



Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Experimental Design and Sample Collection
2.3. Morphometric Analyses
2.4. Selection Analyses
3. Results
3.1. The Variation in Floral Organs’ Color and Size in Pollinated and Non-Pollinated Flowers
3.2. The Pattern of Floral Organs’ Shape Variation
3.3. Analyses of the Asymmetry of Pollinated and Non-Pollinated Floral Organs
3.4. Phenotypic Selection on Flower Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harder, L.D.; Barrett, S.C.H. Pollen Dispersal and Mating Patterns in Animal-Pollinated Plants. In Floral Biology, 1st ed.; Lloyd, D.G., Barrett, S.C.H., Eds.; Springer: Boston, MA, USA, 1996; pp. 140–190. [Google Scholar]
- Harder, L.D.; Barrett, S.C.H. Pollen and ovule fates and reproductive performance by flowering plants. In Ecology and Evolution of Flowers, 1st ed.; Harder, L.D., Routley, M.B., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2006; pp. 61–80. [Google Scholar]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Simpson, B.B.; Neff, J.L. Floral rewards: Alternatives to pollen and nectar. Ann. Missouri Bot. Gard. 1981, 68, 301–322. [Google Scholar] [CrossRef]
- Blarer, A.; Keasar, T.; Shmida, A. Possible mechanisms for the formation of flower size preferences by foraging bumblebees. Ethology 2002, 108, 341–351. [Google Scholar] [CrossRef]
- Knauer, A.C.; Schiestl, F.P. Bees use honest floral signals as indicators of reward when visiting flowers. Ecol. Lett. 2015, 18, 135–143. [Google Scholar] [CrossRef]
- Ito, K.; Suzuki, M.F.; Mochizuki, K. Evolution of honest reward signal in flowers. Proc. R. Soc. B 2021, 288, 20202848. [Google Scholar] [CrossRef]
- Spaethe, J.; Tautz, J.; Chittka, L. Visual constraints in foraging bumblebees: Flower size and color affect search time and flight behavior. Proc. Natl. Acad. Sci. USA 2001, 98, 3898–3903. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Johnson, S.D. Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 2013, 28, 307–315. [Google Scholar] [CrossRef]
- Gómez, J.M.; Torices, R.; Lorite, J.; Klingenberg, C.P.; Perfectti, F. The role of pollinators in the evolution of corolla shape variation, disparity and integration in a highly diversified plant family with a conserved floral bauplan. Ann. Bot. 2016, 117, 889–904. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Jürgens, A. Deceptive behavior in plants. II. Food deception by plants: From generalized systems to specialized floral mimicry. In Plant-Environment Interaction, 1st ed.; Baluška, F., Ed.; Springer: Berlin, Germany, 2009; pp. 223–246. [Google Scholar]
- Johnson, S.D.; Schiestl, F.P. Floral Mimicry, 1st ed.; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Pellissier, L.; Vittoz, P.; Internicola, A.I.; Gigord, L.D.B. Generalized food-deceptive orchid species flower earlier and occur at lower altitudes than rewarding ones. J. Plant Ecol. 2010, 3, 243–250. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235. [Google Scholar] [CrossRef]
- Juillet, N.; Scopece, G. Does floral trait variability enhance reproductive success in deceptive orchids? Perspect. Plant Ecol. Evol. Syst. 2010, 12, 317–322. [Google Scholar] [CrossRef]
- Renner, S.S. Rewardless flowers in the angiosperms and the role of insect cognition in their evolution. In Plant-Pollinator Interactions: From Specialization to Generalization, 1st ed.; Waser, N.M., Olletron, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2006; pp. 123–144. [Google Scholar]
- Souto-Vilarós, D.; Vuleta, A.; Jovanović, S.M.; Budečević, S.; Wang, H.; Sapir, Y.; Imbert, E. Are pollinators the agents of selection on flower colour and size in irises? Oikos 2018, 127, 834–846. [Google Scholar] [CrossRef]
- Engel, E.C.; Irwin, R.E. Linking pollinator visitation rate and pollen receipt. Am. J. Bot. 2003, 90, 1612–1618. [Google Scholar] [CrossRef]
- Makino, T.T.; Sakai, S. Experience changes pollinator responses to floral display size: From size-based to reward-based foraging. Funct. Ecol. 2007, 21, 854–863. [Google Scholar] [CrossRef]
- Fenster, C.B.; Cheely, G.; Dudash, M.R.; Reynolds, R.J. Nectar reward and advertisement in hummingbird-pollinated Silene virginica (Caryophyllaceae). Am. J. Bot. 2006, 93, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Frey, F.M.; Bukoski, M. Floral symmetry is associated with flower size and pollen production but not insect visitation rates in Geranium robertianum (Geraniaceae). Plant Species Biol. 2014, 29, 272–280. [Google Scholar] [CrossRef]
- Møller, A.P.; Eriksson, M. Patterns of fluctuating asymmetry in flowers: Implications for sexual selection in plants. J. Evol. Biol. 1994, 7, 97–113. [Google Scholar] [CrossRef]
- Møller, A.P. Bumblebee preference for symmetrical flowers. Proc. Natl. Acad. Sci. USA 1995, 92, 2288–2292. [Google Scholar] [CrossRef]
- Midgley, J.J.; Johnson, S.D. Some pollinators do not prefer symmetrically marked or shaped daisy (Asteraceae) flowers. Evol. Ecol. 1998, 12, 123–126. [Google Scholar] [CrossRef]
- Frey, F.M.; Davis, R.; Delph, L.F. Manipulation of floral symmetry does not affect seed production in Impatiens pallida. Int. J. Plant Sci. 2005, 166, 659–662. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry analyses revisited. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 279–319. [Google Scholar]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [PubMed]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Tucić, B.; Budečević, S.; Manitašević Jovanović, S.; Vuleta, A.; Klingenberg, C.P. Phenotypic plasticity in response to environmental heterogeneity contributes to fluctuating asymmetry in plants: First empirical evidence. J. Evol. Biol. 2018, 31, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Lavi, R.; Sapir, Y. Are pollinators the agents of selection for the extreme large size and dark color in Oncocyclus irises? New Phytol. 2015, 205, 369–377. [Google Scholar] [CrossRef]
- Mathew, B. The Iris, 1st ed.; Timber Press: Portland, OR, USA, 1981. [Google Scholar]
- Guo, J.; Wilson, C.A. Organization, available space and organ morphology within floral buds of Iris (Iridaceae). Flora 2018, 249, 67–76. [Google Scholar] [CrossRef]
- Remizowa, M.V.; Rudall, P.J.; Choob, V.V.; Sokoloff, D.D. Racemose inflorescences of monocots: Structural and morphogenetic interaction at the flower/inflorescence level. Ann. Bot. 2013, 112, 1553–1566. [Google Scholar] [CrossRef] [PubMed]
- Radović, S.; Urošević, A.; Hočevar, K.; Vuleta, A.; Manitašević Jovanović, S.; Tucić, B. Geometric morphometrics of functionally distinct floral organs in Iris pumila: Analyzing patterns of symmetric and asymmetric shape variations. Arch. Biol. Sci. 2017, 69, 223–231. [Google Scholar] [CrossRef]
- Budečević, S.; Manitašević Jovanović, S.; Vuleta, A.; Tucić, B.; Klingenberg, C.P. Directional asymmetry and direction-giving factors: Lessons from flowers with complex symmetry. Evol. Dev. 2022, 24, 92–108. [Google Scholar] [CrossRef]
- Ashman, T.L.; Knight, T.M.; Steets, J.A.; Amarasekare, P.; Burd, M.; Campbell, D.R.; Dudash, M.R.; Johnston, M.O.; Mazer, S.J.; Mitchell, R.J. Pollen limitation of plant reproduction: Ecological and evolutionary causes and consequences. Ecology 2004, 85, 2408–2421. [Google Scholar] [CrossRef]
- Ackerman, J.D.; Cuevas, A.A.; Hof, D. Are deception-pollinated species more variable than those offering a reward? Plant Syst. Evol. 2011, 293, 91–99. [Google Scholar] [CrossRef]
- Tarasjev, A. Impact of genet size and flowering stage on fruit set in Iris pumila L. clones in wild. Acta Oecologica 2005, 27, 93–98. [Google Scholar] [CrossRef]
- Walsh, R.P.; Michaels, H.J. When it pays to cheat: Examining how generalized food deception increases male and female fitness in a terrestrial orchid. PLoS ONE 2017, 12, e0171286. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Marín, M.; Dorken, M.E.; Barrett, S.C.H. The ecological and evolutionary consequences of clonality for plant mating. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 193–213. [Google Scholar] [CrossRef]
- Vuleta, A.; Hočevar, K.; Manitašević Jovanović, S.; Raičević, J.; Plećaš, M. Assessment of Iris pumila L. pollinator and florivore diversity in a common garden: A pan-trap experiment. In Proceedings of the 4th International Conference on Plant Biology (23rd SPPS Meeting), Belgrade, Serbia, 6–8 October 2022. [Google Scholar]
- Tucić, B.; Milojković, S.; Tarasjev, A.; Vijčić, S. The influence of climatic factors on clonal diversity in a population of Iris pumila. Oikos 1989, 56, 115–120. [Google Scholar] [CrossRef]
- Lambert, P.; Carron, T. Symbolic fusion of luminance-hue-chroma features for region segmentation. Pattern Recognit. 1999, 32, 1857–1872. [Google Scholar] [CrossRef]
- Sletvold, N.; Trunschke, J.; Smit, M.; Verbeek, J.; Ågren, J. Strong pollinator-mediated selection for increased flower brightness and contrast in a deceptive orchid. Evolution 2016, 70, 716–724. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis: With Applications in R, 1st ed.; John Wiley & Sons: Chichester, UK, 2016. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Lande, R.; Arnold, S.J. The measurement of selection on correlated characters. Evolution 1983, 37, 1210–1226. [Google Scholar] [CrossRef]
- Stinchcombe, J.R.; Agrawal, A.F.; Hohenlohe, P.A.; Arnold, S.J.; Blows, M.W. Estimating nonlinear selection gradients using quadratic regression coefficients: Double or nothing? Evolution 2008, 62, 2435–2440. [Google Scholar] [CrossRef]
- Weiß, C.H. Statsoft, Inc., Tulsa, OK: STATISTICA, version 8. Asta-Adv. Stat. Anal. 2007, 91, 339–341. [Google Scholar] [CrossRef]
- Kagawa, K.; Takimoto, G. Inaccurate color discrimination by pollinators promotes evolution of discrete color polymorphism in food-deceptive flowers. Am. Nat. 2016, 187, 194–204. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Dyer, A.G.; Kevan, P.G.; Lunau, K. Functional significance of the optical properties of flowers for visual signalling. Ann Bot. 2019, 123, 263–276. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Spaethe, J. Caution with colour calculations: Spectral purity is a poor descriptor of flower colour visibility. Ann Bot. 2022, 130, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Barišić Klisarić, N.; Miljković, D.; Avramov, S.; Živković, U.; Tarasjev, A. Radial and bilateral fluctuating asymmetry of Iris pumila flowers as indicators of environmental stress. Symmetry 2019, 11, 818. [Google Scholar] [CrossRef]
- Tucić, B.; Miljković, D. Fluctuating asymmetry of floral organ traits in natural populations of Iris pumila from contrasting light habitats. Plant Species Biol. 2010, 25, 173–184. [Google Scholar] [CrossRef]
- Segal, B.; Sapir, Y.; Carmel, Y. Fragmentation and pollination crisis in the self-incompatible Iris bismarckiana (Iridaceae), with implications for conservation. Isr. J. Ecol. Evol. 2006, 52, 111–122. [Google Scholar] [CrossRef]
- Pellegrino, G. Pollinator limitation on reproductive success in Iris tuberosa. AoB Plants 2015, 7, plu089. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.; Flick, A.J.; Bauer, A.A. Phenotypic selection on flower color and floral display size by three bee species. Front. Plant Sci. 2021, 11, 587528. [Google Scholar] [CrossRef]
- Caruso, C.M.; Eisen, K.E.; Martin, R.A.; Sletvold, N. A meta-analysis of the agents of selection on floral traits. Evolution 2019, 73, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Trunschke, J.; Lunau, K.; Pyke, G.H.; Ren, Z.-X.; Wang, H. Flower Color Evolution and the Evidenceof Pollinator-Mediated Selection. Front. Plant Sci. 2021, 12, 617851. [Google Scholar] [CrossRef] [PubMed]
- Scopece, G.; Juillet, N.; Lexer, C.; Cozzolino, S. Fluctuating selection across years and phenotypic variation in food-deceptive orchids. PeerJ 2017, 5, e3704. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Burczyk, J. Selection on floral traits through male fertility in a natural plant population. Evol. Ecol. 2008, 22, 39–54. [Google Scholar] [CrossRef]
- Lehrer, M.; Horridge, G.A.; Zhang, S.W.; Gadagkar, R. Shape vision in bees: Innate preference for flower-like patterns. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1995, 347, 123–137. [Google Scholar]
- Lunau, K.; Wacht, S.; Chittka, L. Colour choices of naive bumble bees and their implications for colour perception. J. Comp. Physiol. A 1996, 178, 477–489. [Google Scholar] [CrossRef]
- Gumbert, A. Color choices by bumble bees (Bombus terrestris): Innate preferences and generalization after learning. Behav. Ecol. Sociobiol. 2000, 48, 36–43. [Google Scholar] [CrossRef]
- Raine, N.E.; Ings, T.C.; Dornhaus, A.; Saleh, N.; Chittka, L. Adaptation, genetic drift, pleiotropy, and history in the evolution of bee foraging behavior. Adv. Study Behav. 2006, 36, 305–354. [Google Scholar]
- Lehrer, M. Shape perception in the honeybee: Symmetry as a global framework. Int. J. Plant Sci. 1999, 160, S51–S65. [Google Scholar] [CrossRef]
- La Rosa, R.J.; Conner, J.K. Floral function: Effects of traits on pollinators, male and female pollination success, and female fitness across three species of milkweeds (Asclepias). Am. J. Bot. 2017, 104, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Morinaga, S.I.; Sakai, S. Functional differentiation in pollination processes between the outer and inner perianths in Iris gracilipes (Iridaceae). Botany 2006, 84, 164–171. [Google Scholar] [CrossRef]




{kind=link}
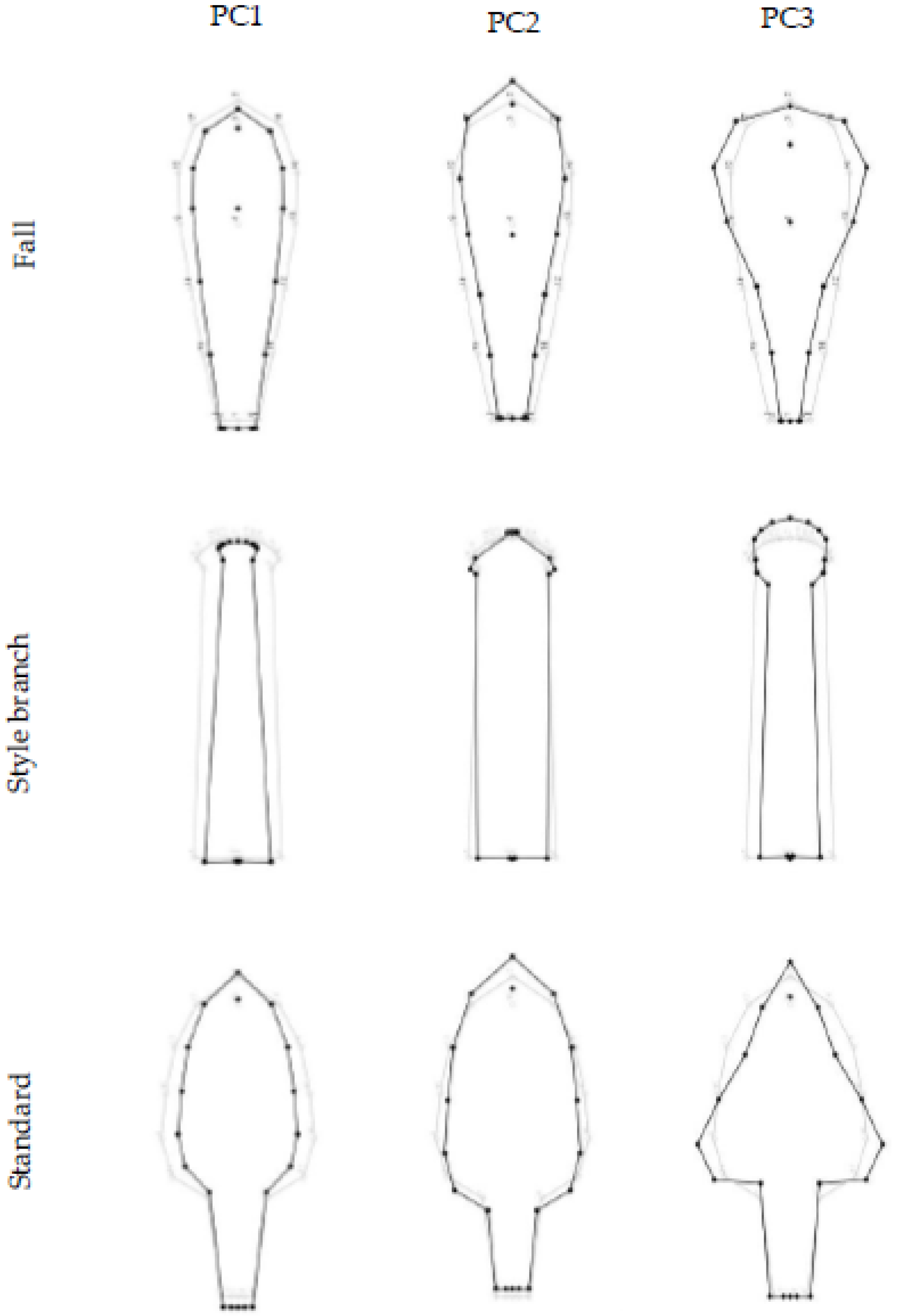{kind=link}
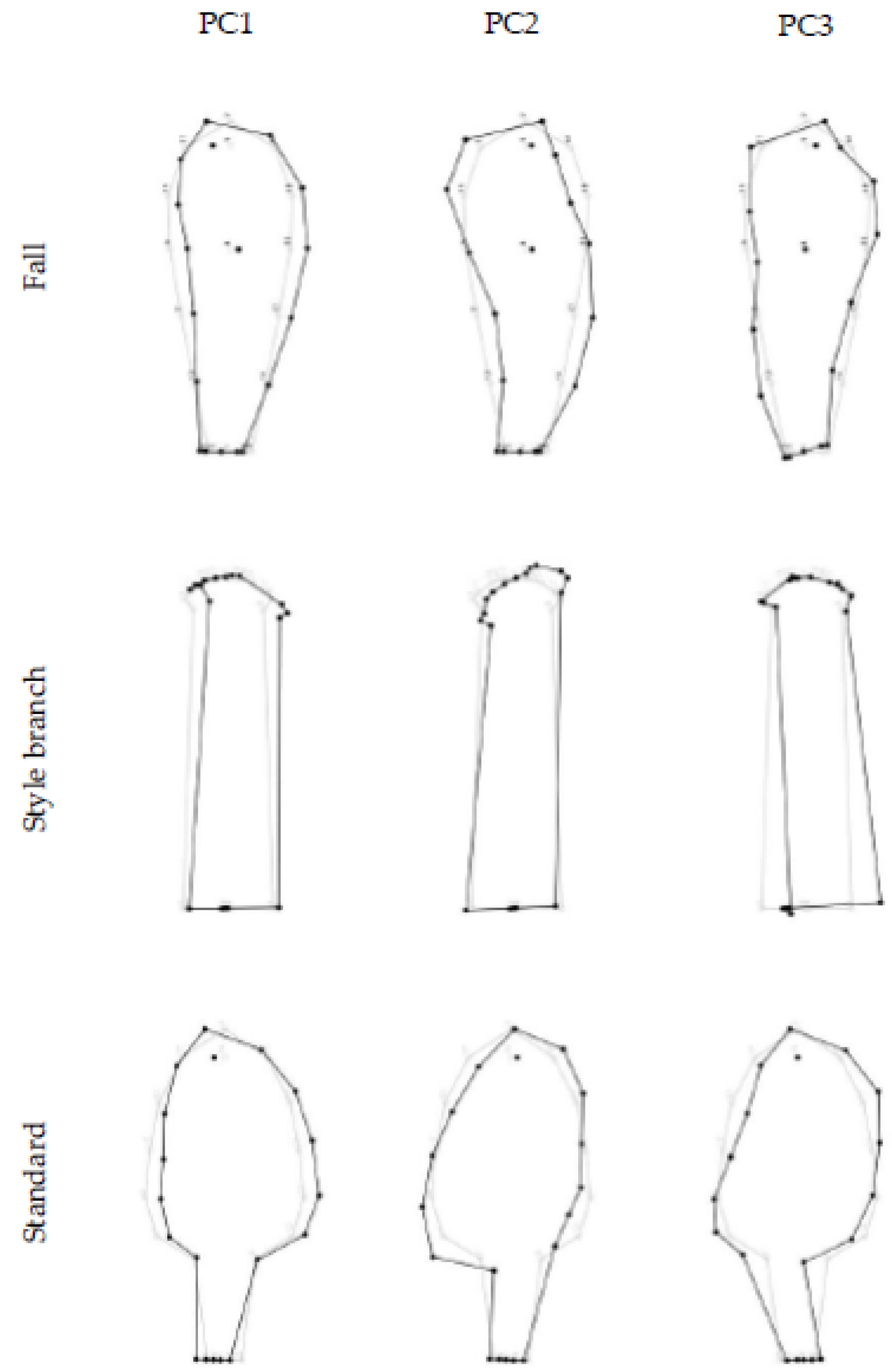{kind=link}
| Pollinated Flower | Non-Pollinated Flower | ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trait | n | SE | CV % | n | SE | CV % | F | p | ||
| Achromatic brightness | 193 | 0.266 | 0.005 | 26.19 | 191 | 0.251 | 0.005 | 24.85 | 5.143 | 0.024 |
| Flower stem height | 193 | 97.42 | 1.49 | 21.29 | 191 | 87.54 | 1.26 | 19.89 | 25.58 | <0.0001 |
| CS fall | 193 | 338.7 | 3.0 | 12.20 | 191 | 321.5 | 2.8 | 12.18 | 17.06 | <0.0001 |
| CS style branch | 193 | 198.2 | 1.4 | 9.62 | 191 | 192.2 | 1.4 | 10.09 | 9.19 | 0.003 |
| CS standard | 193 | 335.1 | 3.1 | 13.00 | 191 | 315.3 | 3.0 | 13.12 | 20.60 | <0.0001 |
| Fall-style branch CS ratio | 193 | 1.713 | 0.013 | 10.39 | 191 | 1.676 | 0.011 | 9.45 | 4.43 | 0.036 |
| Pollinated Floral Organs | Non-Pollinated Floral Organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Effect | SS | MS | df | F | p | SS | MS | df | F | p |
| Fall | ||||||||||
| Individual | 0.73483 | 0.00024 | 3056 | 7.51 | <0.0001 | 0.68965 | 0.00023 | 2992 | 8.34 | <0.0001 |
| Side | 0.05519 | 0.00345 | 16 | 107.7 | <0.0001 | 0.04922 | 0.00308 | 16 | 111.3 | <0.0001 |
| Ind * Side | 0.09787 | 0.00003 | 3056 | 5.39 | <0.0001 | 0.08273 | 0.00003 | 2992 | 4.90 | <0.0001 |
| Error 1 | 0.03593 | 0.00001 | 6048 | 0.03338 | 0.00001 | 5920 | ||||
| Style branch | ||||||||||
| Individual | 0.47477 | 0.00016 | 3056 | 6.64 | <0.0001 | 0.43848 | 0.00014 | 3040 | 5.25 | <0.0001 |
| Side | 0.00449 | 0.00028 | 16 | 11.99 | <0.0001 | 0.00461 | 0.00029 | 16 | 10.49 | <0.0001 |
| Ind * Side | 0.07150 | 0.00002 | 3056 | 1.80 | <0.0001 | 0.08352 | 0.00003 | 3040 | 1.82 | <0.0001 |
| Error 1 | 0.08004 | 0.00001 | 6144 | 0.09210 | 0.00002 | 6112 | ||||
| Standard | ||||||||||
| Individual | 1.13133 | 0.00035 | 3264 | 7.68 | <0.0001 | 1.26642 | 0.00039 | 3247 | 8.59 | <0.0001 |
| Side | 0.07219 | 0.00425 | 17 | 94.11 | <0.0001 | 0.06520 | 0.00384 | 17 | 84.46 | <0.0001 |
| Ind * Side | 0.14727 | 0.00005 | 3264 | 1.11 | 0.0002 | 0.14746 | 0.00005 | 3247 | 1.19 | <0.0001 |
| Error 1 | 0.26593 | 0.00004 | 6562 | 0.24915 | 0.00004 | 6528 | ||||
| Trait | β ± SE | p | γ ± SE | p |
|---|---|---|---|---|
| Achromatic brightness | 0.074 ± 0.050 | 0.143 | −0.052 ± 0.246 | 0.915 |
| Flower stem height | 0.176 ± 0.069 | 0.011 | 0.094 ± 0.318 | 0.883 |
| CS fall | −0.392 ± 0.686 | 0.568 | 3.273 ± 5.116 | 0.749 |
| CS style branch | 0.276 ± 0.532 | 0.604 | −2.271 ± 3.642 | 0.755 |
| CS standard | 0.142 ± 0.132 | 0.282 | −1.885 ± 1.106 | 0.395 |
| Fall-style branch CS ratio | 0.278 ± 0.532 | 0.602 | 1.053 ± 3.849 | 0.891 |
| Fall shape | ||||
| PC1 | −0.014 ± 0.052 | 0.791 | 0.022 ± 0.052 | 0.830 |
| PC2 | 0.012 ± 0.052 | 0.821 | 0.089 ± 0.055 | 0.418 |
| PC3 | 0.151 ± 0.052 | 0.004 | 0.051 ± 0.052 | 0.627 |
| Style branch shape | ||||
| PC1 | 0.048 ± 0.051 | 0.349 | −0.026 ± 0.053 | 0.807 |
| PC2 | −0.065 ± 0.051 | 0.203 | 0.208 ± 0.052 | 0.047 |
| PC3 | 0.090 ± 0.051 | 0.080 | 0.199 ± 0.052 | 0.055 |
| Standard shape | ||||
| PC1 | −0.055 ± 0.051 | 0.285 | −0.105 ± 0.052 | 0.311 |
| PC2 | 0.021 ± 0.051 | 0.680 | −0.016 ± 0.052 | 0.878 |
| PC3 | 0.038 ± 0.051 | 0.456 | −0.034 ± 0.052 | 0.744 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budečević, S.; Hočevar, K.; Manitašević Jovanović, S.; Vuleta, A. Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators. Symmetry 2023, 15, 1149. https://doi.org/10.3390/sym15061149
Budečević S, Hočevar K, Manitašević Jovanović S, Vuleta A. Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators. Symmetry. 2023; 15(6):1149. https://doi.org/10.3390/sym15061149
Chicago/Turabian StyleBudečević, Sanja, Katarina Hočevar, Sanja Manitašević Jovanović, and Ana Vuleta. 2023. "Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators" Symmetry 15, no. 6: 1149. https://doi.org/10.3390/sym15061149
APA StyleBudečević, S., Hočevar, K., Manitašević Jovanović, S., & Vuleta, A. (2023). Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators. Symmetry, 15(6), 1149. https://doi.org/10.3390/sym15061149