kojima T, Nakano H. Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology. Antibodies. 2018; 7(4):38. https://doi.org/10.3390/antib7040038
Chicago/Turabian StyleOjima-Kato, Teruyo, Shiomi Morishita, Yoshino Uchida, Satomi Nagai, Takaaki Kojima, and Hideo Nakano. 2018. "Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology" Antibodies 7, no. 4: 38. https://doi.org/10.3390/antib7040038
APA StyleOjima-Kato, T., Morishita, S., Uchida, Y., Nagai, S., Kojima, T., & Nakano, H. (2018). Rapid Generation of Monoclonal Antibodies from Single B Cells by Ecobody Technology. Antibodies, 7(4), 38. https://doi.org/10.3390/antib7040038