
Oligomeric Symmetry of Purine Nucleoside Phosphorylases


Abstract
:1. Introduction
2. Materials and Methods
3. Results
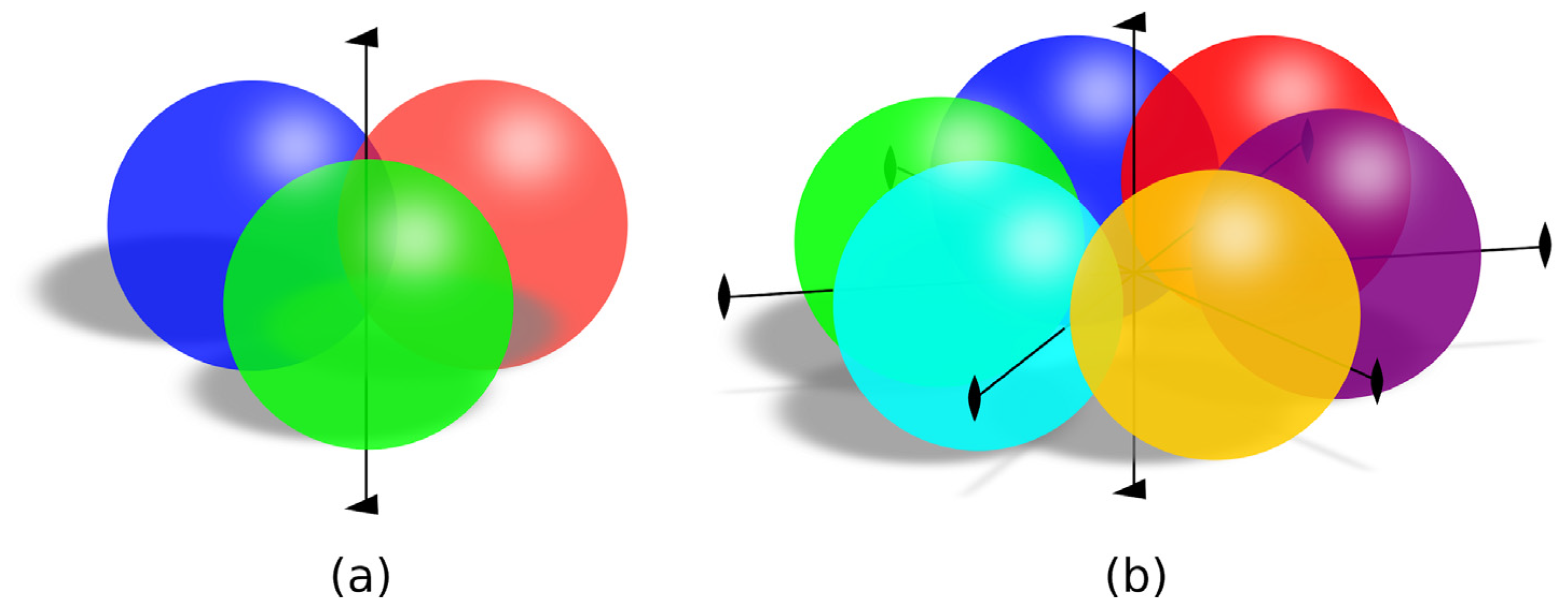
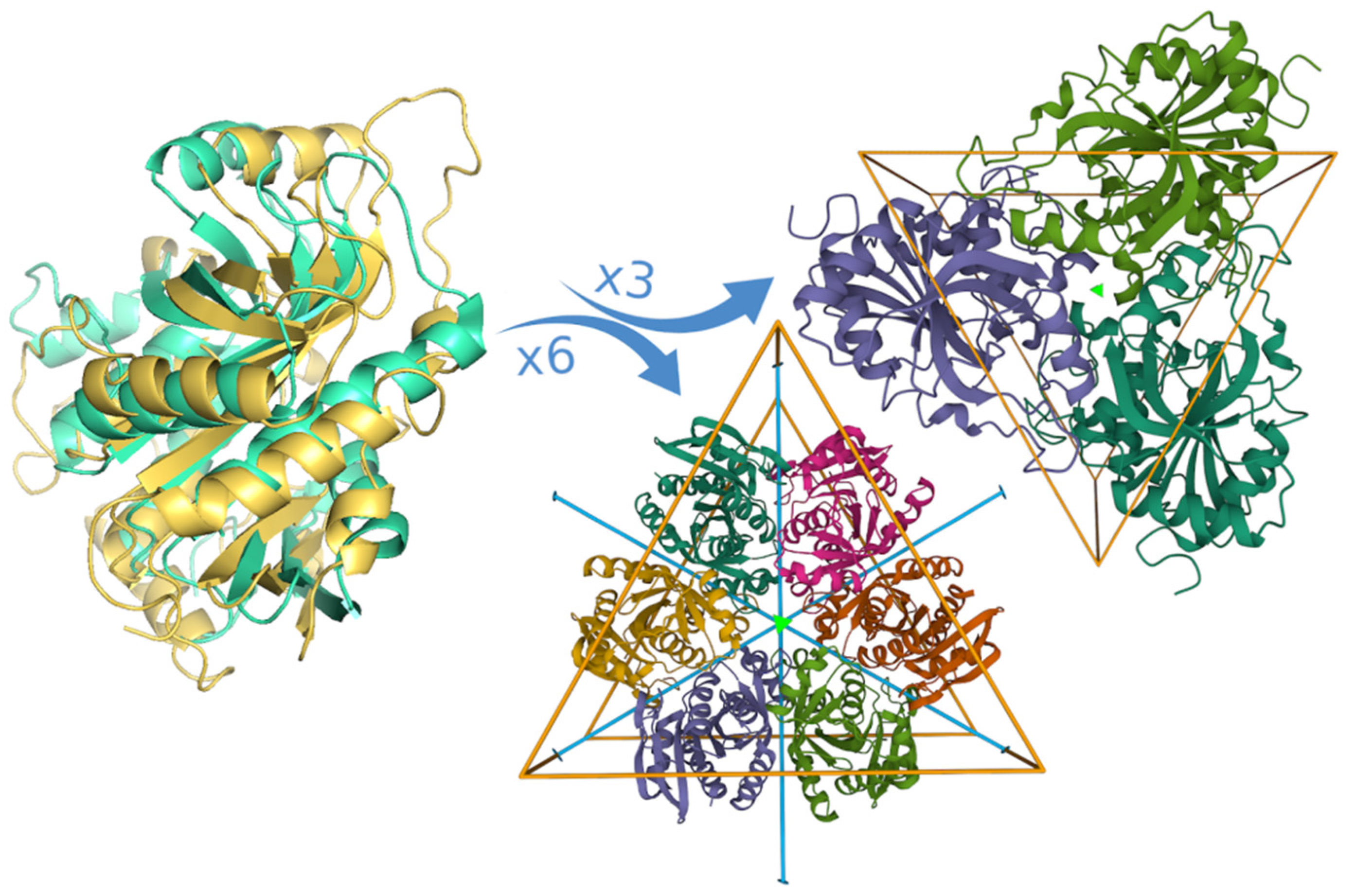
3.1. Structural Alignment
3.2. Multiple Sequence Alignment
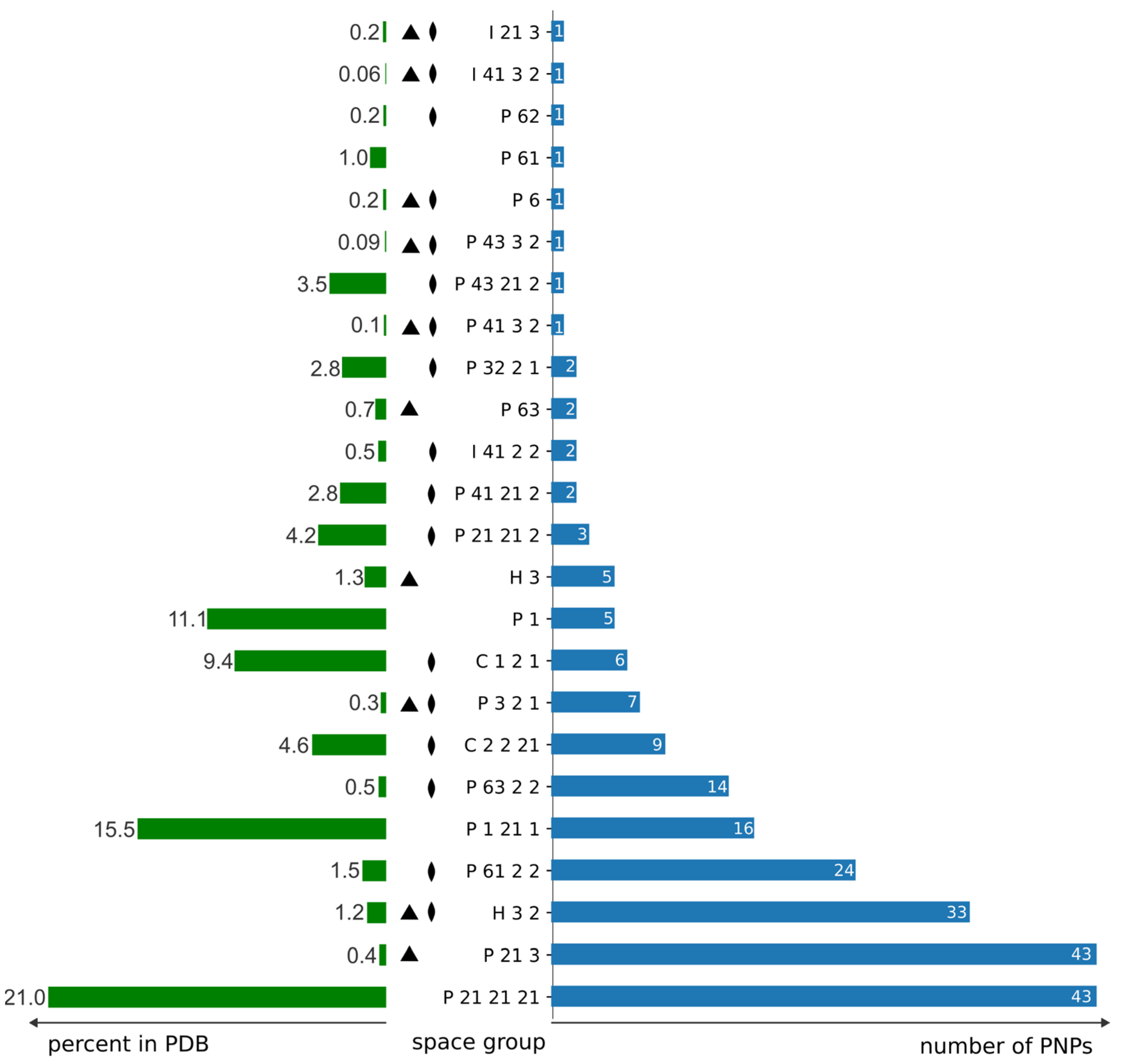
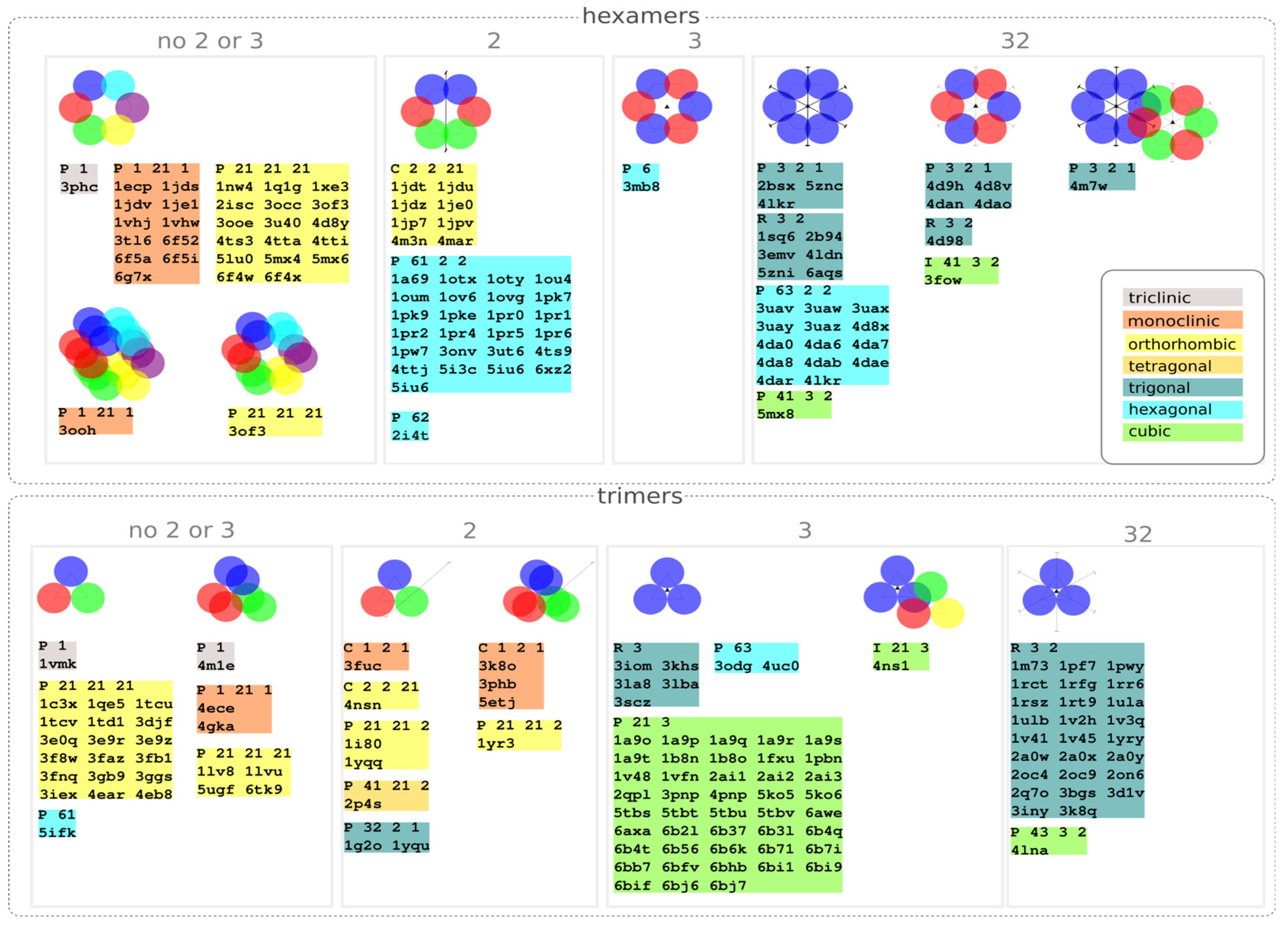
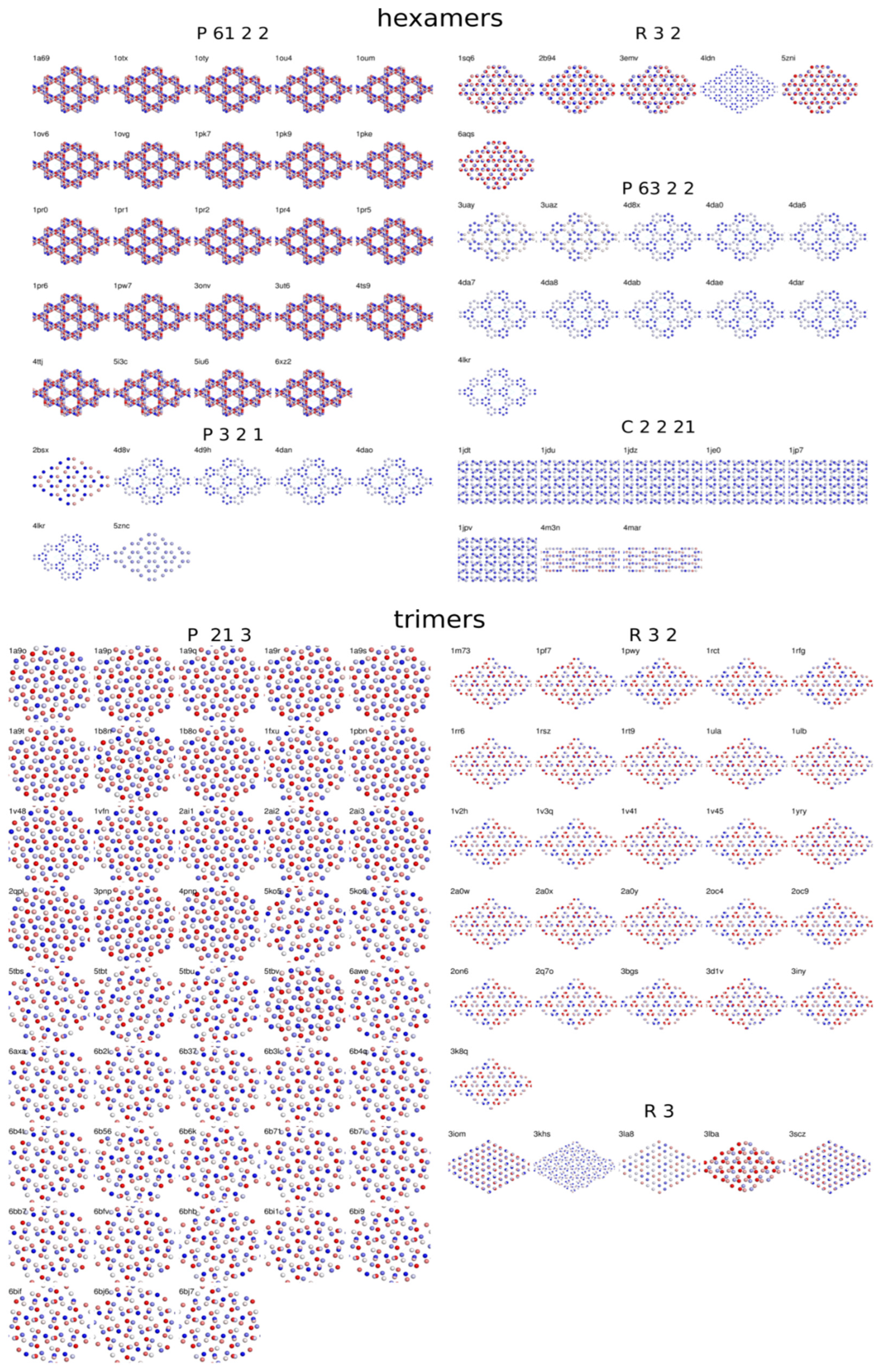
3.3. Distribution of Space Groups
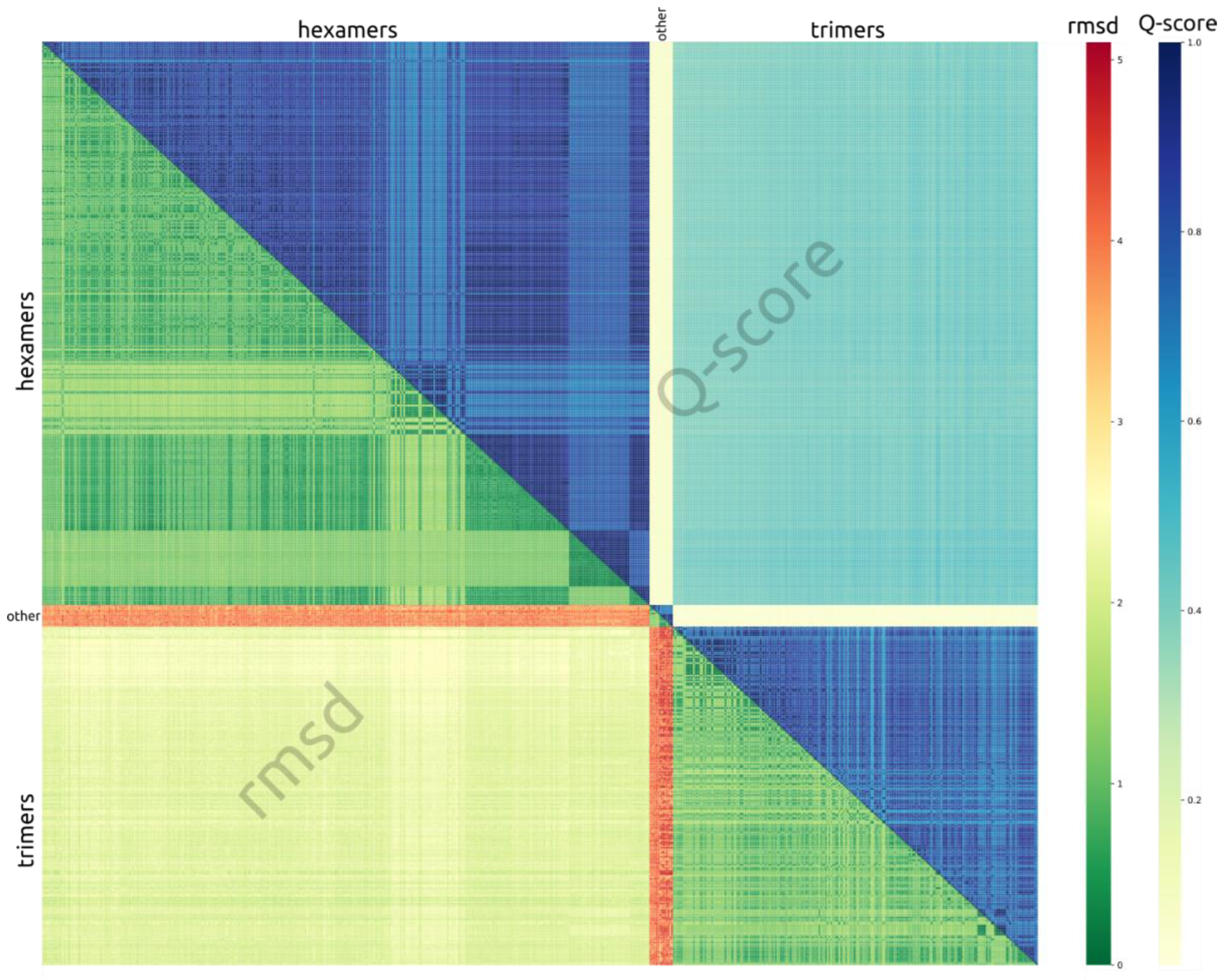
3.4. Crystal Packing Similarities
3.5. Contacts between Subunits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goodsell, D.S.; Olson, A.J. Structural Symmetry and Protein Function. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 105–153. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.W.; Prlić, A.; Altunkaya, A.; Bi, C.; Bradley, A.R.; Christie, C.H.; Di Costanzo, L.; Duarte, J.M.; Dutta, S.; Feng, Z.; et al. The RCSB Protein Data Bank: Integrative View of Protein, Gene and 3D Structural Information. Nucleic Acids Res. 2017, 45, D271–D281. [Google Scholar] [CrossRef] [PubMed]
- Burley, S.K.; Berman, H.M.; Kleywegt, G.J.; Markley, J.L.; Nakamura, H.; Velankar, S. Protein Data Bank (PDB): The Single Global Macromolecular Structure Archive. In Protein Crystallography: Methods in Molecular Biology; Wlodawer, A., Dauter, Z., Jaskolski, M., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1607, pp. 627–641. ISBN 978-1-4939-6998-2. [Google Scholar]
- Marsh, J.A.; Teichmann, S.A. Structure, Dynamics, Assembly, and Evolution of Protein Complexes. Annu. Rev. Biochem. 2015, 84, 551–575. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Prilusky, J.; Levy, E.D. QSalignWeb: A Server to Predict and Analyze Protein Quaternary Structure. Front. Mol. Biosci. 2022, 8, 787510. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, S.; Duarte, J.M.; Prlić, A.; Goksuluk, D.; Zararsiz, G.; Saracbasi, O.; Burley, S.K.; Rose, P.W. Investigation of Protein Quaternary Structure via Stoichiometry and Symmetry Information. PLoS ONE 2018, 13, e0197176. [Google Scholar] [CrossRef]
- Kojić-Prodić, B.; Štefanić, Z. Symmetry versus Asymmetry in the Molecules of Life: Homomeric Protein Assemblies. Symmetry 2010, 2, 884–906. [Google Scholar] [CrossRef]
- Capitani, G.; Duarte, J.M.; Baskaran, K.; Bliven, S.; Somody, J.C. Understanding the Fabric of Protein Crystals: Computational Classification of Biological Interfaces and Crystal Contacts. Bioinformatics 2015, 32, 481–489. [Google Scholar] [CrossRef]
- Dey, S.; Ritchie, D.W.; Levy, E.D. PDB-Wide Identification of Biological Assemblies from Conserved Quaternary Structure Geometry. Nat. Methods 2018, 15, 67–72. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Ponstingl, H.; Kabir, T.; Thornton, J.M. Automatic Inference of Protein Quaternary Structure from Crystals. J. Appl. Crystallogr. 2003, 36, 1116–1122. [Google Scholar] [CrossRef]
- Bzowska, A.; Kulikowska, E.; Shugar, D. Purine Nucleoside Phosphorylases: Properties, Functions, and Clinical Aspects. Pharmacol. Ther. 2000, 88, 349–425. [Google Scholar] [CrossRef] [PubMed]
- Luić, M.; Štefanić, Z. Can Crystal Symmetry and Packing Influence the Active Site Conformation of Homohexameric Purine Nucleoside Phosphorylases? Croat. Chem. Acta 2016, 89, 197–202. [Google Scholar] [CrossRef]
- Narczyk, M.; Bertoša, B.; Papa, L.; Vuković, V.; Leščić Ašler, I.; Wielgus-Kutrowska, B.; Bzowska, A.; Luić, M.; Štefanić, Z. Helicobacter pylori Purine Nucleoside Phosphorylase Shows New Distribution Patterns of Open and Closed Active Site Conformations and Unusual Biochemical Features. FEBS J. 2018, 285, 1305–1325. [Google Scholar] [CrossRef] [PubMed]
- Pugmire, M.J.; Ealick, S.E. Structural Analyses Reveal Two Distinct Families of Nucleoside Phosphorylases. Biochem. J. 2002, 361, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cheng, T.; Chu, J.; Li, S.; He, B. Efficient Synthesis of Purine Nucleoside Analogs by a New Trimeric Purine Nucleoside Phosphorylase from Aneurinibacillus Migulanus AM007. Molecules 2019, 25, 100. [Google Scholar] [CrossRef]
- Mikleušević, G.; Štefanić, Z.; Narczyk, M.; Wielgus-Kutrowska, B.; Bzowska, A.; Luić, M. Validation of the Catalytic Mechanism of Escherichia Coli Purine Nucleoside Phosphorylase by Structural and Kinetic Studies. Biochimie 2011, 93, 1610–1622. [Google Scholar] [CrossRef]
- Roszczenko-Jasińska, P.; Wojtyś, M.I.; Jagusztyn-Krynicka, E.K. Helicobacter pylori Treatment in the Post-Antibiotics Era—Searching for New Drug Targets. Appl. Microbiol. Biotechnol. 2020, 104, 9891–9905. [Google Scholar] [CrossRef]
- Bubić, A.; Narczyk, M.; Petek, A.; Wojtyś, M.I.; Maksymiuk, W.; Wielgus-Kutrowska, B.; Winiewska-Szajewska, M.; Pavkov-Keller, T.; Bertoša, B.; Štefanić, Z.; et al. The Pursuit of New Alternative Ways to Eradicate Helicobacter pylori Continues: Detailed Characterization of Interactions in the Adenylosuccinate Synthetase Active Site. Int. J. Biol. Macromol. 2023, 226, 37–50. [Google Scholar] [CrossRef]
- Mao, C.; Cook, W.J.; Zhou, M.; Koszalka, G.W.; Krenitsky, T.A.; Ealick, S.E. The Crystal Structure of Escherichia Coli Purine Nucleoside Phosphorylase: A Comparison with the Human Enzyme Reveals a Conserved Topology. Structure 1997, 5, 1373–1383. [Google Scholar] [CrossRef]
- Dyzma, A.; Wielgus-Kutrowska, B.; Girstun, A.; Matošević, Z.J.; Staroń, K.; Bertoša, B.; Trylska, J.; Bzowska, A. Trimeric Architecture Ensures the Stability and Biological Activity of the Calf Purine Nucleoside Phosphorylase: In Silico and In Vitro Studies of Monomeric and Trimeric Forms of the Enzyme. Int. J. Mol. Sci. 2023, 24, 2157. [Google Scholar] [CrossRef]
- Bertoša, B.; Mikleušević, G.; Wielgus-Kutrowska, B.; Narczyk, M.; Hajnić, M.; Leščić Ašler, I.; Tomić, S.; Luić, M.; Bzowska, A. Homooligomerization Is Needed for Stability: A Molecular Modelling and Solution Study of Escherichia Coli Purine Nucleoside Phosphorylase. FEBS J. 2014, 281, 1860–1871. [Google Scholar] [CrossRef]
- Wielgus-Kutrowska, B.; Modrak-Wójcik, A.; Dyzma, A.; Breer, K.; Zolkiewski, M.; Bzowska, A. Purine Nucleoside Phosphorylase Activity Decline Is Linked to the Decay of the Trimeric Form of the Enzyme. Arch. Biochem. Biophys. 2014, 549, 40–48. [Google Scholar] [CrossRef]
- Edgar, R.C.; Batzoglou, S. Multiple Sequence Alignment. Curr. Opin. Struct. Biol. 2006, 16, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E. Enhanced Fold Recognition Using Efficient Short Fragment Clustering. J. Mol. Biochem. 2012, 1, 76–85. [Google Scholar] [PubMed]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely Available Python Tools for Computational Molecular Biology and Bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Bostock, M.; Ogievetsky, V.; Heer, J. D3 Data-Driven Documents. IEEE Trans. Vis. Comput. Graph. 2011, 17, 2301–2309. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y. pyMSAviz: MSA Visualization Python Package for Sequence Analysis [Computer Software]. 2022. Available online: https://github.com/moshi4/pyMSAviz (accessed on 27 December 2023).
- Agirre, J.; Atanasova, M.; Bagdonas, H.; Ballard, C.B.; Baslé, A.; Beilsten-Edmands, J.; Borges, R.J.; Brown, D.G.; Burgos-Mármol, J.J.; Berrisford, J.M.; et al. The CCP 4 Suite: Integrative Software for Macromolecular Crystallography. Acta Crystallogr. D Struct. Biol. 2023, 79, 449–461. [Google Scholar] [CrossRef]
- Waskom, M. Seaborn: Statistical Data Visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Grosse-Kunstleve, R.W.; Sauter, N.K.; Moriarty, N.W.; Adams, P.D. The Computational Crystallography Toolbox: Crystallographic Algorithms in a Reusable Software Framework. J. Appl. Crystallogr. 2002, 35, 126–136. [Google Scholar] [CrossRef]
- Bliven, S.; Lafita, A.; Parker, A.; Capitani, G.; Duarte, J.M. Automated Evaluation of Quaternary Structures from Protein Crystals. PLoS Comput. Biol. 2018, 14, e1006104. [Google Scholar] [CrossRef]
- Schrödinger, LLC. The PyMOL Molecular Graphics System, Version 1.8; Schrödinger, LLC: New York, NY, USA, 2015.
- Xu, Q.; Dunbrack, R.L. Principles and Characteristics of Biological Assemblies in Experimentally Determined Protein Structures. Curr. Opin. Struct. Biol. 2019, 55, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Gomaz, B.; Štefanić, Z. Oligomeric symmetry of purine nucleoside phosphorylases. In Proceedings of the The Twenty-Eighth Croatian-Slovenian Crystallographic Meeting Book of Abstracts, Poreč, Croatia, 7–11 September 2022; Croatian Crystallographic Association: Zagreb, Croatia; p. 35. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symmetry | Hexamers | Trimers | Others |
|---|---|---|---|
| only two-fold (point group 2) | 33/37 (89.2%) | 0/11 (0%) | 0/2 |
| only three-fold (point group 3) | 0/0 | 50/50 (100%) | 0 |
| both (point group 32) | 24/31 (77.4%) | 28/28 (100%) | 0 |
| no two-fold or three-fold symmetry | 0/30 | 0/27 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomaz, B.; Štefanić, Z. Oligomeric Symmetry of Purine Nucleoside Phosphorylases. Symmetry 2024, 16, 124. https://doi.org/10.3390/sym16010124
Gomaz B, Štefanić Z. Oligomeric Symmetry of Purine Nucleoside Phosphorylases. Symmetry. 2024; 16(1):124. https://doi.org/10.3390/sym16010124
Chicago/Turabian StyleGomaz, Boris, and Zoran Štefanić. 2024. "Oligomeric Symmetry of Purine Nucleoside Phosphorylases" Symmetry 16, no. 1: 124. https://doi.org/10.3390/sym16010124
APA StyleGomaz, B., & Štefanić, Z. (2024). Oligomeric Symmetry of Purine Nucleoside Phosphorylases. Symmetry, 16(1), 124. https://doi.org/10.3390/sym16010124