
Controlling Wolbachia Transmission and Invasion Dynamics among Aedes Aegypti Population via Impulsive Control Strategy

 ,
, 
 , and
, and
Abstract
:1. Introduction
- Micro injection: In this process, Wolbachia strains are microinjected into aquatic stages such as eggs, larvae and pupae.
- Introgression: In this process, the Wolbachia strains are carried out to next generation through mating. If Wolbachia infected female mated with Wolbachia infected or uninfected male, then the produced offsprings have the Wolbachia strain (Called CI rescue). Suppose the Wolbachia uninfected female mated with a Wolbachia infected male then there is no viable progeny. Finally, if a non-Wolbachia female mated with a non-Wolbachia male then there is no Wolbachia infection in the offspring.
- A novel mathematical model, which considers the total of ten stages in Aedes Aegypti mosquitoes (combining both Wolbachia infected and Wolbachia uninfected) is proposed and the possible optimal stages to release the Wolbachia are discussed, and the most important concept of Wolbachia invasion and Wolbachia gain are adopted.
- The Wolbachia free equilibrium, Wolbachia present Equilibrium, Zero mosquitoes, and both Wolbachia and Non-Wolbachia mosquitoes co-existence equilibrium are derived. And utilizing fixed point theory results, the Existence and Uniqueness results of the Wolbachia invasive model are proposed. To attain optimal control, we utilized an impulsive control strategy.
- We perform global Mittag-Leffler stability analysis of the proposed model via Linear Matrix Inequality (LMI) theory and Lyapunov theory.
- In the end, by utilizing the data from the published literature, we have presented the numerical simulation of the proposed model using MATLAB software.
2. Preliminaries
- (1)
- There exists positive constants and
- (2)
- , , for any scalar .
- (3)
- , , .
3. Model Formulation
4. Equilibrium Points
4.1. Zero Mosquitoes
4.2. Wolbachia Infected Mosquitoes Free Equilibrium
4.3. Wild Mosquitoes Free Equilibrium
4.4. Both Wolbachia Infected Mosquitoes and Non-Wolbachia Mosquitoes Co-Existence Equilibrium
5. Wolbachia Invasion Model
6. Existence and Uniqueness of Solution
7. Stability Analysis
8. Numerical Simulation
- Case 1.
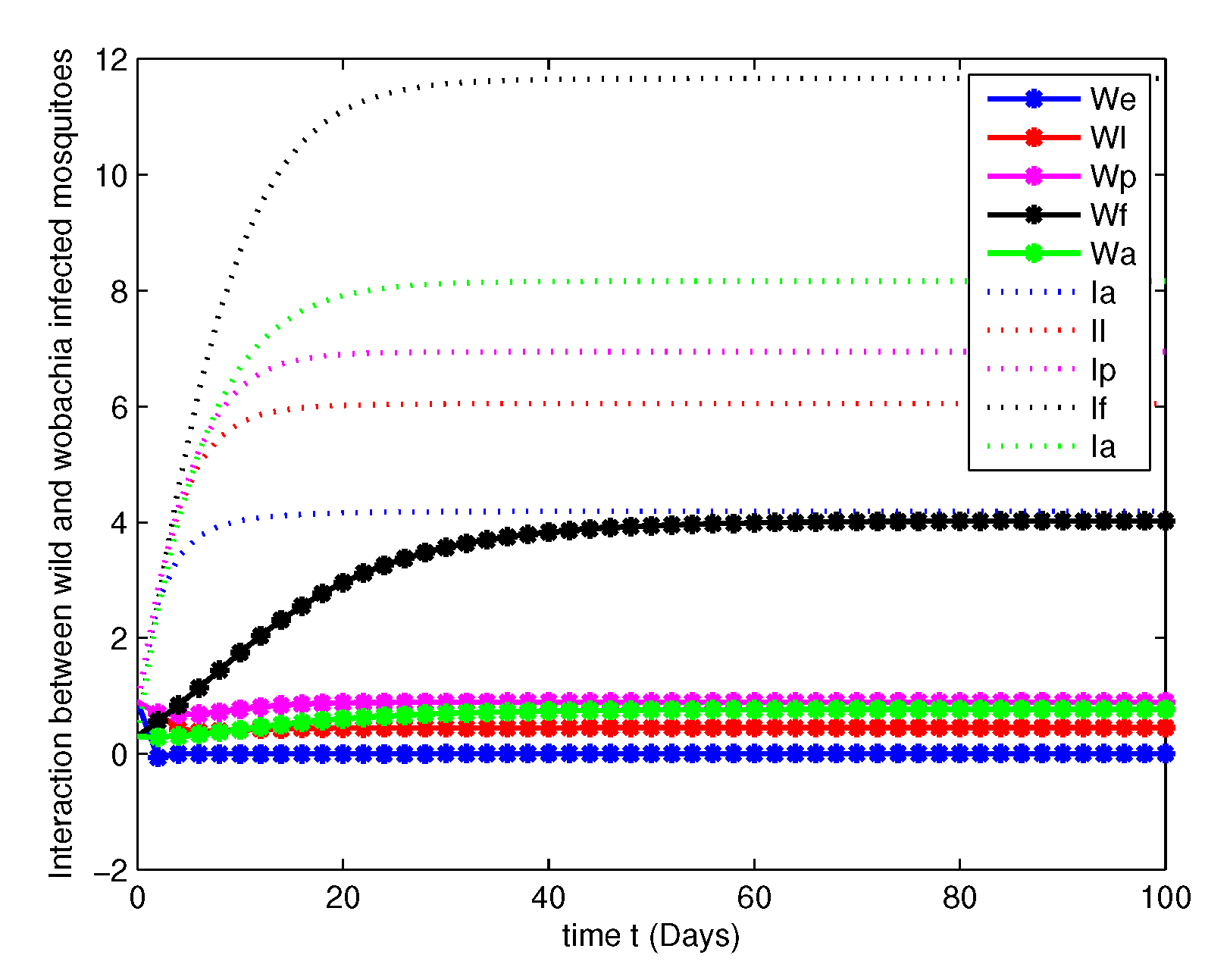
- In this case, we have analyzed the transmission dynamics of Wolbachia among Aedes Aegypti mosquitoes via substituting the values mentioned in Table 2.For this consider the system (5), with initial conditions , , , , , , , , , , total population , and the positive scalar used in Theorem 2 asThe Figure 4, Figure 5, Figure 6 and Figure 7 are depicts the dynamics of Equation (5) along with the parameters in Table 2 at various orders of such as and 1. We can observe by simulation results that, there is a notable decrease in non-Wolbachia mosquitoes and increase in Wolbachia infected mosquitoes.
- Case 2.
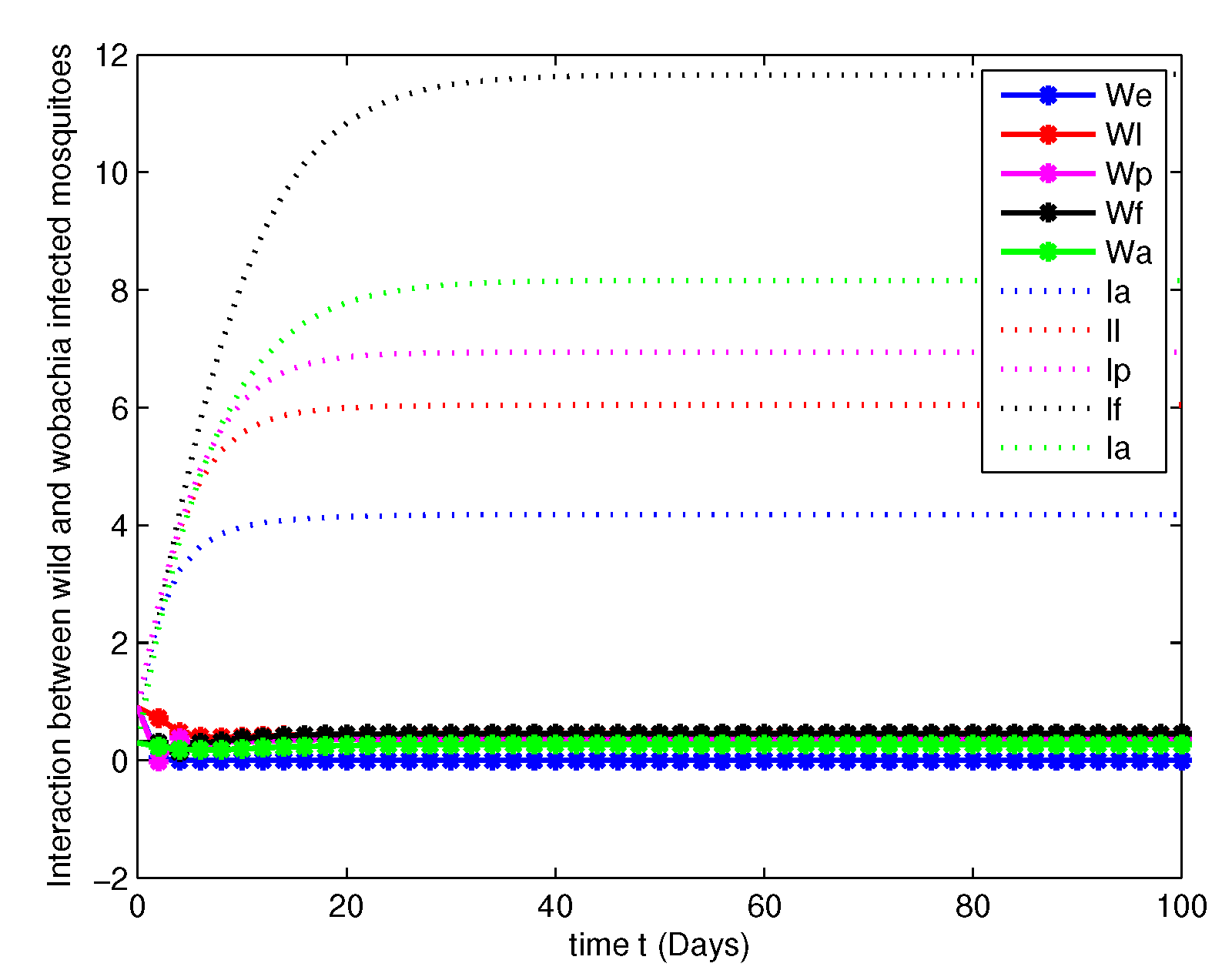
- In this case, we have analyzed the merits and demerits of considering the Wolbachia invasion. For this consider the system of Equation (6) with parameters mentioned in Table 2. We have plotted (6) with initial conditions and total population as considered in Case 1. Along with this, the other parameters , , , and are fitted.Figure 8, Figure 9, Figure 10 and Figure 11 are analyzed the dynamics of the system of Equation (6), with Wolbachia invasion and natural Wolbachia gain at various orders and 1. From this we can observe that, Wolbachia infected mosquitoes tends to annihilation before the eradication of non-Wolbachia mosquitoes. It will lead to the decay in natural CI rescue.
- Case 3.
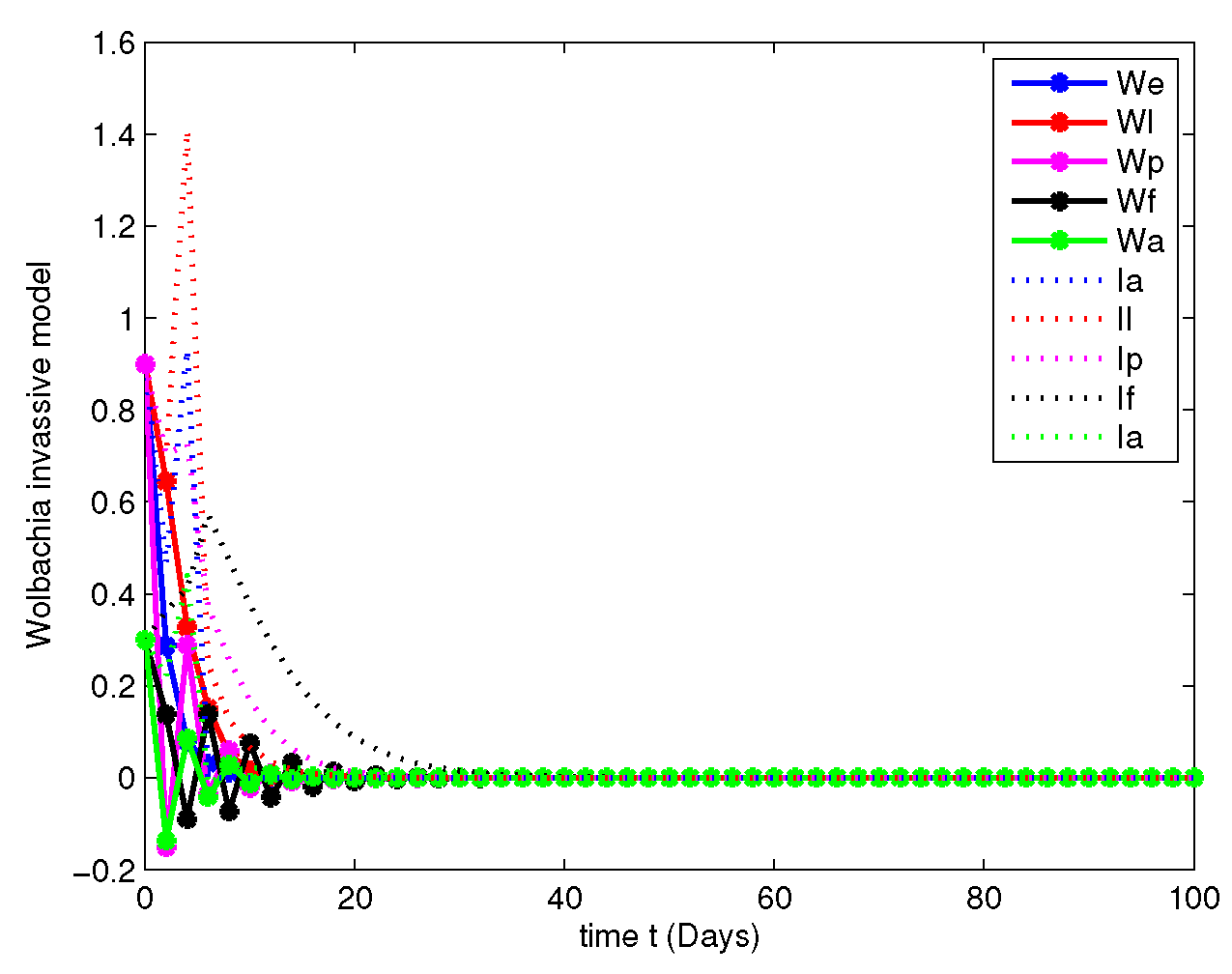
- In this case, the decay due to the natural Wolbachia invasion is managed by releasing Wolbachia infected mosquitoes impulsively. For this case, along with the parameters mentioned in Table 2, we have fitted the values of impulsive control as , , , and , invasion rates are , , , and gain rates are and .Figure 12, Figure 13, Figure 14 and Figure 15 explicitly shows the dynamics of the systems of Equation (7) with impulsive control at orders and 1. From this we get that, at order the system leads to instability, when the system started to posses stable state and at the both population are annihilated at initial stage compared with Figure 7 and Figure 11.
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Wolbachia Infected Mosquitoes Free Equilibrium
- (i).
- By solving,We get the value of as,
- (ii).
- By solvingWe get the value of as,Substitute the value of from (i),
- (iii).
- By solvingWe get the value of as,Substitute the value of from (ii),
- (iv).
- By solvingWe get the value of as,Substitute the value of from (ii),
- (v).
- By solvingWe get the value of as,Substitute the value of and from (iii) and (iv),
Appendix A.2. Wild Mosquitoes Free Equilibrium
- (i)
- By solvingWe get,
- (ii)
- By solvingWe get,
- (iii)
- By solvingWe get,
- (iv)
- By solvingWe get,Substitute the value of from (iii),
- (v)
- By solving,Put ,
Appendix A.3. Both Wolbachia and Non-Wolbachia Mosquitoes Co-Existence Equilibrium
- (i)
- (ii)
- (iii)
- Let
- (iv)
- Where, ;
- (v)
- Where,
- (vi)
- where,
- (vii)
- (viii)
- (ix)
- (x)
- The above equation is a quadratic equation on . That is,where,
References
- kilbas, A.A.; Sirvastava, H.M.; Trujillo, J.J. Theory and Applications of Fractional Differential Equations; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Rahimy, M. Applications of fractional differential equations. Appl. Math. Sci. 2010, 4, 2453–2461. [Google Scholar]
- Samko, S.G.; Kilbas, A.A.; Marichev, O.I. Fractional Integrals and Derivatives: Theory and Applications; Gordon & Breach, Science Publications: London, UK; New York, NY, USA, 1993. [Google Scholar]
- Podlubny, I. Fractional Differential Equations: An Introduction to Fractional Derivatives, Fractional Differential Equations, to Methods of Their Solution and Some of Their Applications; Academic Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Podlubny, I. Geometric and physical interpretation of fractional integration and fractional differentiation. Fract. Calc. Appl. Anal. 2002, 5, 367–386. [Google Scholar]
- Oldham, K.B.; Spanier, J. The Fractional Calculus; Academic Press: New York, NY, USA; London, UK, 1974. [Google Scholar]
- Li, Y.; Chen, Y.; Podlubny, I. Stability of fractional-order nonlinear dynamic systems: Lyapunov direct method and generalized Mittag-Leffler stability. Comput. Math. Appl. 2010, 59, 1810–1821. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, R.; Vaughn, D. Dengue: An escalating problem. BMJ 2002, 324, 1563–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Chye, J.K.; Lim, C.T.; Ng, K.B. Vertical transmission of dengue. Clin. Infect. Dis. 1997, 25, 1374–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.; Sinka, M.; Duda, K.; Mylne, A.; Shearer, F.; Barker, C. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, 1–18. [Google Scholar] [CrossRef]
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol. 2002, 10, 100–103. [Google Scholar] [CrossRef]
- Ong, A.; Sandar, M.; Chen, M.I.; Sin, L.Y. Fatal dengue hemorrhagic fever in adults during a dengue epidemic in Singapore. Int. J. Infect. Dis. 2007, 11, 263–267. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Vector-Borne Diseases. 2020. Available online: http://www.who.int/mediacentre/factsheets/fs387/en/ (accessed on 25 January 2021).
- Alphey, L.; Benedict, M.; Bellini, R.; Clark, G.G.; Dame, D.A.; Service, M.W.; Dobson, S.L. Sterile-insect methods for control of mosquito-borne diseases: An analysis. Vector-Borne Zoonotic Dis. 2010, 10, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, J.; Lefrancois, T. Boosting the sterile insect technique to control mosquitoes. Trends Parasitol. 2014, 30, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Lees, R.S.; Nimmo, D.; Aw, D.; Jin, L.; Gray, P.; Berendonk, T.U. Femalespecific flightless phenotype for mosquito control. Proc. Natl. Acad. Sci. USA 2010, 107, 4550–4554. [Google Scholar] [CrossRef] [Green Version]
- James, A.A. Gene drive systems in mosquitoes: Rules of the road. Trends Parasitol. 2005, 21, 64–67. [Google Scholar] [CrossRef]
- Scott, T.W.; Takken, W.; Knols, B.G.J.; Boëte, C. The ecology of genetically modified mosquitoes. Science 2002, 298, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Masud, M.A.; Kim, B.N.; Kim, Y. Optimal control problems of mosquito-borne disease subject to changes in feeding behaviour of Aedes mosquitoes. Biosystems 2017, 156–157, 23–39. [Google Scholar] [CrossRef]
- Momoh, A.A.; Fugenschuh, A. Optimal control of intervention strategies and cost effectiveness analysis for a zika virus model. Oper. Res. Health Care 2018, 18, 99–111. [Google Scholar] [CrossRef]
- Segoli, M.; Hoffmann, A.A.; Lloyd, J.; Omodei, G.J.; Ritchie, S.A. The effect of virus-blocking Wolbachia on male competitiveness of the dengue vector mosquito, Aedes aegypti. PLOS Negl. Trop. Dis. 2014, 8, e3294. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The WMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Xi, Z.; Khoo, C.C.; Dobson, S.L. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 2005, 310, 326–328. [Google Scholar] [CrossRef] [Green Version]
- Ormaetxe, I.; Walker, T.; Neill, S.L.O. Wolbachia and the biological control of mosquito-borne disease. Embo Rep. 2011, 12, 508–518. [Google Scholar] [CrossRef] [Green Version]
- World Mosquito Program. Available online: https://www.worldmosquitoprogram.org (accessed on 25 January 2021).
- Dutra, H.L.C.; Rocha, M.N.; Dias, F.B.S.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host Microbe 2016, 19, 771–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, P.; Sinkins, S.; Godfray, H. Population dynamic models of the spread of Wolbachia. Am. Nat. 2011, 177, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Hughes, H.; Britton, N. Modelling the use of Wolbachia to control dengue fever transmission. Bull. Math. Biol. 2013, 75, 796–818. [Google Scholar] [CrossRef] [Green Version]
- McMeniman, C.J.; Lane, R.V.; Cass, B.N.; Fong, A.W.; Sidhu, M.; Wang, Y.F.; Neill, S.L.O. Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science 2009, 323, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Jiggins, F. The spread of Wolbachia through mosquito populations. PLoS Biol. 2017, 15, e2002780. [Google Scholar] [CrossRef]
- Ndii, M.Z.; Hickson, R.I.; Allingham, D.; Mercer, G.N. Modelling the transmission dynamics of dengue in the presence of Wolbachia. Math. Biosci. 2015, 262, 157–166. [Google Scholar] [CrossRef]
- Koiller, J.; da Silva, M.A.; Souza, M.O.; Codeco, C.; Iggidr, A.; Sallet, G. Aedes, Wolbachia and Dengue; Inria Nancy-Grand Est: Villers-lès-Nancy, France, 2014; pp. 1–47. [Google Scholar]
- Adekunle, A.I.; Michael, M.T.; McBryde, E.S. Mathematical analysis of a Wolbachia invasive model with imperfect maternal transmission and loss of Wolbachia infection. Infect. Dis. Model. 2019, 4, 265–285. [Google Scholar] [CrossRef]
- Xue, L.; Manore, C.; Thongsripong, P.; Hyman, J. Two-sex mosquito model for the persistence of Wolbachia. J. Biol. Dyn. 2017, 11, 216–237. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.S.; Wooda, D.A.; Keeling, M.J. Age- and bite-structured models for vector-borne diseases. Epidemics 2015, 12, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafikov, M.; Meza, M.E.M.; Correa, D.P.F.; Wyse, A.P. Controlling Aedes aegypti populations by limited Wolbachia-based strategies in a seasonal environment. Math. Methods Appl. Sci. 2019, 42, 5736–5745. [Google Scholar] [CrossRef]
- Supriatna, A.K.; Anggriani, N.; Melanie; Husniah, H. The optimal strategy of Wolbachia- infected mosquitoes release program an application of control theory in controlling Dengue disease. In Proceedings of the 2016 International Conference on Instrumentation, Control and Automation(ICA), Bandung, Indonesia, 29–31 August 2016; pp. 38–43. [Google Scholar]
- Dianavinnarasi, J.; Cao, Y.; Raja, R.; Rajchakit, G.; Lim, C.P. Delay-dependent stability criteria of delayed positive systems with uncertain control inputs: Application in mosquito-borne morbidities control. Appl. Math. Comput. 2020, 382, 125210. [Google Scholar] [CrossRef]
- Dianavinnarasi, J.; Raja, R.; Alzabut, J.; Cao, J.; Niezabitowski, M.; Bagdasar, O. Application of Caputo—Fabrizio operator to suppress the Aedes Aegypti mosquitoes via Wolbachia: An LMI approach. Math. Comput. Simul. 2021. [Google Scholar] [CrossRef]
- Nisar, K.S.; Ahmad, S.; Ullah, A.; Shah, K.; Alrabaiah, H.; Arfan, M. Mathematical analysis of SIRD model of COVID-19 with Caputo fractional derivative based on real data. Results Phys. 2021, 21, 103772. [Google Scholar] [CrossRef]
- Boyd, S.; Ghaoui, L.; Feron, E.; Balakrishnan, V. Linear Matrix Inequalities in System and Control Theory; SIAM Philadelphia: Philadelphia, PA, USA, 1994. [Google Scholar]
- Wu, H.; Zhang, X.; Xue, S.; Wang, L.; Wang, Y. LMI conditions to global Mittag-Leffler stability of fractional-order neural networks with impulses. Neurocomputing 2016, 193, 148–154. [Google Scholar] [CrossRef]
- Stamova, I. Global stability of impulsive fractional differential equations. Appl. Math. Comput. 2014, 237, 605–612. [Google Scholar] [CrossRef]
- Agarwal, R.P.; Meehan, M.; O’Regan, D. Fixed Point Theory and Applications; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Iswarya, M.; Raja, R.; Rajchakit, G.; Alzabut, J.; Lim, C.P. A perspective on graph theory based stability analysis of impulsive stochastic recurrent neural networks with time-varying delays. Adv. Differ. Equ. 2019, 502, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Stamov, G.; Stamova, I.; Alzabut, J. Global exponential stability for a class of impulsive BAM neural networks with distributed delays. Appl. Math. Inf. Sci. 2013, 7, 1539–1546. [Google Scholar] [CrossRef]
- Stamov, G.T.; Alzabut, J.O.; Atanasov, P.; Stamov, A.G. Almost periodic solutions for impulsive delay model of price fluctuations in commodity markets. Nonlinear Anal. Real World Appl. 2011, 12, 3170–3176. [Google Scholar] [CrossRef]
- Zada, A.; Waheed, H.; Alzabut, J.; Wang, X. Existence and stability of impulsive coupled system of fractional integrodifferential equations. Demonstr. Math. 2019, 52, 296–335. [Google Scholar] [CrossRef]
- Zada, A.; Alam, L.; Kumam, P.; Kumam, W.; Ali, G.; Alzabut, J. Controllability of impulsive non-linear delay dynamic systems on time scale. IEEE Access 2020, 8, 93830–93839. [Google Scholar] [CrossRef]
- Ndii, M.Z.; Hickson, R.I.; Mercer, G.N. Modelling the introduction of Wolbachia into Aedes aegypti to reduce dengue transmission. Anziam J. 2012, 53, 213–227. [Google Scholar]
- Yang, H.M.; Macoris, M.L.G.; Galvani, K.C.; Andrighetti, M.T.M.; Wanderley, D.M.V. Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiol. Infect. 2009, 137, 1188–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maidana, N.A.; Yang, H.M. Describing the geographic spread of dengue disease by traveling waves. Math. Biosci. 2008, 215, 64–77. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description |
|---|---|
| , | Reproduction rate of non-Wolbachia mosquitoes |
| and Wolbachia infected mosquitoes respectively | |
| The natural death rate of eggs without Wolbachia infection | |
| The natural death of larvae without Wolbachia infection | |
| The natural death of pupae without Wolbachia infection | |
| The natural death of adult female mosquitoes without Wolbachia infection | |
| The natural death of adult male mosquitoes without Wolbachia infection | |
| The natural death of eggs with Wolbachia infection | |
| The natural death of larvae with Wolbachia infection | |
| The natural death of pupae with Wolbachia infection | |
| The natural death of adult female mosquitoes with Wolbachia infection | |
| The natural death of infected adult male mosquitoes with Wolbachia infection | |
| The rate at which the fraction of non-Wolbachia eggs matured into non-Wolbachia larvae | |
| The rate at which the fraction of non-Wolbachia larvae matured into non-Wolbachia pupae | |
| The rate at which the fraction of non-Wolbachia pupae matured into non-Wolbachia | |
| immature female or male | |
| The rate at which the fraction of the Wolbachia infected mosquito eggs | |
| matured into Wolbachia infected or uninfected larvae | |
| The rate at which the fraction of the Wolbachia infected mosquito larvae | |
| matured into Wolbachia infected or uninfected pupae | |
| The rate at which the fraction of the Wolbachia infected mosquito pupae | |
| matured into Wolbachia infected or uninfected adults | |
| The probability of having male or female mosquitoes |
| Parameters | Description | Data |
|---|---|---|
| Reproduction rate of Wolbachia uninfected mosquitoes | 1.25/day [52] | |
| , , | The death rate of aquatic Wolbachia uninfected mosquitoes | /day [53] |
| The Maturation rate of Wolbachia uninfected mosquitoes | /day [54] | |
| The death rate of adult Wolbachia uninfected mosquitoes | /day [53] | |
| The death rate of aquatic Wolbachia infected mosquitoes | /day [53] | |
| The death rate of adult Wolbachia infected mosquitoes | /day [24] | |
| Reproduction rate of Wolbachia infected mosquitoes | [52] | |
| The maturation rate of Wolbachia infected mosquitoes | /day [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dianavinnarasi, J.; Raja, R.; Alzabut, J.; Niezabitowski, M.; Bagdasar, O. Controlling Wolbachia Transmission and Invasion Dynamics among Aedes Aegypti Population via Impulsive Control Strategy. Symmetry 2021, 13, 434. https://doi.org/10.3390/sym13030434
Dianavinnarasi J, Raja R, Alzabut J, Niezabitowski M, Bagdasar O. Controlling Wolbachia Transmission and Invasion Dynamics among Aedes Aegypti Population via Impulsive Control Strategy. Symmetry. 2021; 13(3):434. https://doi.org/10.3390/sym13030434
Chicago/Turabian StyleDianavinnarasi, Joseph, Ramachandran Raja, Jehad Alzabut, Michał Niezabitowski, and Ovidiu Bagdasar. 2021. "Controlling Wolbachia Transmission and Invasion Dynamics among Aedes Aegypti Population via Impulsive Control Strategy" Symmetry 13, no. 3: 434. https://doi.org/10.3390/sym13030434
APA StyleDianavinnarasi, J., Raja, R., Alzabut, J., Niezabitowski, M., & Bagdasar, O. (2021). Controlling Wolbachia Transmission and Invasion Dynamics among Aedes Aegypti Population via Impulsive Control Strategy. Symmetry, 13(3), 434. https://doi.org/10.3390/sym13030434