Secretory Structures of Pogostemon auricularius: Morphology, Development, and Histochemistry


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM)
2.3. Light Microscopy
3. Results
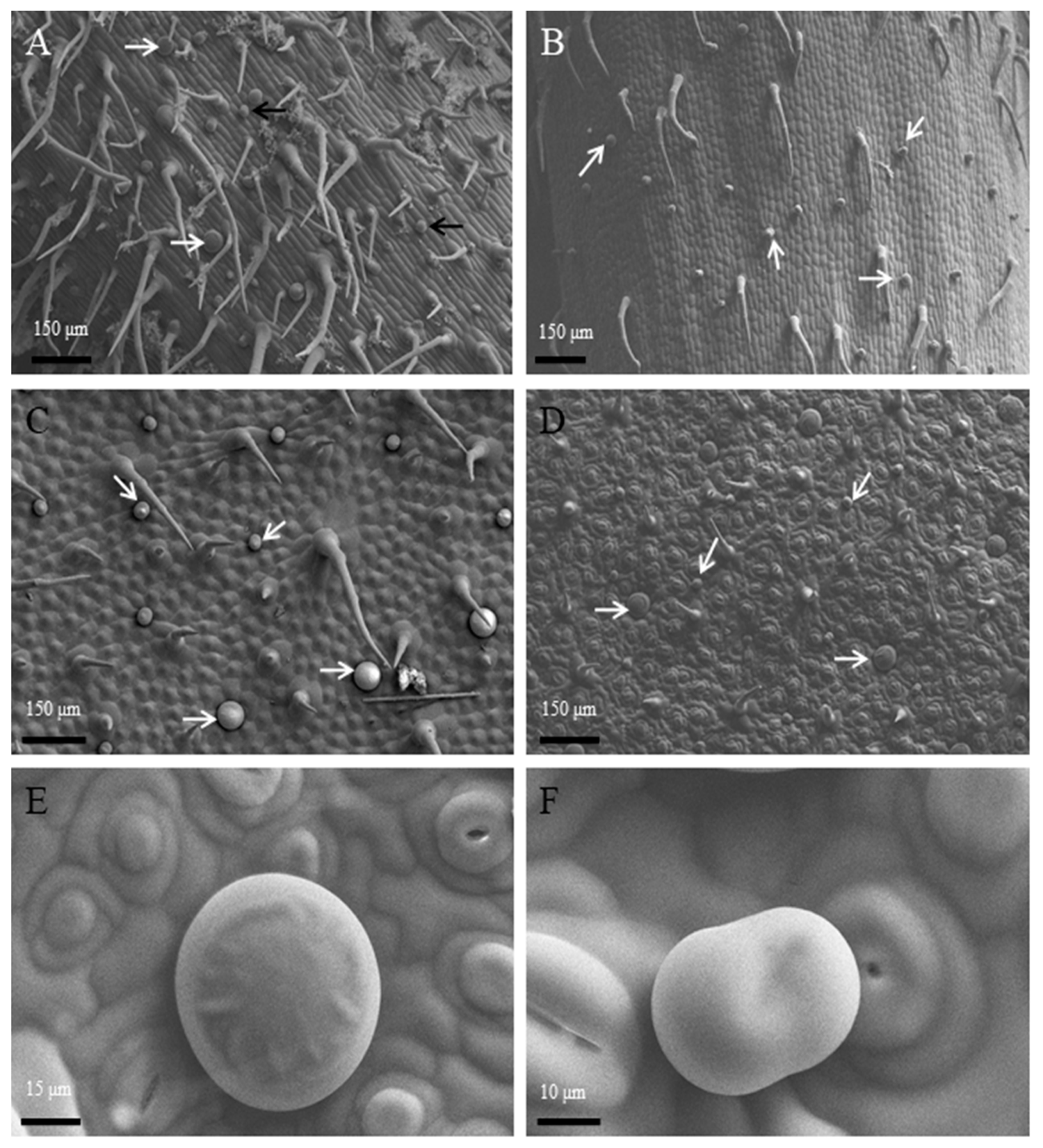
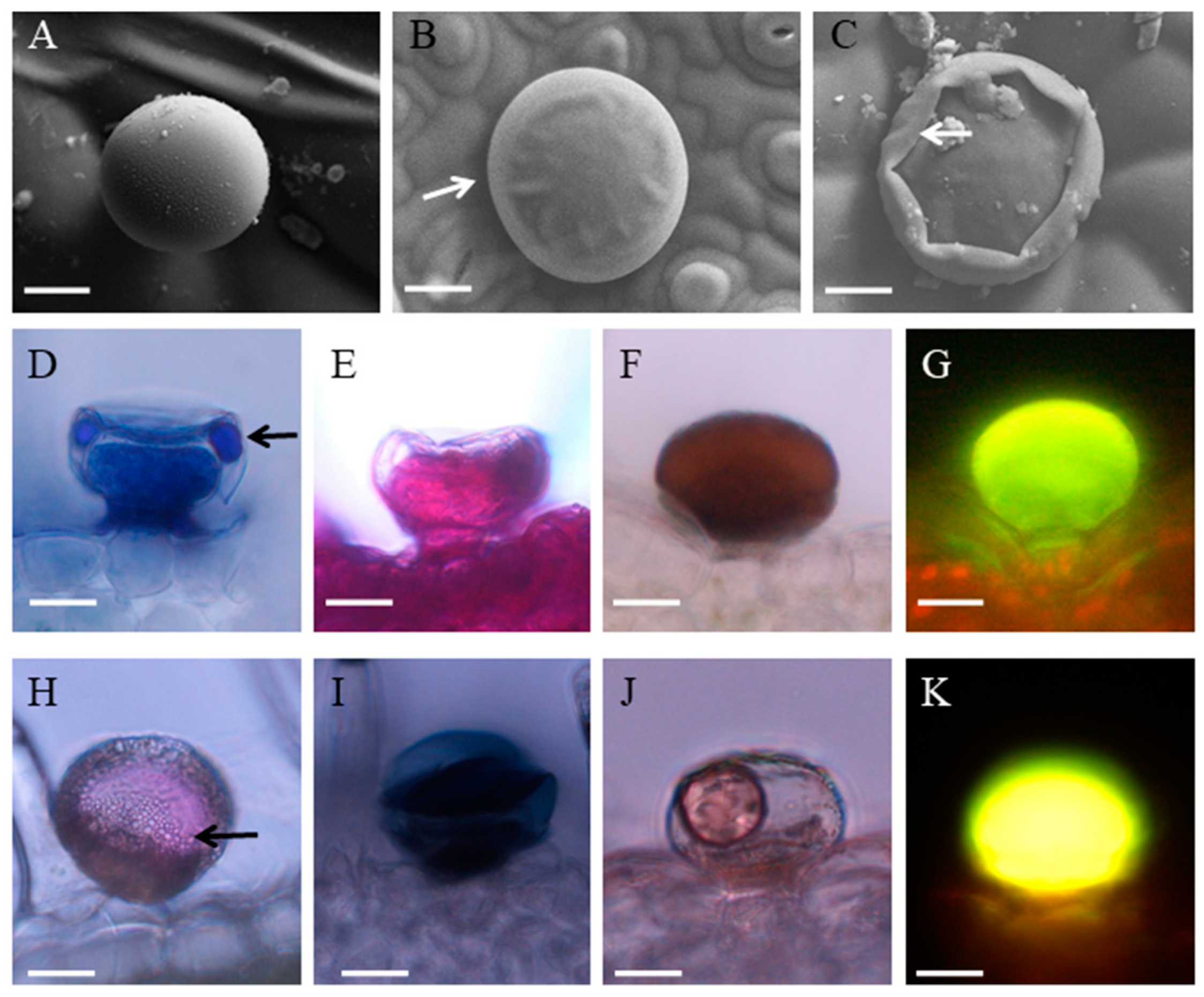
3.1. Morphology, Distribution and Histochemistry of Secretory Structures
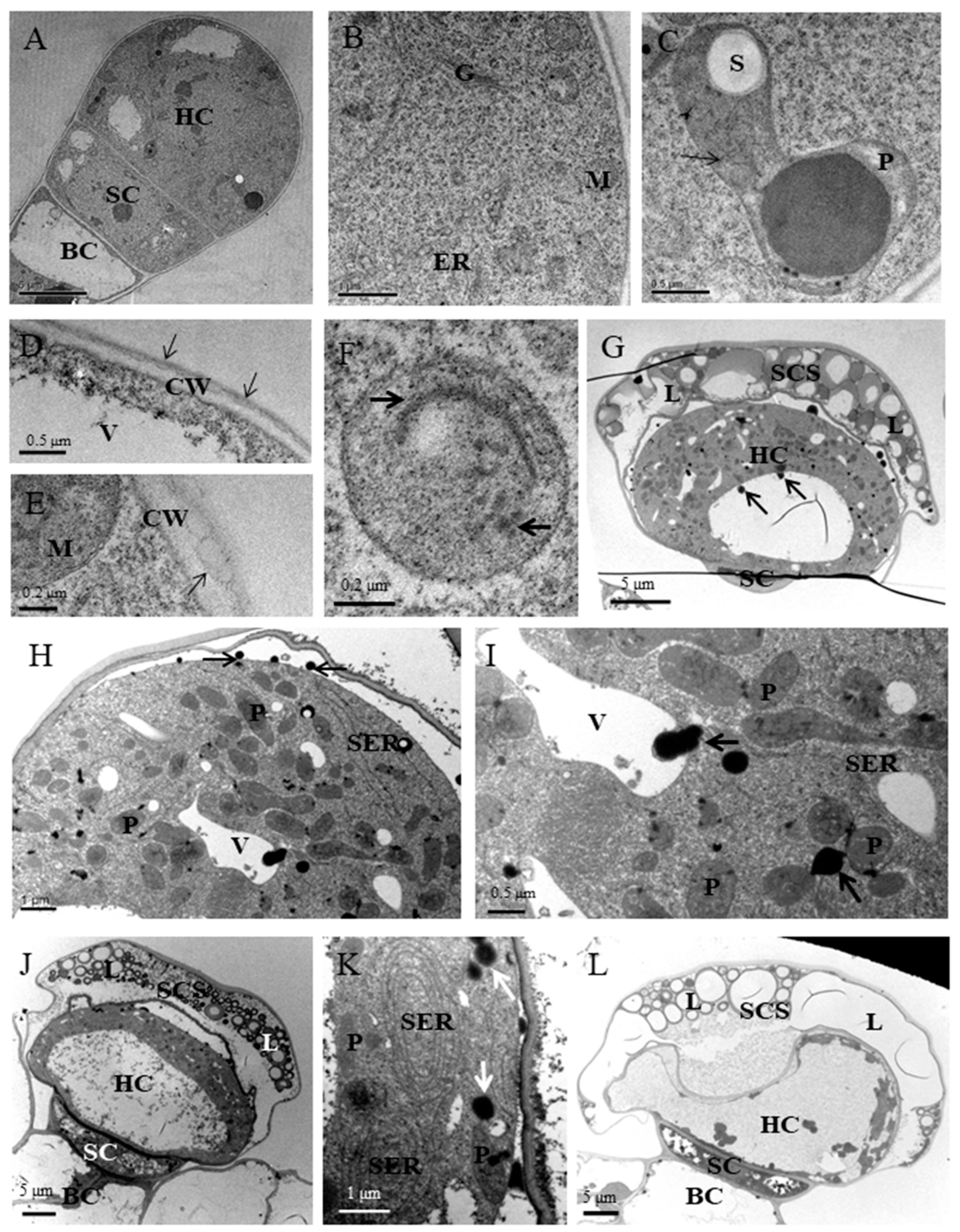
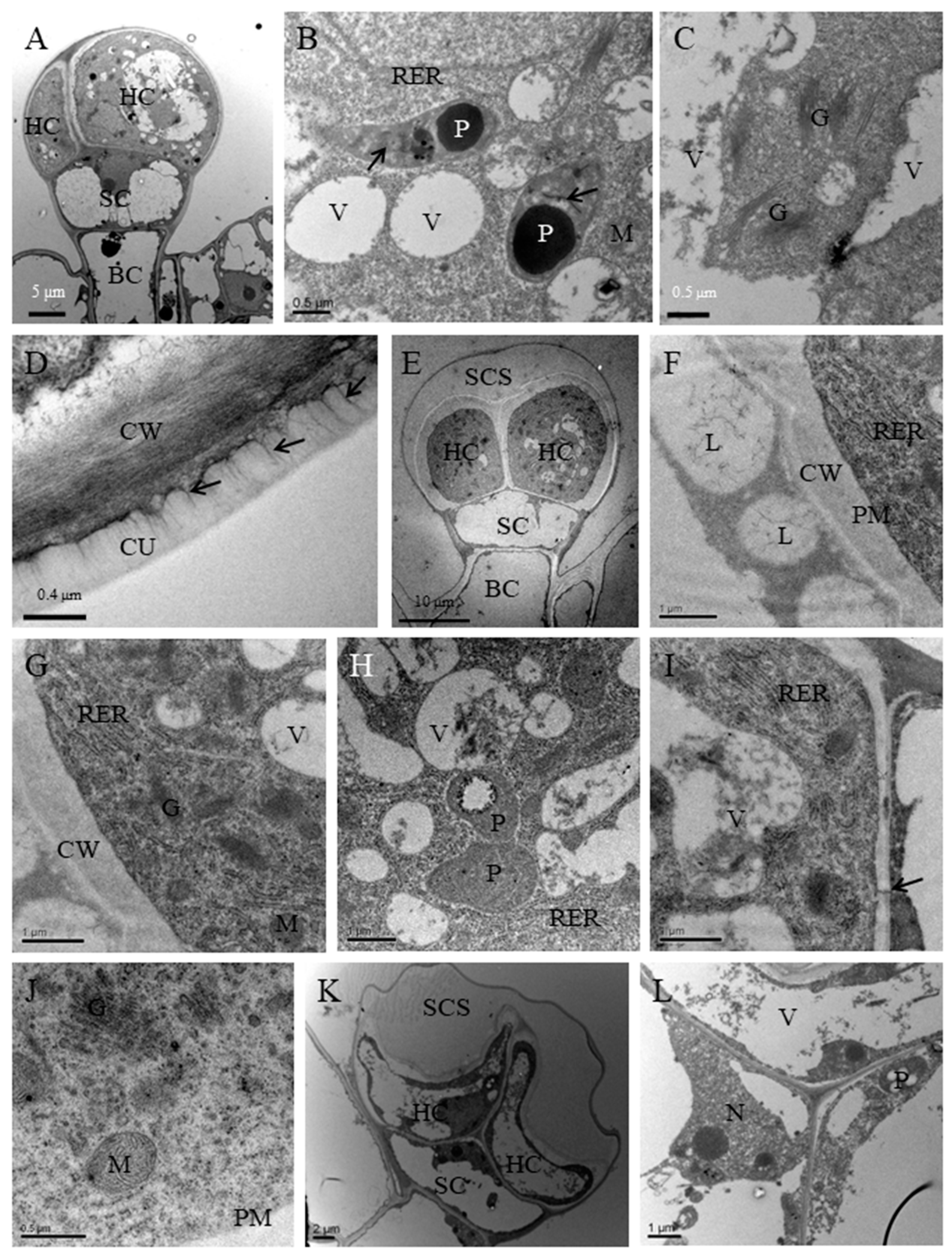
3.2. Ultrastructural Aspects of Secretory Structures
4. Discussion
4.1. Analysis in Morphology, Development and Histochemistry
4.2. Site of Secondary Metabolism Biosynthesis
4.3. Possible Secretory Mechanisms
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Babak, B.A.; Mahmoud, B.; Pegah, T.; Nasrollah, N.; Mahmoud, R.K. An ethno-medicinal study of medicinal plants used for the treatment of diabetes. J. Nephropathol. 2016, 5, 44–50. [Google Scholar]
- Heinrich, G.; Schultze, W. Composition and site of biosynthesis of the essential oil in fruits of Phellodendron amurense Rupr. (Rutaceae). Isr. J. Bot. 1985, 34, 205–217. [Google Scholar]
- Machado, S.R.; Gregorio, E.A.; Guimaraes, E. Ovary peltate trichomes of Zeyheria montana (Bignoniaceae): Developmental ultrastructure and secretion in relation to function. Ann. Bot. 2006, 97, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Thai, T.H.; Hung, N.Q.; Minh, D.T. Chemical composition of the essential oil of Pogostemon auricularius (L.) Hassk. from Vietnam. Tap Chi Sinh Hoc 2014, 28. [Google Scholar] [CrossRef]
- Falak, A.H.; Sunita, A.; Raja, R. Novel cleistanthane diterpenoids from Pogostemon auricularis. J. Nat. Prod. 1988, 51, 212–216. [Google Scholar]
- Falak, A.H.; Raja, R.; Aboo, S. Structure and configuration of a new cleistanthane diterpenoid from Pogostemon auricularis. Nat. Prod. Lett. 1993, 3, 71–77. [Google Scholar]
- Jacek, J.; Agata, K.; Bożena, D. Micromorphological and histochemical attributes of flowers and floral reward in Linaria vulgaris (Plantaginaceae). Protoplasma 2018, 255, 1763–1776. [Google Scholar]
- Bosabalidis, A.M. Structural features of Origanum species. In Oregano: The Genera Origanum and Lippia, 1st ed.; Kintzios, S.E., Ed.; Taylor and Francis: London, UK, 2002; pp. 11–64. [Google Scholar]
- Combrinck, S.; Duplooy, G.W.; Mccrindle, R.I. Morphology and histochemistry of the glandular trichomes of Lippia scaberrima (Verbenaceae). Ann. Bot. 2007, 99, 1111–1119. [Google Scholar] [CrossRef]
- Agata, K. Comparative micromorphology and anatomy of flowers and floral secretory structures in two Viburnum species. Protoplasma 2017, 254, 523–537. [Google Scholar]
- Ascensăo, L.; Marques, N.; Pais, M.S. Peltate glandular trichomes of Leonotis leonurus leaves: Ultrastructure and histochemical characterization of secretions. Int. J. Plant Sci. 1997, 158, 249–258. [Google Scholar] [CrossRef]
- Nick, B.; Stefan, B.; Frank, S.; Gerd, H.; Alain, T. The development of type VI glandular trichomes in the cultivated tomato Solanum lycopersicum and a related wild species S. habrochaites. BMC Plant Biol. 2015, 15, 289. [Google Scholar]
- Hanlidou, E.; Kokkini, S.; Bosabalidis, A.M. Glandular trichomes and essential oil constituents of Calamintha menthifolia (Lamiaceae). Plant Syst. Evol. 1991, 177, 17–26. [Google Scholar] [CrossRef]
- Ascensăo, L.; Mota, L.; Castro, M.M. Glandular Trichomes on the Leaves and Flowers of Plectranthus ornatus: Morphology, distribution and histochemistry. Ann. Bot. 1999, 84, 437–447. [Google Scholar] [CrossRef]
- Karousou, R.; Bosabalidis, A.M.; Kokkini, S. Sideritis syriaca ssp. syriaca: Glandular trichome structure and development in relation to systematics. Nord. J. Bot. 1992, 12, 31–37. [Google Scholar] [CrossRef]
- Dagmar, K.; Maria, M. Light, conventional and environmental scanning electron microscopy of the trichomes of Cucurbita pepo subsp. pepo var. styriaca and histochemistry of glandular secretory products. Ann. Bot. 2004, 94, 515–526. [Google Scholar]
- Lawrence, B.M. Chemical components of labiate oils and their exploitation. In Advances of Labiatae Science; Harlrey, R.M., Reynolds, T., Eds.; The Royal Botanic Gardens Kew: Richmond, BC, Canada, 1992; pp. 399–436. [Google Scholar]
- Spring, O. Chemotaxonomy based on metabolites from glandular trichomes. Adv. Bot. Res. 2000, 31, 153–174. [Google Scholar]
- Kelsey, R.G.; Reynolds, G.W.; Rodriguez, E. The chemistry of biologically active constituents secreted and stored in plant glandular trichomes. In Biology and Chemistry of Plant Trichomes; Rodriguez, E., Healey, P.L., Mehta, I., Eds.; Plenum Press: New York, NY, USA, 1984; pp. 187–244. [Google Scholar]
- Richardson, P.M. The chemistry of the Labiatae: An introduction and overview. In Advances in Labiate Science; Harley, R.M., Reynolds, T., Eds.; Royal Botanic Gardens: Kew, UK, 1992; pp. 291–297. [Google Scholar]
- Kokkini, S.; Karousou, R.; Vokou, D. Pattern of geographic variation of Origanum ulgare trichomes and essential oil content in Greece. Biochem. Syst. Ecol. 1994, 22, 517–528. [Google Scholar] [CrossRef]
- Clark, L.J.; Hamilton, J.G.G.; Chapman, J.V.; Rhodes, M.J.C.; Hallanhan, D.L. Analysis of monoterpenoids in glandular trichomes of catmint Nepeta racemosa. Plant J. 1997, 11, 1387–1393. [Google Scholar] [CrossRef]
- Wood, B.W. Flavonoids, alkali earth, and rare earth elements affect pecan pollen germination. HortScience 2017, 52, 85–88. [Google Scholar] [CrossRef]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Dell, B.; McComb, A.J. Plant resins: Their formation, secretion, and possible functions. Adv. Bot. Res. 1978, 6, 277–316. [Google Scholar]
- Schnepf, E. Tubuläres endoplasmatisches Reticulum in Drüsen mit lipophilen Ausscheidungen von Ficus, Ledum, und Salvia. Biochem. Physiol. Pflanz. 1972, 163, 113–125. [Google Scholar] [CrossRef]
- Amelunxen, F. Elektronenmikroskopische Untersuchungen an den Drüsenschuppen von Mentha piperita L. Planta Medial 1965, 13, 457–473. [Google Scholar] [CrossRef]
- Bosabalidis, A.; Tsekos, I. Glandular scale development and essential oil secretion in dictamnus L. Planta 1982, 156, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Bourett, T.M.; Howard, R.J.; O’Keefe, D.P.; Hallahan, D.L. Gland development on leaf surfaces of Nepeta racemosa. Int. J. Plant Sci. 1994, 155, 623–632. [Google Scholar] [CrossRef]
- Turner, G.W.; Gershenzon, J.; Nielson, E.E.; Froehlich, J.E.; Croteau, R.B. Limonene synthase, the enzyme responsible for monoterpene biosynthesis in peppermint, is localized to leucoplasts of oil gland secretory cells. Plant Physiol. 1999, 120, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, W.; Schwarz, M.; Cartayrade, A.; Arigoni, D.; Zenk, M.H.; Bacher, A. The deoxyxylulose phosphate pathway of terpenoid biosynthesis in plants and microorganisms. Chem. Biol. 1998, 5, R221–R223. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. The 1-deoxy-d-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Phys. 1999, 50, 47–66. [Google Scholar] [CrossRef]
- Skubatz, H.; Kunkell, D.D.; Patt, J.M.; Howald, W.N.; Hartman, T.G.; Meeuse, B.J.D. Pathway of terpene excretion by appendix of Sauromatum guttatum. Proc. Natl. Acad. Sci. USA 1995, 92, 10084–10088. [Google Scholar] [CrossRef] [PubMed]
- Valkama, E.; Salminen, J.P.; Koricheva, J.; Pihlaja, K. Comparative analysis of leaf trichome structure and composition of epicuticular flavonoids in Finnis Birch species. Ann. Bot. 2003, 91, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Dhugga, K.S. Plant Golgi cell wall synthesis: From genes to enzyme activities. Proc. Natl. Acad. Sci. USA 2005, 102, 1815–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, J.M.; Nepi, M.; Galetto, L.; Guimarães, E.; Machado, S.R. Functional aspects of floral nectar secretion of Ananas ananassoides, an ornithophilous bromeliad from the Brazilian savanna. Ann Bot. 2012, 109, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Kirchoff, B.K.; Liao, J.P. The capitate and peltate glandular trichomes of Lavandula pinnata L. (Lamiaceae): Histochemistry, ultrastructure, and secretion. J. Torrey Bot. Soc. 2008, 135, 155–167. [Google Scholar] [CrossRef]
- Robins, R.J.; Juniper, B.E. The secretory cycle of Dionea muscipula Ellis. I. Fine structure and the effect of stimulation on the fine structure of the glands. New Phytol. 1980, 86, 279–296. [Google Scholar] [CrossRef]
- Tse, Y.C.; Mo, B.; Hillmer, S.; Zhao, M.; Lo, S.W.; Robinson, D.G. Identification of multivesicular bodies as prevacuolar compartments in Nicotiana tabacum BY-2 cells. Plant Cell 2004, 16, 672–693. [Google Scholar] [CrossRef] [PubMed]
- Vassilyev, A.E. Developmental and comparative ultrastructure of glandular hairs in the two Urticaceae. I. Urtica dioica. Nord. J. Bot. 1994, 14, 531–545. [Google Scholar] [CrossRef]
- Sacchetti, G.; Romagnoli, C.; Nicoletti, M.; Di Fabio, A.; Bruni, A.; Poli, F. Glandular trichomes of Calceolaria adscendens Lidl. (Scrophulariaceae): Histochemistry, development and ultrastructure. Ann. Bot. 1999, 83, 87–92. [Google Scholar] [CrossRef]
- Kim, E.S.; Mahlberg, P.G. Glandular cuticle formation in Cannabis (Cannabaceae). Am. J. Bot. 1995, 82, 1207–1214. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Zhou, C. Secretory Structures of Pogostemon auricularius: Morphology, Development, and Histochemistry. Symmetry 2019, 11, 13. https://doi.org/10.3390/sym11010013
Guo J, Zhou C. Secretory Structures of Pogostemon auricularius: Morphology, Development, and Histochemistry. Symmetry. 2019; 11(1):13. https://doi.org/10.3390/sym11010013
Chicago/Turabian StyleGuo, Jiansheng, and Cheng Zhou. 2019. "Secretory Structures of Pogostemon auricularius: Morphology, Development, and Histochemistry" Symmetry 11, no. 1: 13. https://doi.org/10.3390/sym11010013
APA StyleGuo, J., & Zhou, C. (2019). Secretory Structures of Pogostemon auricularius: Morphology, Development, and Histochemistry. Symmetry, 11(1), 13. https://doi.org/10.3390/sym11010013