
Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis



Abstract
:1. Introduction
2. Dynamics of Total Nitrogen Transformation
2.1. Equilibrium Points and Local Stability Analysis
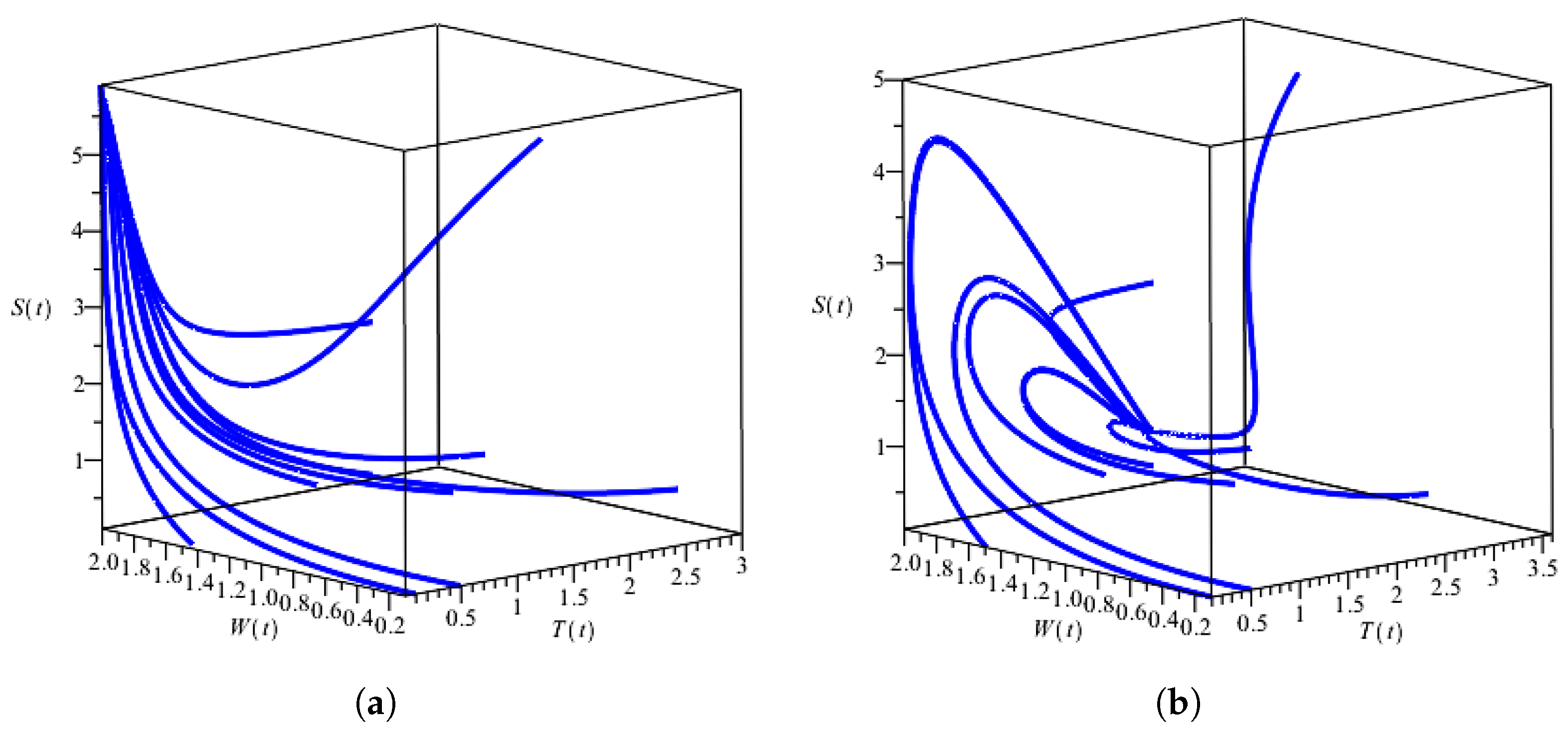
2.2. Numerical Solution
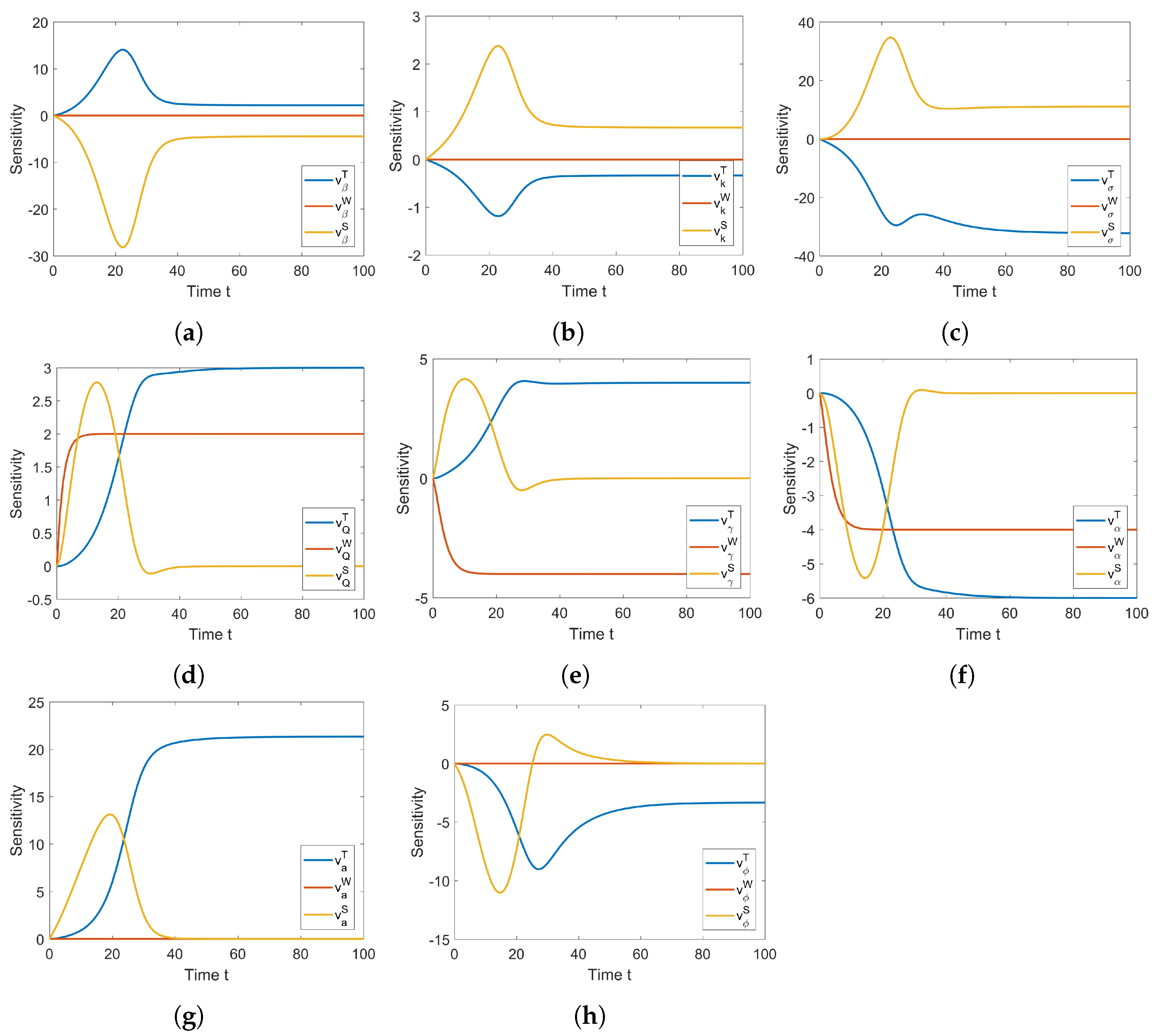
2.3. Sensitivity Analysis
3. Discrete Form Model
3.1. The Local Stability of Non-Zero Equilibrium
3.2. Numerical Simulations
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B
References
- Choi, H.; Geronimo, F.K.; Jeon, M.; Kim, L.H. Evaluation of bacterial community in constructed wetlands treating different sources of wastewater. Ecol. Eng. 2022, 182, 106703. [Google Scholar] [CrossRef]
- Bunwong, K.; Sae-jie, W.; Lenbury, Y. Modelling nitrogen dynamics of a constructed wetland: Nutrient removal process with variable yield. Nonlinear Anal. Theory Methods Appl. 2009, 71, e1538–e1546. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; Ngo, H.H.; Guo, W.; Hu, Z.; Liang, S.; Fan, J.; Liu, H. A review on the sustainability of constructed wetlands for wastewater treatment: Design and operation. Bioresour. Technol. 2015, 175, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Senzia, M.A.; Mashauri, D.A.; Mayo, A.W. Suitability of constructed wetlands and waste stabilisation ponds in wastewater treatment: Nitrogen transformation and removal. Phys. Chem. Earth Parts A/B/C 2003, 28, 1117–1124. [Google Scholar] [CrossRef]
- Han, Z.; Dong, J.; Shen, Z.; Mou, R.; Zhou, Y.; Chen, X.; Fu, X.; Yang, C. Nitrogen removal of anaerobically digested swine wastewater by pilot-scale tidal flow constructed wetland based on in-situ biological regeneration of zeolite. Chemosphere 2019, 217, 364–373. [Google Scholar] [CrossRef]
- Strigul, N.S.; Kravchenko, L.V. Mathematical modeling of PGPR inoculation into the rhizosphere. Environ. Model. Softw. 2006, 21, 1158–1171. [Google Scholar] [CrossRef]
- Jia, L.; Gou, E.; Liu, H.; Lu, S.; Wu, S.; Wu, H. Exploring utilization of recycled agricultural biomass in constructed wetlands: Characterization of the driving force for high-rate nitrogen removal. Environ. Sci. Technol. 2019, 53, 1258–1268. [Google Scholar] [CrossRef]
- Choi, H.; Geronimo, F.K.F.; Jeon, M.; Kim, L.H. Investigation of the Factors Affecting the Treatment Performance of a Stormwater Horizontal Subsurface Flow Constructed Wetland Treating Road and Parking lot Runoff. Water 2021, 13, 1242. [Google Scholar] [CrossRef]
- Rousseau, D.P.L.; Louage, F.; Wang, Q.; Zhang, R. Constructed Wetlands for Urban Wastewater Treatment: An Overview. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 272–284. [Google Scholar]
- Chen, Y.; Zhang, J.; Guo, Z.; Li, M.; Wu, H. Optimizing agricultural biomass application to enhance nitrogen removal in vertical flow constructed wetlands for treating low-carbon wastewater. Environ. Res. 2022, 209, 112867. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Dong, L.; Han, C.; Li, M.; Wu, H. Effects of biochar dosage on treatment performance, enzyme activity and microbial community in aerated constructed wetlands for treating low C/N domestic sewage. Environ. Technol. Innovat. 2021, 24, 101919. [Google Scholar] [CrossRef]
- Jia, L.; Li, C.; Zhang, Y.; Chen, Y.; Li, M.; Wu, S.; Wu, H. Microbial community responses to agricultural biomass addition in aerated constructed wetlands treating low carbon wastewater. J. Environ. Manag. 2020, 270, 110912. [Google Scholar] [CrossRef]
- Wu, Y.; Chung, A.; Tam, N.F.Y.; Pi, N.; Wong, M.H. Constructed mangrove wetland as secondary treatment system for municipal wastewater. Ecol. Eng. 2008, 34, 137–146. [Google Scholar] [CrossRef]
- Leung, J.Y.S.; Cai, Q.; Tam, N.F.Y. Comparing subsurface flow constructed wetlands with mangrove plants and freshwater wetland plants for removing nutrients and toxic pollutants. Ecol. Eng. 2016, 95, 129–137. [Google Scholar] [CrossRef]
- Lobry, J.R.; Flandrois, J.P.; Carret, G.; Pave, A. Monod’s bacterial growth model revisited. Bull. Math. Biol. 1992, 54, 117–122. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Lee, S.C.; Juang, H.T.; Hur, M.T.; Hwang, Y.H. Nitrogen nutritional status and fate of applied N in Mangrove soils. Bot. Bull. Acad. Sin. 1996, 37, 191–196. [Google Scholar]
- Pisman, T.I.; Pechurkin, N.S.; Mariasova, T.S.; Somova, L.A.; Sarangova, A.B. A mathematical model of “plants-microorganisms” interaction on complete mineral medium and under nitrogen limitation. Adv. Space Res. 1999, 24, 383–387. [Google Scholar] [CrossRef]
- Walker, R.L.; Burns, I.G.; Moorby, J. Responses of plant growth rate to nitrogen supply: A comparison of relative addition and N interruption treatments. J. Exp. Bot. 2001, 52, 309–317. [Google Scholar] [CrossRef]
- Pilyugin, S.S.; Waltman, P. Multiple limit cycles in the chemostat with variable yield. Math. Biosci. 2003, 182, 151–166. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, X. Multiple limit cycles in a continuous culture vessel with variable yield. Nonlinear Anal. 2006, 64, 887–894. [Google Scholar] [CrossRef]
- Zhao, Z.; Pang, L.; Zhao, Z.; Luo, C. Impulsive State Feedback Control of the Rhizosphere Microbial Degradation in the Wetland Plant. Discret. Dyn. Nat. Soc. 2015, 2015, 612354. [Google Scholar] [CrossRef]
- Zhao, Z.; Song, Y.; Pang, L. Mathematical modeling of rhizosphere microbial degradation with impulsive diffusion on nutrient. Adv. Differ. Equ. 2016, 2016, 24. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Li, Q.; Chen, L. Effect of rhizosphere dispersal and impulsive input on the growth of wetland plant. Math. Comput. Simul. 2018, 152, 69–80. [Google Scholar] [CrossRef]
- Zhao, Z.; Yin, Q.; Li, Q.; Wu, X. Mathematical model for diffusion of the rhizosphere microbial degradation with impulsive feedback control. J. Biol. Dyn. 2020, 14, 566–577. [Google Scholar] [CrossRef]
- Suandi, D.; Ningrum, I.P.; Alifah, A.N.; Izzah, N.; Reza, M.P.; Muwahidah, I.K. Mathematical Modeling and Sensitivity Analysis of the Existence of Male Calico Cats Population Based on Cross Breeding of All Coat Colour Types. Commun. Biomath. Sci. 2019, 2, 96–104. [Google Scholar] [CrossRef]
- Mapfumo, K.Z.; Pagan’a, J.C.; Juma, V.O.; Kavallaris, N.I.; Madzvamuse, A. A Model for the Proliferation–Quiescence Transition in Human Cells. Mathematics 2022, 10, 2426. [Google Scholar] [CrossRef]
- Rentzeperis, F.; Wallace, D. Local and global sensitivity analysis of spheroid and xenograft models of the acid-mediated development of tumor malignancy. Appl. Math. Model. 2022, 109, 629–650. [Google Scholar] [CrossRef]
- Alsahafi, S.; Woodcock, S. Exploring HIV Dynamics and an Optimal Control Strategy. Mathematics 2022, 10, 749. [Google Scholar] [CrossRef]
- Reinharz, V.; Churkin, A.; Dahari, H.; Barash, D. Advances in Parameter Estimation and Learning from Data for Mathematical Models of Hepatitis C Viral Kinetics. Mathematics 2022, 10, 2136. [Google Scholar] [CrossRef]
- Ansori, M.F.; Sumarti, N.; Sidarto, K.A.; Guandi, I. Analyzing a macroprudential instrument during the covid-19 pandemic using border collision bifurcation. Rev. Electron. Comun. Y Trab. ASEPUMA Rect. 2021, 22, 113–125. [Google Scholar] [CrossRef]
- Du, X.; Han, X.; Lei, C. Behavior Analysis of a Class of Discrete-Time Dynamical System with Capture Rate. Mathematics 2022, 10, 2410. [Google Scholar] [CrossRef]
- Saeed, T.; Djeddi, K.; Guirao, J.L.G.; Alsulami, H.H.; Alhodaly, M.S. A Discrete Dynamics Approach to a Tumor System. Mathematics 2022, 10, 1774. [Google Scholar] [CrossRef]
- Liu, B.; Wu, R. Bifurcation and Patterns Analysis for a Spatiotemporal Discrete Gierer-Meinhardt System. Mathematics 2022, 10, 243. [Google Scholar] [CrossRef]
- He, Z.Y.; Abbes, A.; Jahanshahi, H.; Alotaibi, N.D.; Wang, Y. Fractional-Order Discrete-Time SIR Epidemic Model with Vaccination: Chaos and Complexity. Mathematics 2022, 10, 165. [Google Scholar] [CrossRef]
- Gandolfo, G. Economic Dynamics: Methods and Models, 2nd ed.; Elsevier Science Publisher BV: Amsterdam, The Netherlands, 1985. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Notation | Description | Value | Unit |
|---|---|---|---|
| Maximum possible value of the plant growth rate at infinite total nitrogen concentration in the soil solution | 0.25 | ||
| k | Semi-saturation | 1 | |
| Level of garbage | 0.1 | ||
| Q | Input level of total nitrogen concentration | 1 | |
| Exchange rate of total nitrogen between wastewater and soil solution | 0.3 | ||
| Rate of total nitrogen loss in wastewater through runoff or evaporation | 0.2 | ||
| a | Conversion of nutrients consumed from biomass produced | 0.5 | Dimensionless |
| Rate of total nitrogen loss in the soil solution by leaching or denitrification | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunarsih; Ansori, M.F.; Khabibah, S.; Sasongko, D.P. Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry 2022, 14, 1924. https://doi.org/10.3390/sym14091924
Sunarsih, Ansori MF, Khabibah S, Sasongko DP. Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry. 2022; 14(9):1924. https://doi.org/10.3390/sym14091924
Chicago/Turabian StyleSunarsih, Moch. Fandi Ansori, Siti Khabibah, and Dwi Purwantoro Sasongko. 2022. "Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis" Symmetry 14, no. 9: 1924. https://doi.org/10.3390/sym14091924
APA StyleSunarsih, Ansori, M. F., Khabibah, S., & Sasongko, D. P. (2022). Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry, 14(9), 1924. https://doi.org/10.3390/sym14091924