

A Method for Calculating the Sign and Degree of Chirality of Supercoiled Protein Structures


{kind=link}
{kind=link}
{kind=link}
{kind=link}
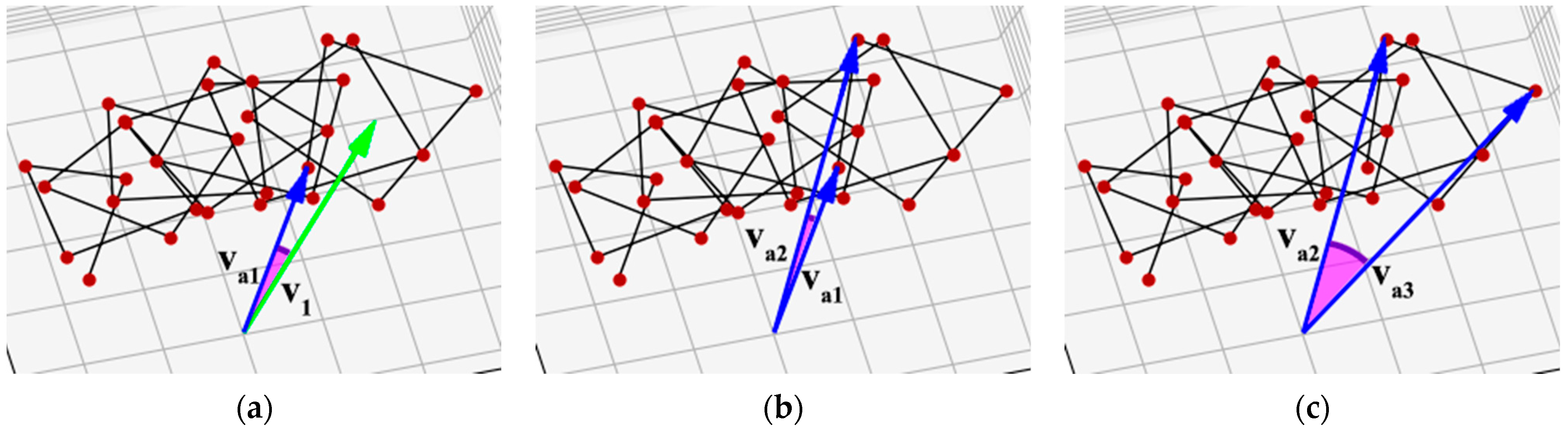{kind=link}
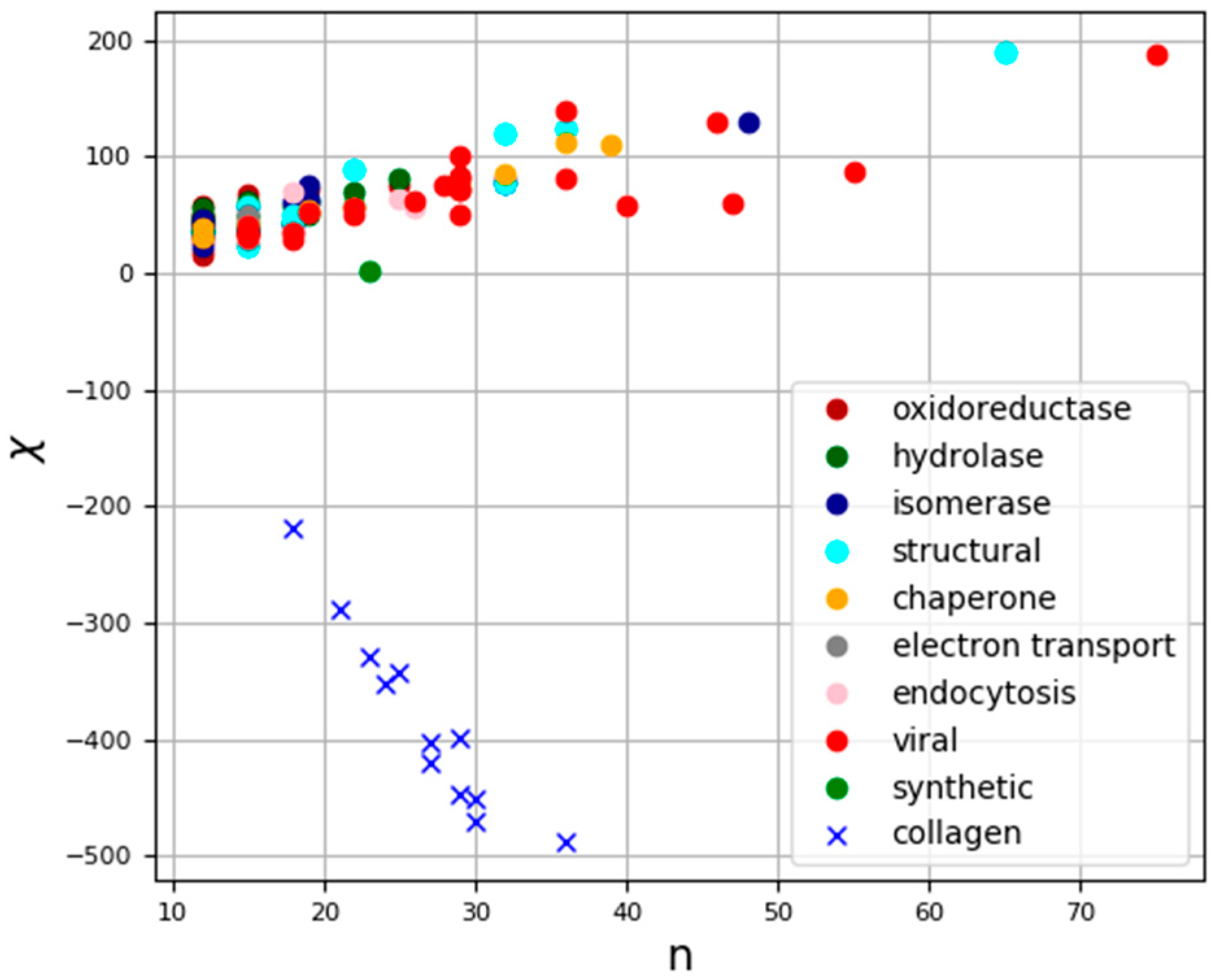{kind=link}
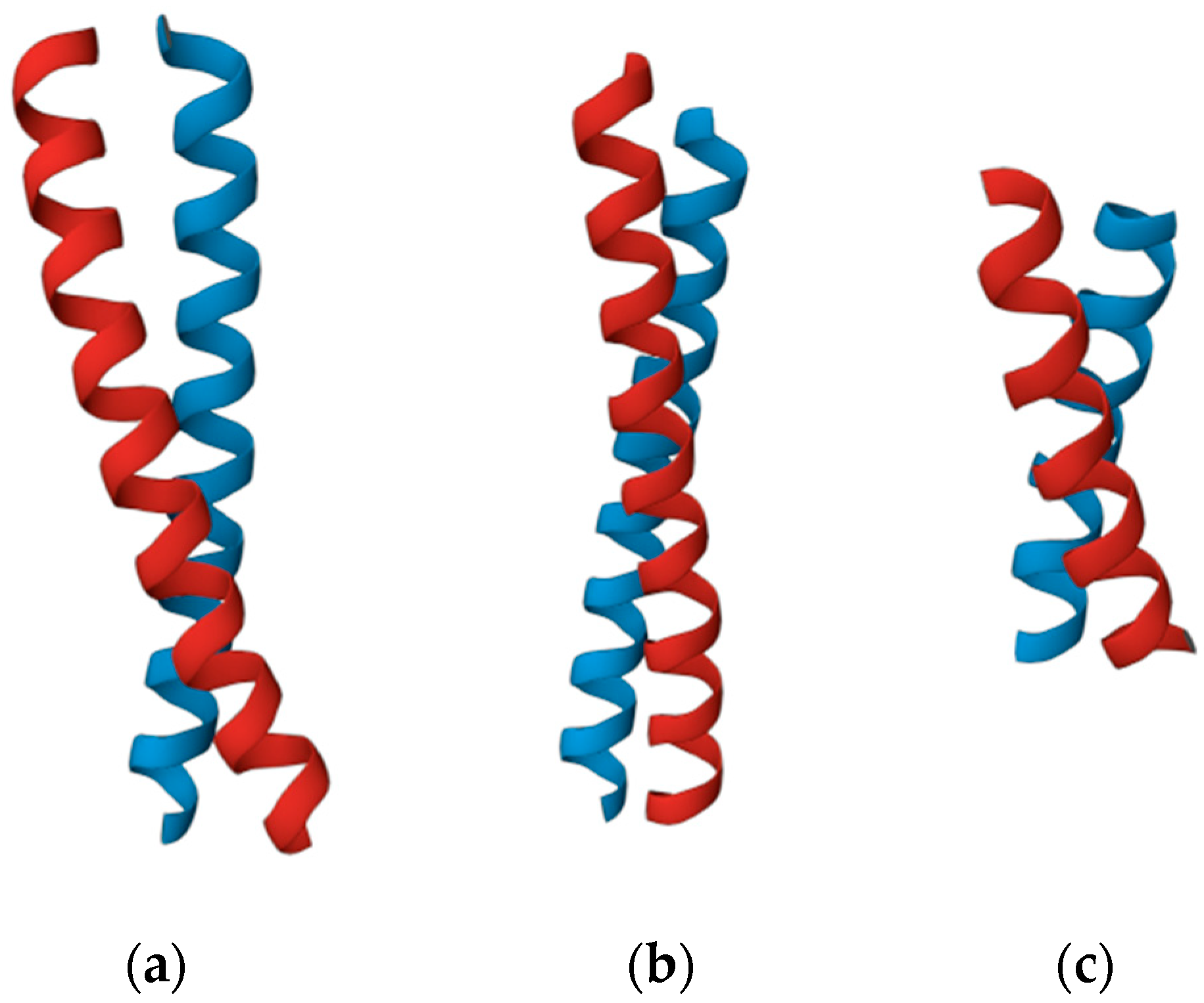{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mezey, P. Chirality Measures and Graph Representations. Comput. Math. Appl. 1997, 34, 105–112. [Google Scholar] [CrossRef]
- Petitjean, M. Chirality and Symmetry Measures: A Transdisciplinary Review. Entropy 2003, 5, 271–312. [Google Scholar] [CrossRef]
- Mezey, P.G. The proof of the metric properties of a fuzzy chirality measure of molecular electron density clouds. J. Mol. Str. Theochem. 1998, 455, 183–190. [Google Scholar] [CrossRef]
- Luzanov, A.V.; Nerukh, D. Complexity and chirality indices for molecular informatics: Differential geometry approach. Funct. Mater. 2005, 12, 55–64. [Google Scholar]
- Ramachandran, G.N.; Ramakrishnan, C.; Sasisekharan, V. Stereochemistry of polypeptide chain configurations. J. Mol. Biol. 1963, 7, 95–99. [Google Scholar] [CrossRef]
- Baruch-Shpigler, Y.; Wang, H.; Tuvi-Arad, I.; Avnir, D. Chiral Ramachandran Plots I: Glycine. Biochemistry 2017, 56, 5635–5643. [Google Scholar] [CrossRef]
- Zacharias, J.; Knapp, E.W. Geometry motivated alternative view on local protein backbone structures. Protein Sci. 2013, 22, 1669–1674. [Google Scholar] [CrossRef]
- Neukirch, S.; Goriely, A.; Hausrath, A.C. Chirality of coiled coils: Elasticity matters. Phys. Rev. Lett. 2008, 100, 038105. [Google Scholar] [CrossRef]
- Zheng, Y.; Mao, K.; Chen, S.; Zhu, H. Chirality Effects in Peptide Assembly Structures. Front. Bioeng. Biotechnol. 2021, 9, 703004. [Google Scholar] [CrossRef] [PubMed]
- Petitjean, M. Extreme asymmetry and chirality—A challenging quantification. Symmetry: Cult. Sci. 2020, 31, 439–447. [Google Scholar] [CrossRef]
- Wang, H.; Avnir, D.; Tuvi-Arad, I. Chiral Ramachandran Plots II: General trends and proteins chirality spectra. Biochemistry 2018, 57, 6395–6403. [Google Scholar] [CrossRef]
- Sidorova, A.; Bystrov, V.; Lutsenko, A.; Shpigun, D.; Belova, E.; Likhachev, I. Quantitative Assessment of Chirality of Protein Secondary Structures and Phenylalanine Peptide Nanotubes. Nanomaterials 2021, 11, 3299. [Google Scholar] [CrossRef]
- Tverdislov, V.A. Chirality as a Primary Switch of Hierarchical Levels in Molecular Biological Systems. Biophysics 2013, 58, 128–132. [Google Scholar] [CrossRef]
- Tverdislov, V.A.; Malyshko, E.V. On regularities of spontaneous formation of structural hierarchies in chiral systems of non-living and living nature. Phys. Usp. 2019, 62, 354. [Google Scholar] [CrossRef]
- Moutevelis, E.; Woolfson, D.N. A periodic table of coiled-coil protein structures. J. Mol. Biol. 2009, 385, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Lupas, A.N.; Gruber, M. The structure of α-helical coiled coils. Adv. Prot. Chem. 2005, 70, 37–78. [Google Scholar] [CrossRef]
- Rose, A.; Meier, I. Scaffolds, levers, rods and springs: Diverse cellular functions of long coiled-coil proteins. CMLS Cell. Mol. Life Sci. 2004, 61, 1996–2009. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.J.L.; Madera, M.; Armstrong, C.T.; Vincent, T.L.; Woolfson, D.N.; Gough, J. The evolution and structure prediction of coiled coils across all genomes. J. Mol. Biol. 2010, 403, 480–493. [Google Scholar] [CrossRef]
- Ohtaki, A.; Kida, H.; Miyata, Y.; Ide, N.; Yonezawa, A.; Arakawa, T.; Iizuka, R.; Noguchi, K.; Kita, A.; Odaka, M.; et al. Structure and molecular dynamics simulation of archaeal prefoldin: The molecular mechanism for binding and recognition of nonnative substrate proteins. J. Mol. Biol. 2008, 376, 1130–1141. [Google Scholar] [CrossRef]
- Perry, J.J.; Asaithamby, A.; Barnebey, A.; Kiamanesch, F.; Chen, D.J.; Han, S.; Tainer, J.A.; Yannone, S.M. Identification of a coiled coil in Werner syndrome protein that facilitates multimerization and promotes exonuclease processivity. J. Biol. Chem. 2010, 285, 25699–25707. [Google Scholar] [CrossRef]
- Strauss, H.M.; Keller, S. Pharmacological interference with protein-protein interactions mediated by coiled-coil motifs. In Protein–Protein Interactions as New Drug Targets, 1st ed.; Klussmann, E., Scott, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 461–482. [Google Scholar] [CrossRef]
- McFarlane, A.A.; Orriss, G.L.; Stetefeld, J. The use of coiled-coil proteins in drug delivery systems. Eur. J. Pharmacol. 2009, 625, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Crick, F.H.C. The packing of α-helices: Simple coiled-coils. Acta Crystallogr. 1953, 6, 689–697. [Google Scholar] [CrossRef]
- Walshaw, J.; Wooflson, D.N. Extended knobs-into holes packing in classical and complex coiled-coil assemblies. J. Struct. Biol. 2003, 144, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.M.; Arndt, K.M. Coiled-coil domains: Stability, specificity and biological implications. ChemBioChem 2004, 5, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H.; Cohen, C.; Parry, D.A.D. Heptad breaks in α-helical coiled coils: Stutters and stammers. Proteins 1996, 26, 134–145. [Google Scholar] [CrossRef]
- Hicks, M.R.; Holberton, D.V.; Kowalczyk, C.; Woolfson, D.N. Coiled-coil assembly by peptides with non-heptad sequence motifs. Fold. Des. 1997, 2, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Grigoryan, G.; DeGrado, W.F. The accommodation index measures the perturbation associated with insertions and deletions in coiled-coils: Application to understand signaling in histidine kinases. Protein Sci. 2017, 26, 414–435. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Kaur, J.; Reinhardt, D.P. Extracellular Matrix (ECM) Molecules. In Stem Cell Biology and Tissue Engineering in Dental Sciences, 1st ed.; Vishwakarma, A., Sharpe, P., Songtao, S., Ramalingam, M., Eds.; Elseveir: London, UK, 2014; pp. 27–34. [Google Scholar]
- Crick, F.H.C. The Fourier Transform of a Coiled-Coil. Acta Crystallogr. 1953, 6, 685–689. [Google Scholar] [CrossRef]
- Offer, G.; Hicks, M.R.; Woolfson, D.N. Generalized Crick equations for modeling noncanonical coiled coils. J. Struct. Biol. 2002, 137, 41–53. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P. Conformation in Fibrous Proteins and Related Synthetic Polypeptides, 1st ed.; Academic Press: New York, NY, USA, 1973; p. 628. [Google Scholar]
- Phillips, G.N. What Is the Pitch of the α-Helical Coiled Coil? Proteins 1992, 14, 425–429. [Google Scholar] [CrossRef]
- Sidorova, A.E.; Malyshko, E.V.; Lutsenko, A.O.; Shpigun, D.K.; Bagrova, O.E. Protein helical structures: Defining handedness and localization features. Symmetry 2021, 13, 879. [Google Scholar] [CrossRef]
- Kobe, B.; Center, R.J.; Kemp, B.E.; Poumbourios, P. Crystal structure of human T-cell leukemia virus type 1 gp21 ectodomain crystallized as a maltose-binding protein chimera reveals structural evolution of retroviral transmembrane proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 4319–4324. [Google Scholar] [CrossRef] [PubMed]
- Strelkov, S.V.; Schumacher, J.; Burkhard, P.; Aebi, U.; Herrmann, H. Crystal Structure of the Human Lamin A Coil 2B Dimer: Implications for the Head-to-tail Association of Nuclear Lamins. J. Mol. Biol. 2004, 343, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- RCSB PDB. Available online: https://www.rcsb.org/ (accessed on 5 May 2023).
- The CC+ Database. Available online: http://coiledcoils.chm.bris.ac.uk/ccplus/search/ (accessed on 5 May 2023).
- Honnappa, S.; John, C.M.; Kostrewa, D.; Winkler, F.K.; Steinmetz, M.O. Structural insights into the EB1–APC interaction. EMBO J. 2005, 24, 261–269. [Google Scholar] [CrossRef]
- Samatey, F.A.; Imada, K.; Nagashima, S.; Vonderviszt, F.; Kumasaka, T.; Yamamoto, M.; Namba, K. Structure of the bacterial flagellar protofilament and implications for a switch for supercoiling. Nature 2001, 410, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Laponogov, I.; Veselkov, D.A.; Sohi, M.K.; Pan, X.-S.; Achari, A.; Yang, C.; Sanderson, M.R. Breakage-Reunion Domain of Streptococcus pneumoniae Topoisomerase IV: Crystal Structure of a Gram-Positive Quinolone Target. PLoS ONE 2007, 2, e301. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lutsenko, A.; Sidorova, A.; Shpigun, D.; Belova, E.; Tverdislov, V. A Method for Calculating the Sign and Degree of Chirality of Supercoiled Protein Structures. Symmetry 2023, 15, 2051. https://doi.org/10.3390/sym15112051
Lutsenko A, Sidorova A, Shpigun D, Belova E, Tverdislov V. A Method for Calculating the Sign and Degree of Chirality of Supercoiled Protein Structures. Symmetry. 2023; 15(11):2051. https://doi.org/10.3390/sym15112051
Chicago/Turabian StyleLutsenko, Aleksey, Alla Sidorova, Denis Shpigun, Ekaterina Belova, and Vsevolod Tverdislov. 2023. "A Method for Calculating the Sign and Degree of Chirality of Supercoiled Protein Structures" Symmetry 15, no. 11: 2051. https://doi.org/10.3390/sym15112051
APA StyleLutsenko, A., Sidorova, A., Shpigun, D., Belova, E., & Tverdislov, V. (2023). A Method for Calculating the Sign and Degree of Chirality of Supercoiled Protein Structures. Symmetry, 15(11), 2051. https://doi.org/10.3390/sym15112051