
Functional Connectivity in Developmental Dyslexia during Speed Discrimination



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Paradigm
2.3. EEG Signal Pre-Processing
2.4. Small-World Propensity
2.5. Statistical Analysis of Global SWP Measures
2.6. Statistical Analysis of Local SWP Measures
2.7. Statistical Analysis of Behavior Parameters
3. Results
3.1. Behavior Measures
3.2. Impact on Motion Measures on Global SWP Measures
3.3. Impact on Motion Measures on the Strength and Betweenness Centrality of Nodes
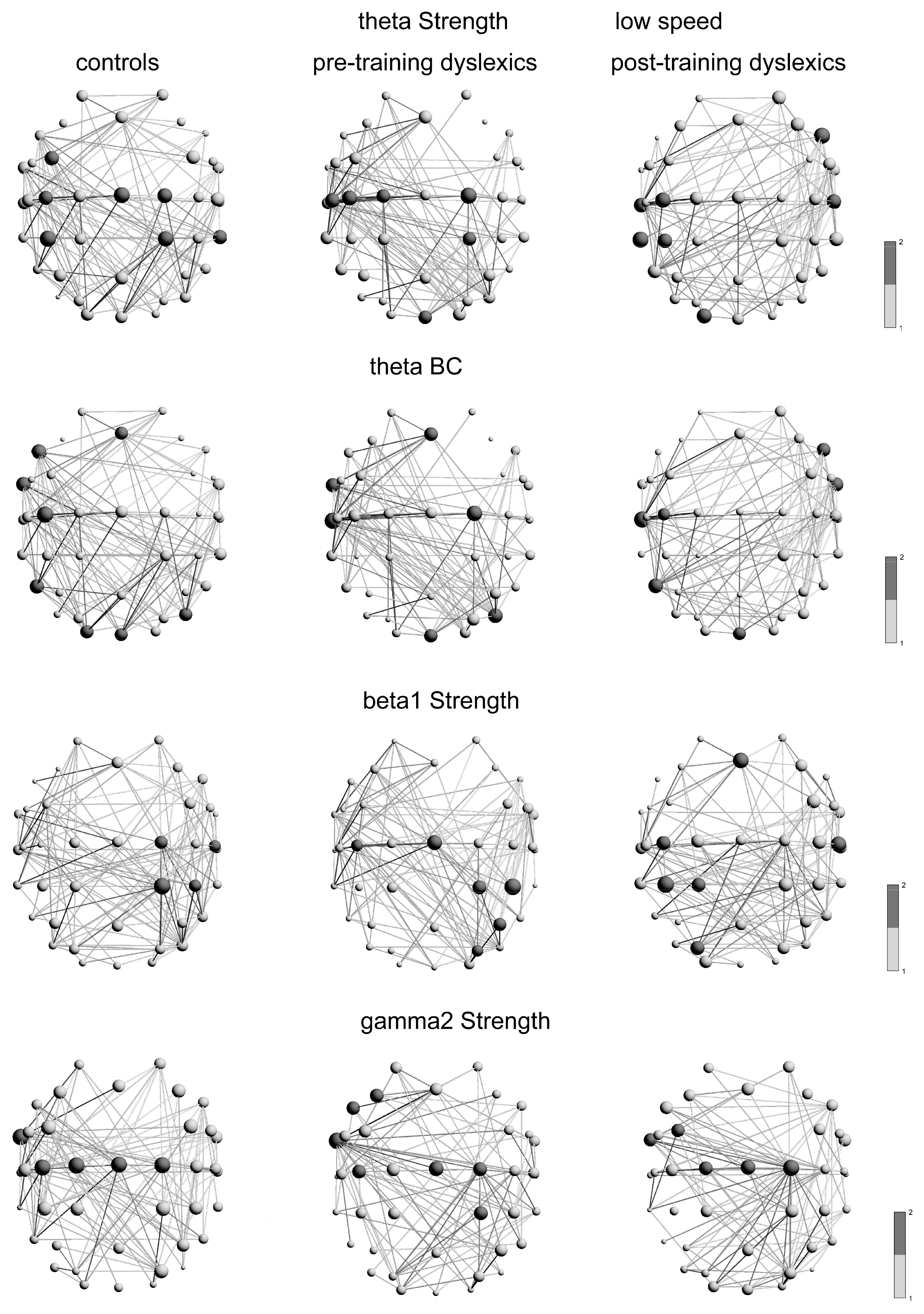
3.3.1. Hub Distributions in the Graphs at Low-Speed Contrast
3.3.2. Hub Distributions at High-Speed Contrast
4. Discussion
4.1. Global Network Measures
4.2. Local Network Measures
4.3. Local FEF-PEF Network Measures
4.4. Constraints
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
| 1. DDE-2 Test | Dyslexics mean ± s.d | Controls mean ± s.d |
| 1.1. Word reading | ||
| Accuracy time | 90.6 ± 4.5 91.7 ± 4.8 | 106 ± 5.58 132 ± 0.76 |
| 1.2. Pseudo-word reading | ||
| Accuracy time | 88.3 ± 4.4 96.7 ± 4.1 | 102 ± 4.65 118 ± 0.68 |
| 1.3. Homonyms Accuracy | 98.5 ± 2.4 | 112 ± 4.82 |
| 1.4. Spelling Accuracy | 112 ± 4.45 | 112 ± 1.82 |
| 1.5. Word writing Accuracy | 85.7 ± 4.4 | 115 ± 6.49 |
| 1.6. Pseudo-word writing Accuracy | 91.9 ± 3.9 | 104 ± 4.25 |
| 1.7. Dictation Accuracy | 89.5 ± 3.4 | 112 ± 4.82 |
| 2. Psychometric tests | Dyslexics mean ± s.d | Control mean ± s.d |
| 2.1. Dictation | ||
| Correct sentences | 10.94 ± 4.51 | 21.00 ± 5.85 |
| 2.2. Text reading | ||
| Correct answers | 119.75 ± 7.88 | 129.41 ± 3.43 |
| time (s) | 191.56 ± 149.55 | 104.77 ± 29.00 |
| 2.3. Phological task Without the first sound | ||
| Correct | 5.52 ± 2.04 | 9.20 ± 1.87 |
| time (s) | 62.5 ± 29.4 | 34.86 ± 10.57 |
| 2.4. Phological task Without the last syllable | ||
| Correct answers | 6.15 ± 2.38 | 8.05 ± 2.08 |
| time | 64.84 ± 30.5 | 37.50 ± 8.81 |
| 3. Girolami-Bolinier | ||
| Correct answers | 50.9 ± 9.2 | |
| 4. Raven test | >98 | >98 |
| Frequency/Metric | Control | Pre-Dys | Post-Dys | Con/Pre-Dys | Con/Post-Dys | Pre-/Post-Dys | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| hubs | hubs | hubs | p | χ2 | p | χ2 | p | χ2 | ||
| δ | Str | FT9, C3, C5, Pz, P08, O2 | FC3-4, F4, C4, C5, T7, CP2, CP3, PO7 | AF3, F7, FC5, FT10, T7, C3, CP1 | 0.410 | 0.67 | 0.496 | 0.46 | 0.957 | 0.002 |
| BC | F7, FT9, C3, C5, P3 | F7, FC3, FT9-10, C3, T7, PO7 | FC5, C1, T7, AF4, FT10, O2 | 0.726 | 0.12 | 0.466 | 0.53 | 0.648 | 0.20 | |
| θ | Str | FC3, C2, C3, Cz, CP2, CP3, TP8 | C1-2, C3, C5, CP2, Oz | F8, C3, T7, TP7, C6, CP3, O1 | 0.022 | 5.19 | 0.640 | 0.21 | 0.015 | 5.88 |
| BC | Fz, F7, FT9, C3, P7, P08, O1, Oz | Fz, FT9, T7, C2, P08, Oz | F8, FT10, C3, T7, P7, Oz | 0.223 | 1.48 | 0.125 | 2.34 | 0.009 | 6.63 | |
| α | Str | Fz, Cz, Pz, CP3, PO7, PO4 | Fz, Pz, P08, P3, FT10, C4, P8 | Fz, Cz, AF4, CP4, P4, P08 | 0.229 | 1.44 | 0.371 | 0.79 | 0.060 | 3.53 |
| BC | Fz, Cz, P08, T7, TP7, PO4, O2 | FT9, Fz, Pz, P08, T7 FT10, P8 | FT9, Cz, P08, PO3 P4, P8 | 0.188 | 1.73 | 0.834 | 0.04 | 0.359 | 0.83 | |
| β1 | Str | C2, C6, CP2, CP4 | C3, Cz, CP2, CP4, P4, PO4 | Fz, C3, C6, CP1, CP3, PO3 | 0.081 | 3.02 | 0.257 | 1.28 | 0.004 | 7.91 |
| BC | T7, C2, CP2, P8, PO8 | FT9-10, C3, C5, Cz, PO4, P08, O2 | Fz, FT10, T7, C2, C6, CP3, PO3, P8, O1 | 0.131 | 2.27 | 0.447 | 0.57 | 0.028 | 4.79 | |
| β2 | Str | AF3-4, Fz, FC4, CP2, CP3-4, P4 | FC6, C2, C4, C6, CP2, CP4, PO4 | AF3, FC4, C4, C6, CP2, CP4, P4 | 0.378 | 0.77 | 0.398 | 0.71 | 0.906 | 0.01 |
| BC | AF3-4, Fz, TP7, C2, C6, CP2, CP4, P3-4, PO4 | AF3, Fz, FT10, C4, CP4, PO4, PO7 | AF3, Fz, F8, P8, PO4 | 0.233 | 1.41 | 0.209 | 1.57 | 0.802 | 0.06 | |
| γ1 | Str | C2, C4, C6, CP2, CP4, P4 | AF4, F4, FC4, C4, C6, CP2, CP4, P4, P8 | AF3, CP1, C4, C6, CP2, P4 | 0.310 | 1.03 | 0.431 | 0.61 | 0.877 | 0.02 |
| BC | T7, C2, C6, T8, P4 | FT9, T7, C2, CP2, P8 | AF3-4, FC6, C1, CP1, T7, C4, C6,CP2, TP8 | 0.970 | 0.001 | 0.971 | 0.001 | 0.987 | 0.0002 | |
| γ2 | Str | FT9, C1-2, Cz, C3 | F3, F7, FT9, Cz, C2, C3, CP2 | FC3, FT9, C1-2, Cz | 0.0006 | 11.73 | 0.613 | 0.25 | 6.12 × 10−5 | 16.06 |
| BC | FT9-10, C3, C5, T8, CP2, PO4 | FT9, C2 | FT9, F8, C2, C4, PO4, P8 | 0.330 | 0.947 | 0.831 | 0.04 | 0.256 | 1.28 | |
| Frequency/Metric | Control | Pre-Dys | Post-Dys | Con/Pre-Dys | Con/Post-Dys | Pre-/Post-Dys | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| hubs | hubs | hubs | p | χ2 | p | χ2 | p | χ2 | ||
| δ | Str | AF3, FT9-10, CP1, T8, P8, O2 | Fz, F8, FC6, CP2, CP4, P4 | FT9, C1, Pz, CP3, AF4, FC6, C2 | 0.278 | 1.17 | 0.337 | 0.91 | 0.969 | 0.001 |
| BC | F3, F8, C1, Cz | FT9, F8, C1, P07-08, T7, PO3, CP2, O1 | FT9-10, F8, C5 | 0.090 | 2.86 | 0.607 | 0.26 | 0.220 | 1.50 | |
| θ | Str | F3, Fz, FC4, C1, Cz, CP4 | AF4, FC5, FT9, C3-4, C1, Cz, P08, Pz | F3, FC3, F8, TP7, C3, CP1, Pz | 0.011 | 6.33 | 0.636 | 0.22 | 0.002 | 9.47 |
| BC | F3, FT10, Fz, C1-2, P08 | FT9-10, F8, C3-4, C5, P08, Oz | Fz, FT10, C3, CP1, TP7, T8, Pz | 0.226 | 1.46 | 0.813 | 0.05 | 0.306 | 1.04 | |
| α | Str | Cz, C4, CP2, Pz, P4, PO4 | AF4, C5-6, C4, P8 | Fz, FT10, C3, C5, C1, Pz, T7, TP7 | 0.907 | 0.01 | 0.365 | 0.81 | 0.256 | 1.28 |
| BC | AF3, C3, Fz, C1, Cz, P08, T7, PO4 | AF4, Fz, P08, C6, Oz | Fz, FT10, T7, TP7 | 0.044 | 4.05 | 0.056 | 3.65 | 0.929 | 0.007 | |
| β1 | Str | FC3, C2, C3-4, FC4, C6, CP2, CP4, Pz, P4 | C3, C5, FC6, FT10, C2, T8, CP2, P4 | Fz, C1, CP1-2, P4 | 0.878 | 0.02 | 0.549 | 0.35 | 0.494 | 0.466 |
| BC | F7, FC3, C3, Fz, P08, C6, PO4 | FT10, C2, C3, C5, P7 | FT9, Fz, C1, T7, PO4, P08, O2 | 0.850 | 0.03 | 0.935 | 0.01 | 0.671 | 0.179 | |
| β2 | Str | FT10, C2, C4, CP2, P4 | Fz, FC4, C6, CP1-2 | AF4, FC3, FC6, C5, C4, PO4 | 0.025 | 5.02 | 0.004 | 8.16 | 0.427 | 0.630 |
| BC | AF3, FT10, T8, P08, PO4 | AF3, F3, C3, Fz, T7, F8, FC6, CP2 | C5, P7, C4, PO4 | 0.394 | 0.72 | 0.243 | 1.36 | 0.729 | 0.119 | |
| γ1 | Str | AF4, FT10, C4, C6, CP2, CP4 | C3-4, FC4,C6, CP2, CP4, TP8, PO4 | C3, C1, C6, CP2, P4 | 0.491 | 0.47 | 0.693 | 0.15 | 0.772 | 0.083 |
| BC | AF4, FT10, C5, CP2, P7 | AF3, C2, T8, TP8, PO4, P08, O1 | AF3-4, FC4, C1, T7-8, C6, P08 | 0.432 | 0.61 | 0.814 | 0.05 | 0.566 | 0.329 | |
| γ2 | Str | F3, FC3-4, FT9, C1, C3, Cz | F3, FT9, C2, C3, Cz, CP2 | F3, FC3, FT9-10, Cz, CP2, CP4, PO4 | 4.8 × 10−6 | 20.89 | 0.866 | 0.02 | 2.3 × 10−6 | 22.36 |
| BC | F7, FT10, C5, T7, PO4, Oz | FT9-10, C1-2, Cz | FT9, C2, C3, CP4 | 0.186 | 1.74 | 0.039 | 4.21 | 0.0009 | 10.99 | |
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Livingstone, M.S.; Rosen, G.D.; Drislane, F.W.; Galaburda, A.M. Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia. Proc. Natl. Acad. Sci. USA 1991, 88, 7943–7947. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.; Pammer, K.; Vidyasagar, T.R. A temporal sampling basis for visual processing in developmental dyslexia. Front. Hum. Neurosci. 2020, 14, 213. [Google Scholar] [CrossRef] [PubMed]
- Kristen, P. Temporal sampling in vision and the implications for dyslexia. Front. Hum. Neurosci. 2013, 7, 933. [Google Scholar]
- Rima, S.; Schmid, M.C. V1-bypassing thalamo-cortical visual circuits in blindsight and developmental dyslexia. Curr. Opin. Physiol. 2020, 16, 14–20. [Google Scholar] [CrossRef]
- Demb, J.B.; Boynton, G.M.; Best, M.; Heeger, D.J. Psychophysical evidence for a magnocellular pathway deficit in dyslexia. Vis. Res. 1998, 38, 1555–1559. [Google Scholar] [CrossRef] [Green Version]
- Ramus, F.; Rosen, S.; Dakin, S.C.; Day, B.L.; Castellote, J.M.; White, S.; Frit, U. Theories of developmental dyslexia: Insights from a multiple case study of dyslexic adults. Brain 2003, 126, 841–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rima, S.; Kerbyson, G.; Jones, E.; Schmid, M.C. Advantage of detecting visual events in the right hemifield is affected by reading skill. Vis. Res. 2020, 169, 41–48. [Google Scholar] [CrossRef]
- Wilmer, J.B.; Richardson, A.J.; Chen, Y.; Stein J., F. Two visual motion processing deficits in developmental dyslexia associated with different reading skills deficits. J. Cogn. Neurosci. 2004, 16, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Stein, J. Dyslexia: The role of vision and visual attention. Curr. Dev. Disord. Rep. 2014, 1, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Vidyasagar T, R.; Pammer, K. Impaired visual search in dyslexia relates to the role of the magnocellular pathway in attention. Neuroreport 1999, 10, 1283–1287. [Google Scholar] [CrossRef]
- Iles, J.; Walsh, V.; Richardson, A. Visual search performance in dyslexia. Dyslexia 2000, 6, 163–177. [Google Scholar] [CrossRef]
- Vidyasagar, T.R. Reading into neuronal oscillations in the visual system: Implications for developmental dyslexia. Front. Hum. Neurosci. 2013, 7, 811. [Google Scholar] [CrossRef] [Green Version]
- Brannan, J.R.; Williams, M.C. Allocation of visual attention in good and poor readers. Percept. Psychophys. 1987, 41, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Geiger, G.; Lettvin, J.Y.; Fahle, M. Dyslexic children learn a new visual strategy for reading: A controlled experiment. Vis. Res. 1994, 34, 1223–1233. [Google Scholar] [CrossRef]
- Richards, G.P.; Samuels, S.J.; Turnure, J.E.; Ysseldyke, J.E. Sustained and selective attention in children with learning disabilities. J. Learn. Disabil. 1990, 23, 129–136. [Google Scholar] [CrossRef]
- Casco, C.; Tressoldi, P.; Dellantonio, A. Visual selective attention and reading efficiency are related in children. Cortex 1998, 34, 531–546. [Google Scholar] [CrossRef]
- Facoetti, A.; Paganoni, P.; Turatto, M.; Marzola, V.; Mascetti, G.G. Visuospatial Attention in developmental dyslexia. Cortex 2000, 36, 109–123. [Google Scholar] [CrossRef]
- Lachmann, T.; Schmitt, A.; Braet, W.; van Leeuwen, C. Letters in the forest: Global precedence TEMPeffect disappears for letters but not for non-letters under reading-like conditions. Front. Psychol. 2014, 5, 705. [Google Scholar] [CrossRef] [Green Version]
- Lachmann, T.; van Leeuwen, C. Reading as functional coordination: Not recycling but a novel synthesis. Front. Psychol. 2014, 5, 1046. [Google Scholar] [CrossRef] [Green Version]
- Förster, J.; Dannenberg, L. GLOMOsys: A systems account of global versus local processing. Psychol. Inq. 2010, 21, 175–197. [Google Scholar] [CrossRef]
- Von Karolyi, C. Visual-spatial strength in dyslexia: Rapid discrimination of impossible figures. J. Learn. Disabil. 2001, 34, 380–391. [Google Scholar] [CrossRef]
- Shany, M.; Breznitz, Z. Rate- and accuracy-disabled subtype profiles among adults with dyslexia in the Hebrew orthography. Dev. Neuropsychol. 2011, 36, 889–913. [Google Scholar] [CrossRef]
- Glodstein-Marcusohn, Y.; Glodfarb, L.; Shany, M. Global and Local Visual Processing in Rate/Accuracy Subtypes of Dyslexia. Front. Psychol. 2020, 11, 828. [Google Scholar]
- Higuchi, H.; Iwaki, S.; Uno, A. Altered visual character and object recognition in Japanese-speaking adolescents with developmental dyslexia. Neurosci. Lett. 2020, 723, 134841. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, L.; Pouretemad, H.; Khatibi, A.; Stein, J. Magnocellular based visual motion training improves reading in Persian. Sci. Rep. 2019, 9, 1142. [Google Scholar] [CrossRef] [PubMed]
- Chouake, T.; Levy, T.; Javitt, D.C.; Lavidor, M. Magnocellular training improves visual word recognition. Front. Hum. Neurosci. 2012, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton, T. Improving dorsal stream function in dyslexics by training figure/ground motion discrimination improves attention, reading fluency, and working memory. Front. Hum. Neurosci. 2016, 10, 397. [Google Scholar] [CrossRef] [Green Version]
- Lawton, T.; Shelley-Tremblay, J. Training on movement figure-ground discrimination remediates low-level visual timing deficits in the dorsal stream, improving high-level cognitive functioning, including attention, reading fluency, and working memory. Front. Hum. Neurosci. 2017, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Magosso, E.; Cuppini, C.; Bertini, C. Audiovisual rehabilitation in hemianopia: A model-based theoretical investigation. Front. Hum. Neurosci. 2017, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Cuppini, C.; Ursino, M.; Magosso, E.; Ross, L.A.; Foxe, J.J.; Molholm, S. A computational analysis of neural mechanisms underlying the maturation of multisensory speech integration in neurotypical children and those on the autism spectrum. Front. Hum. Neurosci. 2017, 11, 518. [Google Scholar] [CrossRef] [Green Version]
- Dekker, T.M.; Ban, H.; van der Velde, B.; Sereno, M.I.; Welchman, A.E.; Nardini, M. Late development of cue integration is linked to sensory fusion in cortex. Curr. Biol. 2015, 25, 2856–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ursino, M.; Cuppini, C.; Magosso, E. A neural network for learning the meaning of objects and words from a featural representation. Neural Netw. 2015, 63, 234–253. [Google Scholar] [CrossRef] [PubMed]
- Culham, J.; He, S.; Dukelow, S.; Verstraten, F.A. Visual motion and the human brain: What has neuroimaging told us? Acta Psychol. 2001, 107, 69–94. [Google Scholar] [CrossRef]
- Braddick, O.J.; O’Brien, J.M.; Wattam-Bell, J.; Atkinson, J.; Hartley, T.; Turner, R. Brain areas sensitive to coherent visual motion. Perception 2001, 30, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Murakami, I.; Cavanagh, P. Visual jitter: Evidence for visual-motion-based compensation of retinal slip due to small eye movements. Vis. Res. 2001, 41, 173–186. [Google Scholar] [CrossRef] [Green Version]
- Demb, J.B.; Boynton, G.M.; Heeger, D.J. Functional magnetic resonance imaging of early visual pathways in dyslexia. J. Neurosci. 1998, 18, 6939–6951. [Google Scholar] [CrossRef] [PubMed]
- Greenlee, M.W.; Lang, H.-J.; Mergner, T.; Seeger, W. Visual short-term memory of stimulus velocity in patients with unilateral posterior brain damage. J. Neurosci. 1995, 15, 2287–2300. [Google Scholar] [CrossRef]
- Vaina, L.M. Selective impairment of visual motion interpretation following lesions of the right occipito-parietal area in humans. Biol. Cybern. 1989, 61, 347–359. [Google Scholar] [CrossRef]
- Corbetta, M.F.; Miezin, M.; Dobmeyer, S.; Shulman, G.L.; Petersen, S.E. Selective and divided attention during visual discriminations of shape, color, and speed: Functional anatomy by positron emission tomography. J. Neurosci. 1991, 11, 2383–2402. [Google Scholar] [CrossRef] [PubMed]
- Morrone, M.C.; Tosetti, M.; Montanaro, D.; Fiorentini, A.; Cioni, G.; Burr, D.C. A cortical area that responds specifically to optic flow, revealed by fMRI. Nat. Neurosci. 2000, 3, 1322–1328. [Google Scholar] [CrossRef]
- Sunaert, S.; Van Hecke, P.; Marchal, G.; Orban, G.A. Attention to speed of motion, speed discrimination, and task difficulty: An fMRI study. NeuroImage 2000, 11, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Pitzalis, S.; Serra, C.; Sulpizio, V.; Committeri, G.; de Pasquale, F.; Fattori, P.; Galletti, C.; Sepe, R.; Galati, G. Neural bases of self- and object-motion in a naturalistic vision. Hum. Brain Mapp. 2020, 41, 1084–1111. [Google Scholar] [CrossRef] [Green Version]
- Warren, P.A.; Rushton, S.K. Optic flow processing for the assessment of object movement during ego movement. Curr. Biol. 2009, 19, 1555–1560. [Google Scholar] [CrossRef]
- Warren, P.A.; Rushton, S.K. Perception of scene-relative object movement: Optic flow parsing and the contribution of monocular depth cues. Vis. Res. 2009, 49, 1406–1419. [Google Scholar] [CrossRef]
- Tootell, R.B.; Reppas, J.B.; Kwong, K.K.; Malach, R.; Born, R.T.; Brady, T.J.; Rosen, B.R.; Belliveau, J.W. Functional analysis of human MT and related visual cortical areas using magnetic resonance imaging. J. Neurosci. 1995, 15, 3215–3230. [Google Scholar] [CrossRef]
- Pitzalis, S.; Sdoia, S.; Bultrini, A.; Committeri, G.; Di Russo, F.; Fattori, P.; Galati, G. Selectivity to translational egomotion in human brain motion areas. PLoS ONE 2013, 8, e60241. [Google Scholar] [CrossRef] [Green Version]
- Sereno, M.I.; Huang, R.S. A human parietal face area contains aligned head-centered visual and tactile maps. Nat. Neurosci. 2006, 9, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Cardin, V.; Smith, A.T. Sensitivity of human visual and vestibular cortical regions to egomotion-compatible visual stimulation. Cereb. Cortex 2010, 20, 1964–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitzalis, S.; Sereno, M.I.; Committeri, G.; Fattori, P.; Galati, G.; Patria, F.; Galletti, C. Human V6: The medial motion area. Cereb. Cortex 2010, 20, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, C.; Galletti, C.; Di Marco, S.; Fattori, P.; Galati, G.; Sulpizio, V.; Pitzalis, S. Egomotion-related visual areas respond to active leg movements. Hum. Brain Mapp. 2019, 40, 3174–3191. [Google Scholar] [CrossRef] [Green Version]
- Galletti, C.; Fattori, P. Neuronal mechanisms for detection of motion in the field of view. Neuropsychologia 2003, 41, 1717–1727. [Google Scholar] [CrossRef]
- Fischer, E.; Bülthoff, H.H.; Logothetis, N.K.; Bartels, A. Visual motion responses in the posterior cingulate sulcus: A comparison to V5/MT and MST. Cereb. Cortex 2012, 22, 865–876. [Google Scholar] [CrossRef]
- De Jong, B.M.; Shipp, S.; Skidmore, B.; Frackowiak, R.S.J.; Zeki, S. The cerebral activity related to the visual perception of forward motion in depth. Brain 1994, 117, 1039–1054. [Google Scholar] [CrossRef] [Green Version]
- Vilhelmsen, K.; van der Weel, F.R.R.; van der Weer, A.L.H. A high-density EEG study of differences between three high speeds of simulated forward motion from optic flow in adult participants. Front. Syst. Neurosci. 2015, 9, 146. [Google Scholar] [CrossRef] [Green Version]
- Wandell, B.A.; Winawer, J. Imaging retinotopic maps in the human brain. Vis. Res. 2011, 51, 718–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremmer, F.; Schlack, A.; Shah, N.J.; Zafiris, O.; Kubischik, M.; Hoffmann, K.P.; Fink, G.R. Polymodal motion processing in posterior parietal and premotor cortex. Neuron 2001, 29, 287–296. [Google Scholar] [CrossRef]
- Taskov, T.; Dushanova, J. Reading-network in developmental dyslexia before and after visual training. Symmetry 2020, 12, 1842. [Google Scholar] [CrossRef]
- Raichev, P.; Geleva, T.; Valcheva, M.; Rasheva, M.; Raicheva, M. Chapter: Protokol za nevrologichno I nevropsihologichno izslenvane na detsa sas spetsifichni obuchitelni zatrudneniya. In Protocol on Neurological and Neuropsychological Studies of Children with Specific Learning Disabilities. Integrated Learning and Resource Teacher; Evgenieva, E., Ed.; Publishing House “Dr. Ivan Bogorov”: Sofia, Bulgaria, 2005; pp. 82–105. (In Bulgarian) [Google Scholar]
- Matanova, V.; Todorova, E. DDE-2 Testova bateriya za otsenka na disleksiya na razvitieto—Balgarska adaptatsiya. In DDE-2 Test. Battery for Evaluation of Dyslexia of Development; Sartori, G., Remo, J., Tressoldi, P.E., Matanova, V., Todorova, E., Kolev, M., Eds.; OS Bulgaria Ltd.: Sofia, Bulgaria; Available online: https://www.giuntipsy.bg/bg/prod-19-testova-bateriq-za-ocenka-na-disleksiq-na-razvitieto.htm (accessed on 1 October 2013). (In Bulgarian)
- Sartori, G.; Remo, J.; Tressoldi, P.E. Updated and revised edition for the evaluation of dyslexia. In DDE-2, Battery for the Developmental Dyslexia and Evolutionary Disorders-2, 1995; Giunti, O.S.: Florence, Italy, 2007. [Google Scholar]
- Kalonkina, A.; Lalova, Y. Normative indicators for the test battery for a written speech assessment. In Logopedical Diagnostics (30-38); Tyubele, S., Iossifova, R., Eds.; Rommel Publishing House: Sofia, Bulgaria, 2016. [Google Scholar]
- Raven, J.; Raven, J.C.; Court, J.H. Manual for Raven’s Progressive Matrices and Vocabulary Scales. Section 2: The Coloured Progressive Matrices; Oxford Psychologists Press: Oxford, UK, 1998. [Google Scholar]
- Girolami-Boulinier, A. Contrôle des Aptitudes à la Lecture et àl’Ecriture (CALE); CALE: Paris, Masson, 1985. (In French) [Google Scholar]
- Yakimova, R. Narusheniya na Pismenata rech. Abnormalities of Written Speech; Rommel Publishing House: Sofia, Bulgaria, 2004. (In Bulgarian) [Google Scholar]
- Sun, D.; Roth, S.; Black, M.J. A quantitative analysis of current practices in optical flow estimation and the principles behind them. Int. J. Comput. Vis. 2014, 106, 115–137. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Roth, S.; Black, M.J. Secrets of optical flow estimation and their principles. In Proceedings of the 2010 IEEE Computer Society Conference on Computer Vision and Pattern Recognition, San Francisco, CA, USA, 13–18 June 2010; pp. 2432–2439. [Google Scholar]
- Eden, G.; Van Meter, J.; Rumsey, J.; Maisog, J.M.; Woods, R.P.; Zeffiro, T.A. Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature 1996, 382, 66–69. [Google Scholar] [CrossRef]
- Joshi, M.R.; Falkenberg, H.K. Development of radial optic flow pattern sensitivity at different speeds. Vis. Res. 2015, 110, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Dushanova, J.; Christov, M. Auditory event-related brain potentials for an early discrimination between normal and pathological brain aging. Neural Reg. Res. 2013, 8, 1390–1399. [Google Scholar]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; vanWingerden, M.; Battaglia, F.; Pennartz, C.M.A. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. NeuroImage 2011, 55, 1548–1565. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.R.; D’Esposito, M. The segregation and integration of distinct brain networks and their relationship to cognition. J. Neurosci. 2016, 36, 12083–12094. [Google Scholar] [CrossRef] [Green Version]
- Onnela, J.P.; Saramäki, J.; Kertész, J.; Kaski, K. Intensity and coherence of motifs in weighted complex networks. Phys. Rev. E 2005, 71, 065103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccaletti, S.; Latora, V.; Moreno, Y.; Chavez, M.; Hwanga, D.U. Complex networks: Structure and dynamics. Phys. Rep. 2006, 424, 175–308. [Google Scholar] [CrossRef]
- Stam, C.J.; van Straaten, E.C. The organization of physiological brain networks. Clin. Neurophysiol. 2012, 123, 1067–1087. [Google Scholar] [CrossRef] [PubMed]
- Muldoon, S.F.; Bridgeford, E.W.; Bassett, D.S. Small-world propensity and weighted brain networks. Sci. Rep. 2016, 6, 22057. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Nelson, B.G.; Mueller, B.A.; Camchong, J.; Lim, K.O. Altered resting state complexity in schizophrenia. NeuroImage 2012, 59, 2196–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG-and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Mason, D.M.; Newton, M.A. A Rank Statistics Approach to the Consistency of a General Bootstrap. Ann. Statist. 1992, 20, 1611–1624. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annett, M. A classification of hand preference by association analysis. Br. J. Psychol. 1970, 61, 303–321. [Google Scholar] [CrossRef]
- Dushanova, J.; Lalova, Y.; Kalonkina, A.; Tsokov, S. Speech-Brain Frequency Entrainment of Dyslexia with and without Phonological Deficits. Brain Sci. 2020, 10, 920. [Google Scholar] [CrossRef]
- Koessler, L.; Maillard, L.; Benhadid, A.; Vignal, J.P.; Felblinger, J.; Vespignani, H.; Braun, M. Automated cortical projection of EEG sensors: Anatomical correlation via the international 10-10 system. NeuroImage 2009, 46, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hülsmann, E.R.M. Traveling cortical netwaves compose a mindstream. bioRxiv 2020, 1–25, Preprint. [Google Scholar]
- Giacometti, P.; Perdue, K.L.; Diamond, S.G. Algorithm to find high density EEG scalp coordinates and analysis of their correspondence to structural and functional regions of the brain. J. Neurosci. Methods 2014, 229, 84–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, C.E.; Lakatos, P. The gamma oscillation: Master or slave? Brain Topogr. 2009, 22, 24–26. [Google Scholar] [CrossRef]
- Lobier, M.; Dubois, M.; Valdois, S. The role of cisual processing speed in reading speed development. PLoS ONE 2013, 8, e58097. [Google Scholar] [CrossRef] [Green Version]
- Bosse, M.L.; Tainturier, M.J.; Vadois, S. Developmental dyslexia: The visual attention span deficit hypothesis. Cognition 2007, 104, 198–230. [Google Scholar] [CrossRef] [Green Version]
- Rayner, K. Eye movements in reading and information processing: 20 years of research. Psychol. Bull. 1998, 124, 372–422. [Google Scholar] [CrossRef] [PubMed]
- Busch, N.A.; Van Rullen, R. Spontaneous EEG scillations reveal periodic sampling of visual attention. Proc. Natl. Acad. Sci. USA 2010, 107, 16048–16053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathewson, K.E.; Prudhomme, C.; Fabiani, M.; Back, D.M.; Lleras, A.; Gratton, G. Making waves in the stream of consciousness: Entraining oscillations in EEG alpha and fluctuations in visual awareness with rhythmic visual stimulation. J. Cogn. Neurosci. 2012, 24, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.; Martinkauppi, S.; Rama, P.; Salli, E.; Korvenoja, A.; Aronen, H.J. Distribution of cortical activation during visuospatial n-back tasks as revealed by functional magnetic resonance imaging. Cereb Cortex 1998, 8, 743–752. [Google Scholar] [CrossRef]
- Spitzer, H.; Desimone, R.; Moran, J. Increased attention enhances both behavioral and neuronal performance. Science 1988, 240, 338–340. [Google Scholar] [CrossRef] [Green Version]
- Schluppeck, D.; Curtis, C.E.; Glimcher, P.W.; Heeger, D.J. Sustained activity in topographic areas of human posterior parietal cortex during memory-guided saccades. J. Neurosci. 2006, 26, 5098–5108. [Google Scholar] [CrossRef] [Green Version]
- Koenderink, J.J.; Physics, P. Optic flow. Vis. Res. 2001, 26, 161–179. [Google Scholar] [CrossRef]
- Pandya, D.N.; Seltzer, B. Intrinsic connections and architectonics of posterior parietal cortex in the rhesus monkey. J. Comp. Neurol. 1986, 204, 196–210. [Google Scholar] [CrossRef]
- Luppino, G.; Hamed, S.B.; Gamberini, M.; Matelli, M.; Galletti, C. Occipital (V6) and parietal (V6A) areas in the anterior wall of the parieto-occipital sulcus of the macaque: A cytoarchitectonic study. Eur. J. Neurosci. 2005, 21, 3056–3076. [Google Scholar] [CrossRef]
- Cardin, V.; Hemsworth, L.; Smith, A.T. Adaptation to heading direction dissociates the roles of human MST and V6 in the processing of optic flow. J. Neurophysiol. 2012, 108, 794–801. [Google Scholar] [CrossRef]
- Quinlan, D.J.; Culham, J.C. fMRI reveals a preference for near viewing in the human parieto-occipital cortex. NeuroImage 2007, 36, 167–187. [Google Scholar] [CrossRef]
- Frank, S.M.; Greenlee, M.W. The parieto-insular vestibular cortex in humans: More than a single area? J. Neurophysiol. 2018, 120, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oostende, S.; Sunaert, S.; Van Hecke, P.; Marchal, G.; Orban, G.A. The kinetic occipital (KO) region in man: An fMRI study. Cereb. Cortex 1997, 7, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Sereno, M.I.; Dale, A.M.; Reppas, J.B.; Kwong, K.K.; Belliveau, J.W.; Brady, T.J.; Rosen, B.R.; Tootell, R.B.H. Borders of multiple visual areas in humans revealed by functional magnetic resonance imaging. Science 1995, 268, 889–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnoldussen, D.M.; Goossens, J.; van den Berg, A.V. Adjacent visual representations of self-motion in different reference frames. Proc. Natl. Acad. Sci. USA 2011, 108, 11668–11673. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, R.F.; Becker, H.G.; Haarmeier, T. Processing of coherent visual motion in topographically organized visual areas in human cerebral cortex. Brain Topogr. 2013, 26, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Orban, G.A.; Sunaert, S.; Todd, J.T.; Van Hecke, P.; Marchal, G. Human cortical regions involved in extracting depth from motion. Neuron 1999, 24, 929–940. [Google Scholar] [CrossRef] [Green Version]
- Sereno, M.I.; Pitzalis, S.; Martinez, A. Mapping of contralateral space in retinotopic coordinates by a parietal cortical area in humans. Science 2001, 294, 1350–1354. [Google Scholar] [CrossRef] [Green Version]
- Frank, S.M.; Wirth, A.M.; Greenlee, M.W. Visual-vestibular processing in the human Sylvian fissure. J. Neurophysiol. 2016, 116, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Gitelman, D.R.; Nobre, A.C.; Parrish, T.B.; LaBar, K.S.; Kim, Y.-H.; Meyer, J.R.; Mesulam, M.-M. A large-scale distributed network for covert spatial attention: Further anatomical delineation based on stringent behavioral and cognitive controls. Brain 1999, 122, 1093–1106. [Google Scholar] [CrossRef] [Green Version]
- Greenlee, M.W.; Frank, S.M.; Kaliuzhna, M.; Blanke, O.; Bremmer, F.; Churan, J.; Cuturi, L.F.; MacNeilage, P.R.; Smith, A.T. Multisensory integration in self motion perception. Multisens. Res. 2016, 29, 525–556. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.J.; Zeki, S. The neurology of saccades and covert shifts in spatial attention. Brain 2000, 123, 2273–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astafiev, S.V.; Shulman, G.L.; Stanley, C.M.; Snyder, A.Z.; Van Essen, D.C.; Corbetta, M. Functional organization of human intraparietal and frontal cortex for attending, looking, and pointing. J. Neurosci. 2003, 23, 4689–4699. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.S.; Chen, C.F.; Sereno, M.I. Neural substrates underlying the passive observation and active control of translational egomotion. J. Neurosci. 2015, 35, 4258–4267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colby, C.L.; Duhamel, J.R.; Goldberg, M.E. Ventral intraparietal area of the macaque: Anatomic location and visual response properties. J. Neurophysiol. 1993, 69, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Bisley, J.W.; Goldberg, M.E. Neuronal activity in the lateral intraparietal area and spatial attention. Science 2003, 299, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.T.; Wilkie, R.M.; Wann, J.P. Neural systems in the visual control of steering. J. Neurosci. 2007, 27, 8002–8010. [Google Scholar] [CrossRef] [PubMed]
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends Cogn. Sci. 2005, 9, 474–480. [Google Scholar] [CrossRef]
- Saalmann, Y.B.; Pigarev, I.N.; Vidyasagar, T.R. Neural mechanisms of visual attention: How top-down feedback highlights relevant locations. Science 2007, 316, 1612–1615. [Google Scholar] [CrossRef]
- Buschman, T.J.; Miller, E.K. Top-down and bottom-up control of attention in the prefrontal and posterior parietal cortices. Science 2007, 315, 1860–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Hutcheon, B.; Yarom, Y. Resonance, oscillation and the intrinsic frequency preference of neurons. Trends Neurosci. 2000, 23, 216–222. [Google Scholar] [CrossRef]
- Economo, M.N.; White, J.A. Membrane properties and the balance between excitation and inhibition control gamma-frequency oscillations arising from feedback inhibition. PLoS Comput. Biol. 2012, 8, e1002354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhorn, R.; Bauer, R.; Jordan, W.; Brosch, M.; Kruse, W.; Munk, M.; Reitboeck, H.J. Coherent oscillations: A mechanism feature linking in the visual cortex. Multiple electrode and correlation analyses in the cat. Biol. Cybern. 1988, 60, 121–130. [Google Scholar] [CrossRef]
- Pierrot-Deseilligny, C.; Milea, D.; Müri, R.M. Eye movement control by the cerebral cortex. Curr. Opin. Neurol. 2004, 17, 17–25. [Google Scholar] [CrossRef]
- Grosbras, M.H.; Lobel, E.; Van de Moortele, P.F.; LeBihan, D.; Berthoz, A. An anatomical landmark for the supplementary eye fields in human revealed with functional magnetic resonance imaging. Cereb. Cortex 1999, 9, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Rosano, C.; Krisky, C.M.; Welling, J.S.; Eddy, W.F.; Lun, A.B.; Thulborn, K.R.; Sweeney, J.A. Pursuit and saccadic eye movement subregions in human frontal eye field: A high-resolution fMRI investigation. Cereb. Cortex 2002, 12, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Müri, R.M.; Iba-Zizen, M.T.; Derosier, C.; Cabanis, E.A.; Pierrot-Deseillign, C. Location of the human posterior eye field with functional magnetic resonance imaging. J. Neurol. Neurosurg. Psychiatry 1996, 60, 445–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, M.; Bucher, S.F.; Seelos, K.C.; Brandt, T. Horizontal or vertical optokinetic stimulation activates visual motion-sensitive, ocular motor and vestibular cortex areas with right hemispheric dominance. An fMRI study. Brain 1998, 121, 1479–1495. [Google Scholar] [CrossRef] [Green Version]
- Dieterich, M.; Bense, S.; Stephan, T.; Yousry, T.A.; Brandt, T. FMRI signal increases and decreases in cortical areas during small-field optokinetic stimulation and central fixation. Exp. Brain Res. 2003, 148, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Pierrot-Deseilligny, C.; Rivaud, S.; Gaymard, B.; Agid, Y. Cortical control of reflexive visually guided saccades. Brain 1991, 114, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Rivaud, S.; Müri, R.M.; Gaymard, B.; Vermersch, A.I.; Pierrot-Deseilligny, C. Eye movement disorders after frontal eye field lesions in humans. Exp. Brain Res. 1994, 102, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, B.; François, C.; Ploner, C.J.; Condy, C.; Rivaud-Péchoux, S. A direct prefrontotectal tract against distractibility in the human brain. Ann. Neurol. 2003, 53, 542–545. [Google Scholar] [CrossRef]
- Brotchie, P.R.; Lee, M.B.; Chen, D.Y.; Lourensz, M.; Jackson, G.; Bradley, W.G., Jr. Head position modulates activity in the human parietal eye fields. NeuroImage 2003, 18, 178–184. [Google Scholar] [CrossRef]
- Kawashima, R.; Naitoh, E.; Matsumara, M.; Ono, S.; Satoh, K.; Gotoh, R.; Koyama, M.; Inoue, K.; Yoshioka, S.; Fukuda, H. Topographic representation in human intraparietal sulcus of reaching and saccade. NeuroReport 1996, 7, 1253–1256. [Google Scholar] [CrossRef]
- Simon, O.; Mangin, J.F.; Cohen, L.; Bihan, D.L.; Dehaene, S. Topographical layout of hand, eye, calculation and language-related areas in the human parietal lobe. Neuron 2002, 33, 475–487. [Google Scholar] [CrossRef] [Green Version]
- De Souza, J.F.X.; Dukelow, S.P.; Gati, J.S.; Menon, R.S.; Andersen, R.A.; Vilis, T. Eye position signal modulates a human parietal pointing region during memory-guided movements. J. Neurosci. 2000, 20, 5835–5840. [Google Scholar] [CrossRef] [Green Version]
- Macaluso, E.; Driver, J.; Frith, C.D. Multimodal spatial representations engaged in human parietal cortex during both saccadic and manual spatial orienting. Curr. Biol. 2003, 13, 990–999. [Google Scholar] [CrossRef] [Green Version]
- Yantis, S.; Schwarzbach, J.; Serences, J.T.; Carlson, R.L.; Steinmetz, M.A.; Pekar, J.J.; Courtney, S.M. Transient neural activity in human parietal cortex during spatial attention shifts. Nat. Neurosci. 2002, 5, 995–1002. [Google Scholar] [CrossRef]
- Mort, D.J.; Perry, R.J.; Mannan, S.K.; Kmannan, S.; Hodgson, T.L.; Anderson, E.; Quest, R.; McRobbie, D.; McBride, A.; Husain, M.; et al. Differential cortical activation during voluntary and reflexive saccades in man. NeuroImage 2003, 18, 231–246. [Google Scholar] [CrossRef]
- Raffi, M.; Carrozzini, C.; Maioli, M.G.; Squatrito, S. Multimodal representation of optic flow in area PEc of macaque monkey. Neuroscience 2010, 171, 1241–1255. [Google Scholar] [CrossRef]
- Pisella, L.; Berberovic, N.; Mattingley, J.B. Impaired working memory for location but not for colour or shape in visual neglect: A comparison of parietal and non-parietal lesions. Cortex 2004, 40, 379–390. [Google Scholar] [CrossRef]
- Vallar, G.; Perani, D. The anatomy of unilateral neglect after right-hemisphere stroke lesions. A clinical/CT-scan correlation study in man. Neuropsychologia 1986, 24, 609–622. [Google Scholar] [CrossRef]
- Saleh, R.; Alias, N.A. Learner Needs Analysis for Mobile Learning Comic Application among Dyslexic Children. Int. J. Educ. Inf. Technol. 2012, 6, 185–192. [Google Scholar]
- Rahman, F.A.; Mokhtar, F.; Alias, N.A.; Saleh, R. Multimedia Elements as Instructions for Dyslexic Children. Int. J. Educ. Inf. Technol. 2012, 6, 193–200. [Google Scholar]
- Pistoia, M.; Pinnelli, S.; Borrelli, G. Use of a robotic platform in dyslexia-affected pupils: The ROBIN project experience. Int. J. Educ. Inf. Technol. 2015, 9, 46–49. [Google Scholar]


| Success/RT | Con | Pre-D | Post-D | Con/Pre-D | Con/Post-D | Pre-/Post-D | |||
|---|---|---|---|---|---|---|---|---|---|
| (%), (s) | mean ± s.e. | mean ± s.e. | mean ± s.e. | p | χ2 | p | χ2 | p | χ2 |
| 1. slow | 72.5 ± 3.1 | 60.8 ± 3.9 | 75.3 ± 5.6 | 0.02 | 5.83 | 0.94 | 0.004 | 0.01 | 6.07 |
| RT | 1.14 ± 0.023 | 1.56 ± 0.05 | 1.64 ± 0.05 | <0.0001 | 41.38 | <0.001 | 34.4 | 0.441 | 0.59 |
| 2. fast | 76.3 ± 2.9 | 61 ± 3.84 | 78.3 ± 2.8 | 0.0004 | 16.32 | 0.74 | 0.05 | 0.0001 | 19.02 |
| RT | 1.13 ± 0.024 | 1.44 ± 0.03 | 1.66 ± 0.08 | 0.004 | 7.90 | <0.001 | 19.8 | <0.0001 | 21.44 |
| Frequency/Metric | Con | Pre-D | Post-D | Con/Pre-D | Con/Post-D | Pre-/Post-D | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| p | χ2 | p | χ2 | p | χ2 | |||||
| δ | ϕ | 0.583 ± 0.016 | 0.611 ± 0.014 | 0.590 ± 0.018 | 0.191 | 1.70 | 0.529 | 0.39 | 0.440 | 0.59 |
| ΔC | 0.497 ± 0.031 | 0.413 ± 0.025 | 0.473 ± 0.034 | 0.039 | 4.22 | 0.459 | 0.54 | 0.292 | 1.11 | |
| ΔL | 0.234 ± 0.020 | 0.273 ± 0.019 | 0.264 ± 0.017 | 0.257 | 1.27 | 0.096 | 2.76 | 0.523 | 0.40 | |
| θ | ϕ | 0.506 ± 0.011 | 0.540 ± 0.010 | 0.514 ± 0.013 | 0.035 | 4.40 | 0.858 | 0.03 | 0.081 | 3.03 |
| ΔC | 0.667 ± 0.019 | 0.594 ± 0.017 | 0.660 ± 0.021 | 0.008 | 7.03 | 0.793 | 0.06 | 0.020 | 5.40 | |
| ΔL | 0.141 ± 0.008 | 0.185 ± 0.011 | 0.145 ± 0.008 | 0.003 | 8.48 | 0.464 | 0.53 | 0.048 | 3.89 | |
| α | ϕ | 0.478 ± 0.009 | 0.512 ± 0.008 | 0.510 ± 0.011 | 0.006 | 7.34 | 0.039 | 4.21 | 0.751 | 0.10 |
| ΔC | 0.724 ± 0.014 | 0.653 ± 0.014 | 0.670 ± 0.019 | 0.0009 | 11.00 | 0.043 | 4.05 | 0.352 | 0.86 | |
| ΔL | 0.106 ± 0.005 | 0.136 ± 0.010 | 0.115 ± 0.006 | 0.276 | 1.18 | 0.302 | 1.06 | 0.961 | 0.002 | |
| β1 | ϕ | 0.460 ± 0.008 | 0.497 ± 0.008 | 0.435 ± 0.008 | 0.010 | 6.57 | 0.020 | 5.33 | 6.65 × 10−6 | 20.29 |
| ΔC | 0.755 ± 0.012 | 0.690 ± 0.014 | 0.789 ± 0.014 | 0.007 | 7.18 | 0.017 | 5.68 | 3.18 × 10−6 | 21.71 | |
| ΔL | 0.077 ± 0.004 | 0.102 ± 0.006 | 0.064 ± 0.006 | 0.024 | 5.09 | 0.006 | 7.47 | 4.96 × 10−6 | 20.85 | |
| β2 | ϕ | 0.447 ± 0.007 | 0.494 ± 0.007 | 0.434 ± 0.007 | 2.02 × 10−5 | 18.17 | 0.310 | 1.02 | 4.96 × 10−7 | 25.28 |
| ΔC | 0.775 ± 0.011 | 0.694 ± 0.012 | 0.794 ± 0.011 | 4.06 × 10−5 | 21.23 | 0.291 | 1.11 | 7.95 × 10−8 | 28.81 | |
| ΔL | 0.067 ± 0.004 | 0.100 ± 0.007 | 0.059 ± 0.004 | 0.008 | 6.82 | 0.278 | 1.17 | 0.0002 | 13.55 | |
| γ1 | ϕ | 0.479 ± 0.008 | 0.540 ± 0.009 | 0.481 ± 0.009 | 1.13 × 10−5 | 19.27 | 0.927 | 0.008 | 5.45 × 10−5 | 16.28 |
| ΔC | 0.721 ± 0.013 | 0.618 ± 0.014 | 0.721 ± 0.015 | 4.44 × 10−6 | 21.06 | 0.963 | 0.002 | 1. 2 × 10−5 | 19.16 | |
| ΔL | 0.104 ± 0.006 | 0.136 ± 0.007 | 0.093 ± 0.005 | 0.006 | 7.45 | 0.377 | 0.77 | 0.0004 | 12.34 | |
| γ2 | ϕ | 0.484 ± 0.013 | 0.475 ± 0.011 | 0.484 ± 0.012 | 0.262 | 1.25 | 0.951 | 0.003 | 0.242 | 1.36 |
| ΔC | 0.432 ± 0.024 | 0.488 ± 0.022 | 0.476 ± 0.027 | 0.066 | 3.36 | 0.178 | 1.81 | 0.643 | 0.21 | |
| ΔL | 0.442 ± 0.026 | 0.385 ± 0.021 | 0.406 ± 0.024 | 0.061 | 3.50 | 0.660 | 0.192 | 0.173 | 1.84 | |
| Frequency/Metric | Con | Pre-D | Post-D | Con/Pre-D | Con/Post-D | Pre-/Post-D | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| p | χ2 | p | χ2 | p | χ2 | |||||
| δ | ϕ | 0.625 ± 0.020 | 0.653 ± 0.015 | 0.647 ± 0.019 | 0.631 | 0.23 | 0.656 | 0.19 | 0.293 | 1.10 |
| ΔC | 0.389 ± 0.045 | 0.415 ± 0.026 | 0.383 ± 0.034 | 0.576 | 0.31 | 0.958 | 0.002 | 0.462 | 0.54 | |
| ΔL | 0.273 ± 0.021 | 0.267 ± 0.019 | 0.258 ± 0.020 | 0.294 | 1.09 | 0.381 | 0.76 | 0.652 | 0.20 | |
| θ | ϕ | 0.517 ± 0.014 | 0.538 ± 0.011 | 0.522 ± 0.013 | 0.181 | 1.78 | 0.758 | 0.09 | 0.294 | 1.09 |
| ΔC | 0.653 ± 0.023 | 0.614 ± 0.018 | 0.646 ± 0.021 | 0.163 | 1.93 | 0.778 | 0.07 | 0.251 | 1.31 | |
| ΔL | 0.137 ± 0.011 | 0.161 ± 0.009 | 0.148 ± 0.009 | 0.049 | 3.84 | 0.334 | 0.93 | 0.440 | 0.59 | |
| α | ϕ | 0.509 ± 0.009 | 0.534 ± 0.010 | 0.462 ± 0.010 | 0.145 | 2.11 | 0.0006 | 11.57 | 2.24 × 10−6 | 22.37 |
| ΔC | 0.670 ± 0.016 | 0.635 ± 0.015 | 0.746 ± 0.015 | 0.153 | 2.03 | 0.0007 | 11.35 | 1.90 × 10−6 | 22.68 | |
| ΔL | 0.126 ± 0.008 | 0.130 ± 0.006 | 0.103 ± 0.006 | 0.421 | 0.64 | 0.060 | 3.51 | 0.002 | 9.34 | |
| β1 | ϕ | 0.487 ± 0.010 | 0.492 ± 0.009 | 0.446 ± 0.007 | 0.912 | 0.01 | 0.005 | 7.75 | 0.003 | 8.43 |
| ΔC | 0.712 ± 0.016 | 0.701 ± 0.014 | 0.773 ± 0.012 | 0.897 | 0.02 | 0.004 | 8.09 | 0.002 | 9.42 | |
| ΔL | 0.089 ± 0.006 | 0.091 ± 0.006 | 0.075 ± 0.005 | 0.987 | 0.0002 | 0.294 | 1.09 | 0.223 | 1.48 | |
| β2 | ϕ | 0.455 ± 0.009 | 0.498 ± 0.009 | 0.438 ± 0.009 | 0.003 | 8.65 | 0.110 | 2.55 | 9.42 × 10−7 | 24.04 |
| ΔC | 0.762 ± 0.014 | 0.689 ± 0.015 | 0.791 ± 0.013 | 0.002 | 8.94 | 0.09 | 2.77 | 5.74 × 10−7 | 24.99 | |
| ΔL | 0.071 ± 0.005 | 0.089 ± 0.007 | 0.056 ± 0.003 | 0.155 | 2.02 | 0.152 | 2.04 | 0.003 | 8.46 | |
| γ1 | ϕ | 0.512 ± 0.009 | 0.532 ± 0.010 | 0.491 ± 0.009 | 0.373 | 0.79 | 0.099 | 2.71 | 0.011 | 6.43 |
| ΔC | 0.672 ± 0.015 | 0.623 ± 0.017 | 0.704 ± 0.015 | 0.242 | 1.36 | 0.078 | 3.08 | 0.003 | 8.46 | |
| ΔL | 0.117 ± 0.006 | 0.133 ± 0.010 | 0.103 ± 0.006 | 0.801 | 0.06 | 0.032 | 4.56 | 0.087 | 2.92 | |
| γ2 | ϕ | 0.492 ± 0.012 | 0.484 ± 0.012 | 0.502 ± 0.012 | 0.525 | 0.40 | 0.673 | 0.177 | 0.253 | 1.30 |
| ΔC | 0.399 ± 0.024 | 0.462 ± 0.026 | 0.479 ± 0.025 | 0.105 | 2.62 | 0.021 | 5.25 | 0.778 | 0.07 | |
| ΔL | 0.453 ± 0.026 | 0.392 ± 0.023 | 0.374 ± 0.023 | 0.131 | 2.26 | 0.075 | 3.16 | 0.850 | 0.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taskov, T.; Dushanova, J. Functional Connectivity in Developmental Dyslexia during Speed Discrimination. Symmetry 2021, 13, 749. https://doi.org/10.3390/sym13050749
Taskov T, Dushanova J. Functional Connectivity in Developmental Dyslexia during Speed Discrimination. Symmetry. 2021; 13(5):749. https://doi.org/10.3390/sym13050749
Chicago/Turabian StyleTaskov, Tihomir, and Juliana Dushanova. 2021. "Functional Connectivity in Developmental Dyslexia during Speed Discrimination" Symmetry 13, no. 5: 749. https://doi.org/10.3390/sym13050749
APA StyleTaskov, T., & Dushanova, J. (2021). Functional Connectivity in Developmental Dyslexia during Speed Discrimination. Symmetry, 13(5), 749. https://doi.org/10.3390/sym13050749