Nanoscopic Approach to Study the Early Stages of Epithelial to Mesenchymal Transition (EMT) of Human Retinal Pigment Epithelial (RPE) Cells In Vitro

 ,
,
Abstract
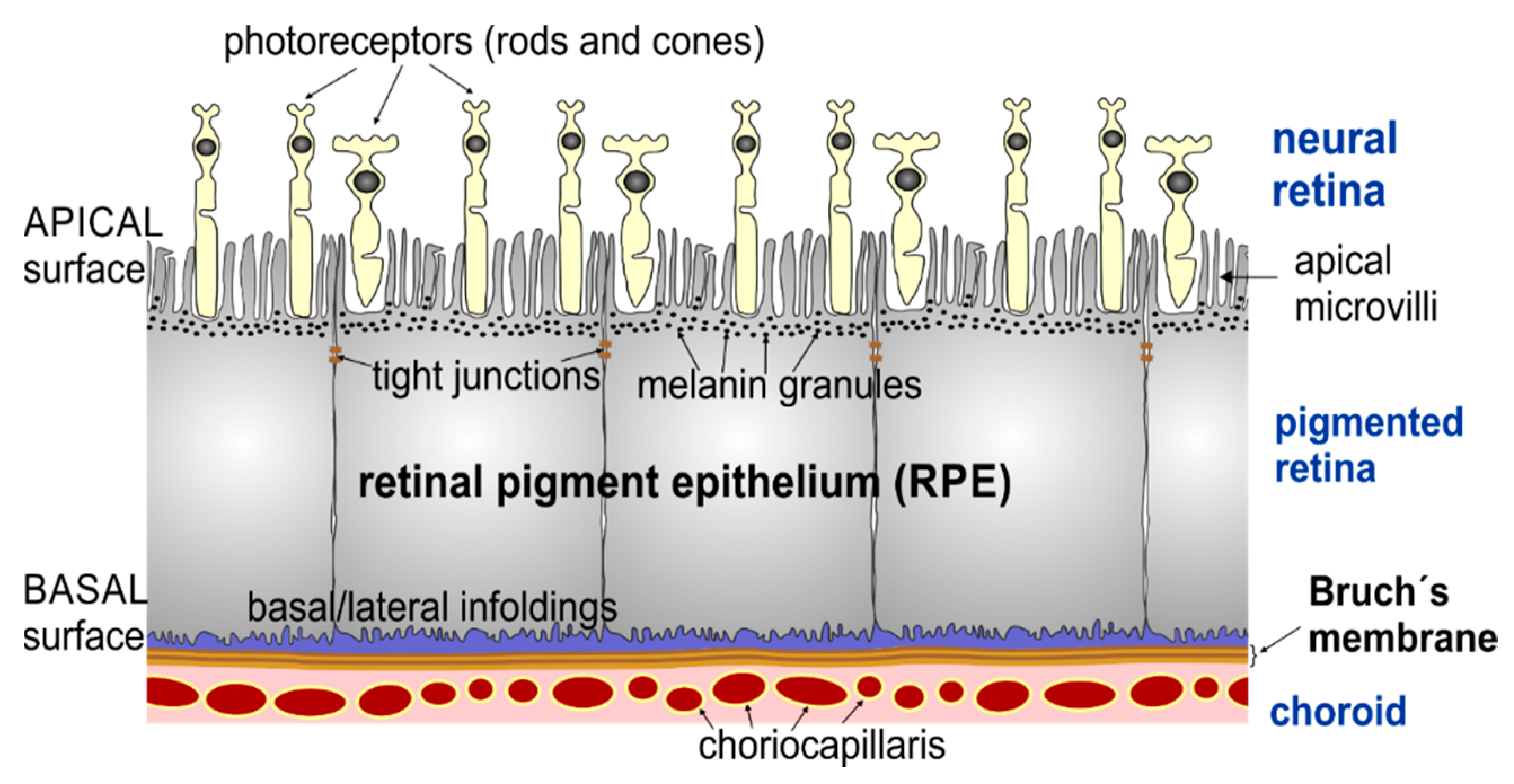
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Primary Human RPE Cells
2.2. Atomic Force Microscopy (AFM)
2.3. Laser-Scanning Confocal Microscopy (LSCM)
2.4. RNA Isolation, cDNA Synthesis, and Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analyses
3. Results
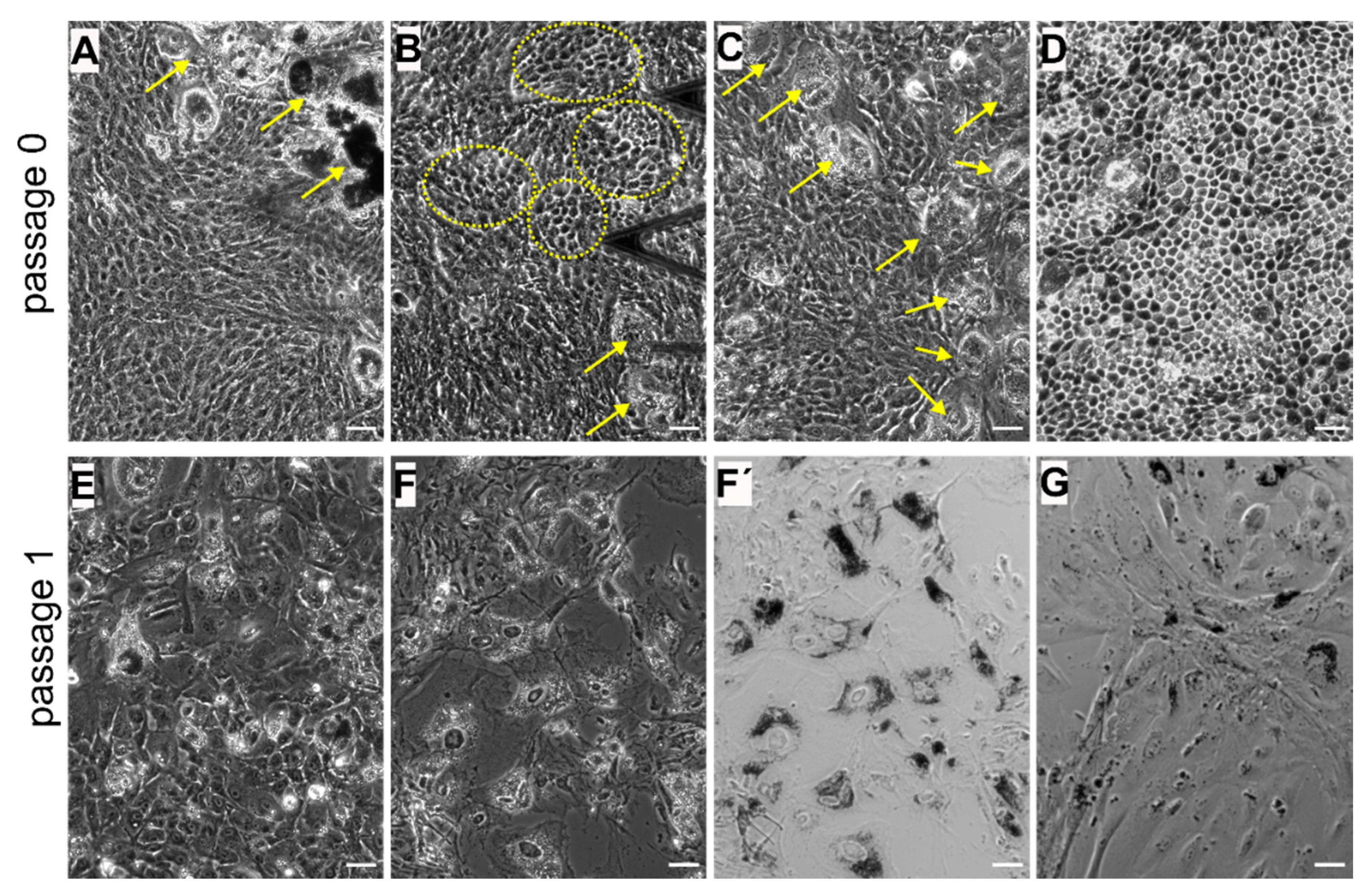
3.1. Diversity of Different RPE Phenotypes at Confluency in Vitro: Assesment with Optical Microscopy
3.2. Cytoskeletal Changes in Cultured hRPE Cells: Correlative AFM and Fluorescence Structural Analysis
3.2.1. Re-Morphogenesis of RPE Cells at Early Stages of EMT: Initial Passages 0 and p1
3.2.2. Cytoskeletal Changes in RPE Cells at Intermediate Stages of EMT: Passages 2 and 4
3.3. Expression Pattern of RPE Differentiation Markers in Cultured hRPE Cells: qRT-PCR Data
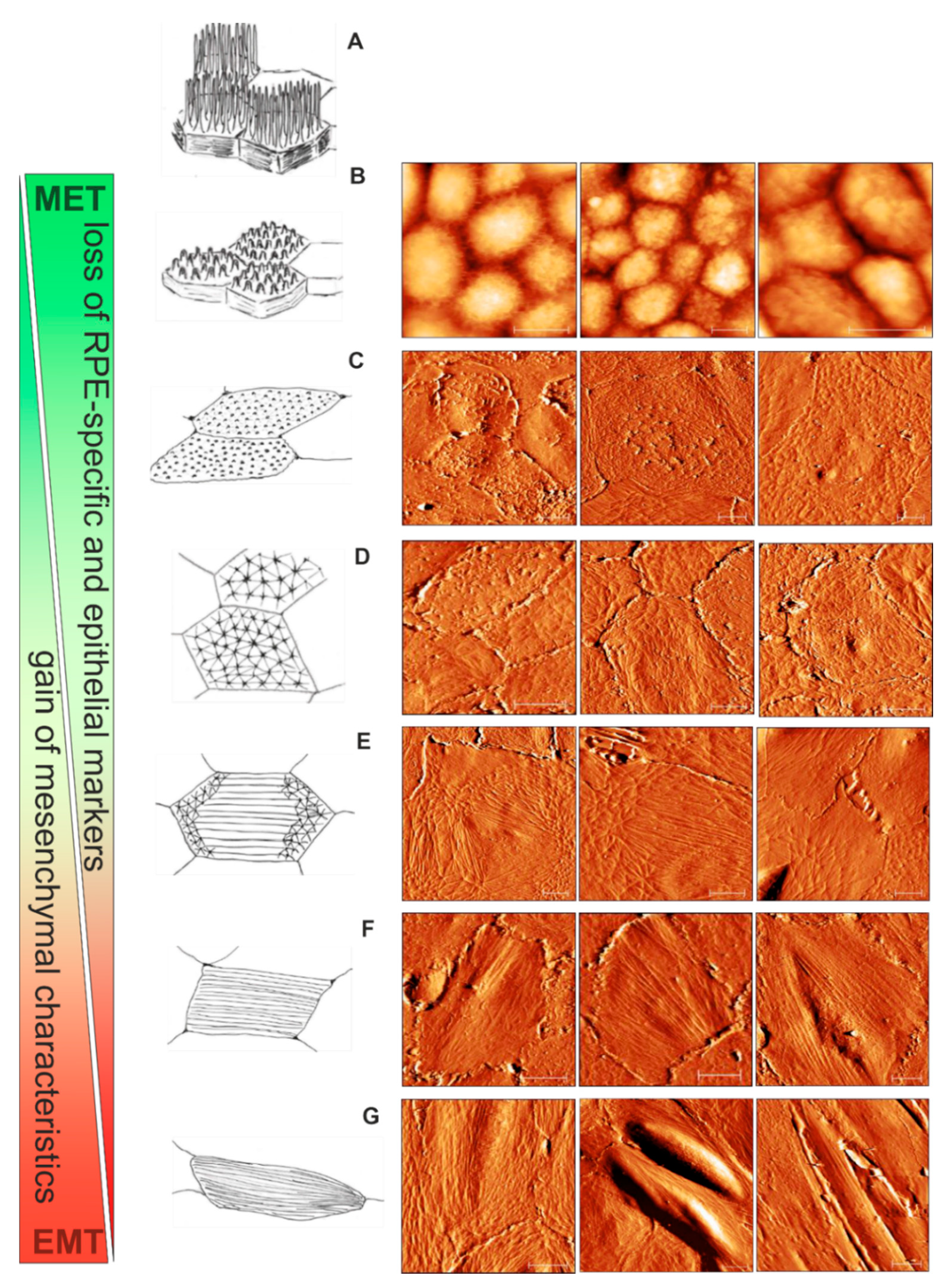
3.4. Scheme for RPE Cytoskeletal Reorganization during EMT: from Cobblestone to Fibroblast-Like Appeareance
4. Discussion
5. Conclusions and Further Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivelä, T.; Jääskeläinen, J.; Vaheri, A.; Carpén, O. Ezrin, a membrane-organizing protein, as a polarization marker of the retinal pigment epithelium in vertebrates. Cell Tissue Res. 2000, 301, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Bonilha, V.L. Retinal pigment epithelium (RPE) cytoskeleton in vivo and in vitro. Exp. Eye Res. 2014, 126, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Bonilha, V.L.; Finnemann, S.C.; Rodriguez-Boulan, E. Ezrin promotes morphogenesis of apical microvilli and basal infoldings in retinal pigment epithelium. J. Cell Biol. 1999, 147, 1533–1548. [Google Scholar] [CrossRef] [Green Version]
- Priglinger, C.S.; Obermann, J.; Szober, C.M.; Merl-Pham, J.; Ohmayer, U.; Behler, J.; Gruhn, F.; Kreutzer, T.C.; Wertheimer, C.; Geerlof, A.; et al. Epithelial-to-Mesenchymal Transition of RPE Cells In Vitro Confers Increased β1,6-N-Glycosylation and Increased Susceptibility to Galectin-3 Binding. PLoS ONE 2016, 11, e0146887. [Google Scholar] [CrossRef] [Green Version]
- Grierson, I.; Hiscott, P.; Hogg, P.; Robey, H.; Mazure, A.; Larkin, G. Development, repair and regeneration of the retinal pigment epithelium. Eye 1994, 8, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, C.; Hiscott, P.; Grierson, I. Retinal Pigment Epithelium Differentiation and Dedifferentiation—Vitreo-Retinal Surgery; Kirchhof, B., Wong, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 101–119. [Google Scholar] [CrossRef]
- Lopez, P.F.; Sippy, B.D.; Lambert, H.M.; Thach, A.B.; Hinton, D.R. Transdifferentiated retinal pigment epithelial cells are immunoreactive for vascular endothelial growth factor in surgically excised age-related macular degeneration-related choroidal neovascular membranes. Investig. Ophthalmol. Vis. Sci. 1996, 37, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Kampik, A.; Kenyon, K.R.; Michels, R.G.; Green, W.R.; de la Cruz, Z.C. Epiretinal and vitreous membranes. Comparative study of 56 cases. Arch. Ophthalmol. 1981, 99, 1445–1454. [Google Scholar] [CrossRef]
- Machemer, R.; Laqua, H. Pigment epithelium proliferation in retinal detachment (massive periretinal proliferation). Am. J. Ophthalmol. 1975, 80, 1–23. [Google Scholar] [CrossRef]
- Vinores, S.A.; Campochiaro, P.A.; McGehee, R.; Orman, W.; Hackett, S.F.; Hjelmeland, L.M. Ultrastructural and immunocytochemical changes in retinal pigment epithelium, retinal glia, and fibroblasts in vitreous culture. Investig. Ophthalmol. Vis. Sci. 1990, 31, 2529–2545. [Google Scholar]
- Hiscott, P.S.; Grierson, I.; McLeod, D. Retinal pigment epithelial cells in epiretinal membranes: An immunohistochemical study. Br. J. Ophthalmol. 1984, 68, 708–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscott, P.; Morino, I.; Alexander, R.; Grierson, I.; Gregor, Z. Cellular components of subretinal membranes in proliferative vitreoretinopathy. Eye 1989, 3, 606–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisanti, S.; Guidry, C. Transdifferentiation of retinal pigment epithelial cells from epithelial to mesenchymal phenotype. Investig. Ophthalmol. Vis. Sci. 1995, 36, 391–405. [Google Scholar]
- Hiscott, P.; Sheridan, C.; Magee, R.M.; Grierson, I. Matrix and the retinal pigment epithelium in proliferative retinal disease. Prog. Retin. Eye Res. 1999, 18, 167–190. [Google Scholar] [CrossRef]
- Ghosh, S.; Shang, P.; Terasaki, H.; Stepicheva, N.; Hose, S.; Yazdankhah, M.; Weiss, J.; Sakamoto, T.; Bhutto, I.A.; Xia, S.; et al. A Role for βA3/A1-Crystallin in Type 2 EMT of RPE Cells Occurring in Dry Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, AMD104–AMD113. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Martini, B.; Pandey, R.; Ogden, T.E.; Ryan, S.J. Cultures of human retinal pigment epithelium. Modulation of extracellular matrix. Investig. Ophthalmol. Vis. Sci. 1992, 33, 516–521. [Google Scholar]
- Rak, D.J.; Hardy, K.M.; Jaffe, G.J.; McKay, B.S. Ca++-switch induction of RPE differentiation. Exp. Eye Res. 2006, 82, 648–656. [Google Scholar] [CrossRef]
- Kimura, K.; Orita, T.; Liu, Y.; Yang, Y.; Tokuda, K.; Kurakazu, T.; Noda, T.; Yanai, R.; Morishige, N.; Takeda, A.; et al. Attenuation of EMT in RPE cells and subretinal fibrosis by an RAR-γ agonist. J. Mol. Med. 2015, 93, 749–758. [Google Scholar] [CrossRef]
- Nassar, K.; Grisanti, S.; Tura, A.; Lüke, J.; Lüke, M.; Soliman, M.; Grisanti, S. A TGF-β receptor 1 inhibitor for prevention of proliferative vitreoretinopathy. Exp. Eye Res. 2014, 123, 72–86. [Google Scholar] [CrossRef]
- Sundaram, V.; Barsam, A.; Virgili, G. Intravitreal low molecular weight heparin and 5-Fluorouracil for the prevention of proliferative vitreoretinopathy following retinal reattachment surgery. Cochrane Database Syst. Rev. 2013, 1, CD006421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedemann, P.; Hilgers, R.D.; Bauer, P.; Heimann, K. Adjunctive daunorubicin in the treatment of proliferative vitreoretinopathy: Results of a multicenter clinical trial. Daunomycin Study Group. Am. J. Ophthalmol. 1998, 126, 550–559. [Google Scholar] [CrossRef]
- Sanabria Ruiz-Colmenares, M.R.; Pastor Jimeno, J.C.; Garrote Adrados, J.A.; Telleria Orriols, J.J.; Yugueros Fernández, M.I. Cytokine gene polymorphisms in retinal detachment patients with and without proliferative vitreoretinopathy: A preliminary study. Acta Ophthalmol. Scand. 2006, 84, 309–313. [Google Scholar] [CrossRef]
- Pastor-Idoate, S.; Rodríguez-Hernández, I.; Rojas, J.; Fernández, I.; García-Gutiérrez, M.T.; Ruiz-Moreno, J.M.; Rocha-Sousa, A.; Ramkissoon, Y.; Harsum, S.; MacLaren, R.E.; et al. The T309G MDM2 Gene Polymorphism Is a Novel Risk Factor for Proliferative Vitreoretinopathy. PLoS ONE 2013, 8, e82283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, S.; Muether, P.S.; Caramoy, A.; Hahn, M.; Abdel-Salam, M.; Diestelhorst, M.; Kirchhof, B.; Fauser, S. Anterior chamber aqueous flare is a strong predictor for proliferative vitreoretinopathy in patients with rhegmatogenous retinal detachment. Retina 2012, 32, 38–42. [Google Scholar] [CrossRef]
- Kuo, H.K.; Chen, Y.H.; Wu, P.C.; Wu, Y.C.; Huang, F.; Kuo, C.W.; Lo, L.H.; Shiea, J. Attenuated glial reaction in experimental proliferative vitreoretinopathy treated with liposomal doxorubicin. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3167–3174. [Google Scholar] [CrossRef] [Green Version]
- Machemer, R. Proliferative vitreoretinopathy (PVR): A personal account of its pathogenesis and treatment. Proctor lecture. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1771–1783. [Google Scholar]
- Pastor, J.C.; De La Rúa, E.R.; Martín, F. Proliferative vitreoretinopathy: Risk factors and pathobiology. Prog. Retin. Eye Res. 2002, 21, 127–144. [Google Scholar] [CrossRef]
- Binnig, G.; Quate, C.F.; Gerber, C. Atomic force microscope. Phys. Rev. Lett. 1986, 56, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Chtcheglova, L.A.; Hinterdorfer, P. Simultaneous AFM topography and recognition imaging at the plasma membrane of mammalian cells. Semin. Cell Dev. Biol. 2018, 73, 45–56. [Google Scholar] [CrossRef]
- Dufrêne, Y.F.; Ando, T.; Garcia, R.; Alsteens, D.; Martinez-Martin, D.; Engel, A.; Gerber, C.; Müller, D.J. Imaging modes of atomic force microscopy for application in molecular and cell biology. Nat. Nanotechnol. 2017, 12, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.J.; Dufrêne, Y.F. Atomic force microscopy: A nanoscopic window on the cell surface. Trends Cell Biol. 2011, 21, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.; Meder, D.; Simons, K.; Müller, D. The effect of raft lipid depletion on microvilli formation in MDCK cells, visualized by atomic force microscopy. FEBS Lett. 2004, 565, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Henderson, E.; Haydon, P.G.; Sakaguchi, D.S. Actin filament dynamics in living glial cells imaged by atomic force microscopy. Science 1992, 257, 1944–1946. [Google Scholar] [CrossRef] [PubMed]
- Braet, F.; Seynaeve, C.; De Zanger, R.; Wisse, E. Imaging surface and submembranous structures with the atomic force microscope: A study on living cancer cells, fibroblasts and macrophages. J. Microsc. 1998, 190, 328–338. [Google Scholar] [CrossRef]
- Last, J.A.; Russell, P.; Nealey, P.F.; Murphy, C.J. The applications of atomic force microscopy to vision science. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6083–6094. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, D.; Lian, R.; Han, Y.; Guo, Y.; Li, Z.; Tang, S.; Chen, J. Matrigel and Activin A promote cell-cell contact and anti-apoptotic activity in cultured human retinal pigment epithelium cells. Exp. Eye Res. 2016, 147, 37–49. [Google Scholar] [CrossRef]
- Burke, J.M.; Skumatz, C.M.; Irving, P.E.; McKay, B.S. Phenotypic heterogeneity of retinal pigment epithelial cells in vitro and in situ. Exp. Eye Res. 1996, 62, 63–73. [Google Scholar] [CrossRef]
- Alge, C.S.; Suppmann, S.; Priglinger, S.G.; Neubauer, A.S.; May, C.A.; Hauck, S.; Welge-Lussen, U.; Ueffing, M.; Kampik, A. Comparative Proteome Analysis of Native Differentiated and Cultured Dedifferentiated Human RPE Cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3629–3641. [Google Scholar] [CrossRef] [Green Version]
- Chtcheglova, L.A.; Waschke, J.; Wildling, L.; Drenckhahn, D.; Hinterdorfer, P. Nano-scale dynamic recognition imaging on vascular endothelial cells. Biophys. J. 2007, 93, L11–L13. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.; Liu, J.; Dubra, A.; Fariss, R. In Vivo Imaging of the Human Retinal Pigment Epithelial Mosaic Using Adaptive Optics Enhanced Indocyanine Green Ophthalmoscopy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4376–4384. [Google Scholar] [CrossRef] [PubMed]
- McKay, B.S.; Burke, J.M. Separation of Phenotypically Distinct Subpopulations of Cultured Human Retinal Pigment Epithelial Cells. Exp. Cell Res. 1994, 213, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Tarau, I.S.; Berlin, A.; Curcio, C.A.; Ach, T. The Cytoskeleton of the Retinal Pigment Epithelium: From Normal Aging to Age-Related Macular Degeneration. Int. J. Mol. Sci. 2019, 20, 3578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarides, E.; Revel, J.P. The molecular basis of cell movement. Sci. Am. 1979, 240, 100–113. [Google Scholar] [CrossRef]
- Girasole, M.; Pompeo, G.; Cricenti, A.; Congiu-Castellano, A.; Andreola, F.; Serafino, A.; Frazer, B.H.; Boumis, G.; Amiconi, G. Roughness of the plasma membrane as an independent morphological parameter to study RBCs: A quantitative atomic force microscopy investigation. Biochim. Biophys. Acta Biomembr. 2007, 1768, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Hecht, E.; Usmani, S.M.; Albrecht, S.; Wittekindt, O.H.; Dietl, P.; Mizaikoff, B.; Kranz, C. Atomic force microscopy of microvillous cell surface dynamics at fixed and living alveolar type II cells. Anal. Bioanal. Chem. 2011, 399, 2369–2378. [Google Scholar] [CrossRef]
- D Antonio, P.; Lasalvia, M.; Perna, G.; Capozzi, V. Scale-independent roughness value of cell membranes studied by means of AFM technique. Biochim. Biophys. Acta Biomembr. 2012, 1818, 3141–3148. [Google Scholar] [CrossRef] [Green Version]
- Ts’o, M.O.M.; Friedman, E. The Retinal Pigment Epithelium: I. Comparative Histology. JAMA Ophthalmol. 1967, 78, 641–649. [Google Scholar] [CrossRef]
- Ach, T.; Huisingh, C.; McGwin, G., Jr.; Messinger, J.D.; Zhang, T.; Bentley, M.J.; Gutierrez, D.B.; Ablonczy, Z.; Smith, R.T.; Sloan, K.R.; et al. Quantitative autofluorescence and cell density maps of the human retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4832–4841. [Google Scholar] [CrossRef] [Green Version]
- Del Priore, L.V.; Kuo, Y.H.; Tezel, T.H. Age-Related Changes in Human RPE Cell Density and Apoptosis Proportion In Situ. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3312–3318. [Google Scholar]
- Gao, H.; Hollyfield, J.G. Aging of the human retina. Differential loss of neurons and retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1–17. [Google Scholar]
- Bhatia, S.K.; Rashid, A.; Chrenek, M.A.; Zhang, Q.; Bruce, B.B.; Klein, M.; Boatright, J.H.; Jiang, Y.; Grossniklaus, H.E.; Nickerson, J.M. Analysis of RPE morphometry in human eyes. Mol. Vis. 2016, 22, 898–916. [Google Scholar] [PubMed]
- Guidry, C.; Medeiros, N.E.; Curcio, C.A. Phenotypic Variation of Retinal Pigment Epithelium in Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2002, 43, 267–273. [Google Scholar]
- Ach, T.; Tolstik, E.; Messinger, J.D.; Zarubina, A.V.; Heintzmann, R.; Curcio, C.A. Lipofuscin redistribution and loss accompanied by cytoskeletal stress in retinal pigment epithelium of eyes with age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3242–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, K.; He, S.; Terasaki, H.; Nazari, H.; Zhang, H.; Spee, C.; Kannan, R.; Hinton, D.R. Resveratrol inhibits epithelial-mesenchymal transition of retinal pigment epithelium and development of proliferative vitreoretinopathy. Sci. Rep. 2015, 5, 16386. [Google Scholar] [CrossRef]
- Rotsch, C.; Radmacher, M. Drug-induced changes of cytoskeletal structure and mechanics in fibroblasts: An atomic force microscopy study. Biophys. J. 2000, 78, 520–535. [Google Scholar] [CrossRef] [Green Version]
- Pillet, F.; Chopinet, L.; Formosa, C.; Dague, É. Atomic Force Microscopy and pharmacology: From microbiology to cancerology. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 1028–1050. [Google Scholar] [CrossRef]
- Chtcheglova, L.A.; Atalar, F.; Ozbek, U.; Wildling, L.; Ebner, A.; Hinterdorfer, P. Localization of the ergtoxin-1 receptors on the voltage sensing domain of hERG K+ channel by AFM recognition imaging. Pflug. Arch. Eur. J. Physiol. 2008, 456, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Opas, M. The Transmission of Forces Between Cells and Their Environment BT—Cytomechanics: The Mechanical Basis of Cell Form and Structure; Bereiter-Hahn, J., Anderson, O.R., Reif, W.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 273–285. [Google Scholar] [CrossRef]
- Lazarides, E. Immunofluorescence studies on the structure of actin filaments in tissue culture cells. J. Histochem. Cytochem. 1975, 23, 507–528. [Google Scholar] [CrossRef]
- Lazarides, E. Actin, alpha-actinin, and tropomyosin interaction in the structural organization of actin filaments in nonmuscle cells. J. Cell Biol. 1976, 68, 202–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarides, E.; Burridge, K. α-Actinin: Immunofluorescent localization of a muscle structural protein in nonmuscle cells. Cell 1975, 6, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Tucker, R.W.; Sanford, K.K.; Frankel, F.R. Tubulin and actin in paired nonneoplastic and spontaneously transformed neoplastic cell lines in vitro: Fluorescent antibody studies. Cell 1978, 13, 629–642. [Google Scholar] [CrossRef]
- Gordon, W.E.; Bushnell, A. Immunofluorescent and ultrastructural studies of polygonal microfilament networks in respreading non-muscle cells. Exp. Cell Res. 1979, 120, 335–348. [Google Scholar] [CrossRef]
- Ireland, G.W.; Voon, F.C. Polygonal networks in living chick embryonic cells. J. Cell Sci. 1981, 52, 55–69. [Google Scholar]
- Soranno, T.; Bell, E. Cytostructural dynamics of spreading and translocating cells. J. Cell Biol. 1982, 95, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Ireland, G.W.; Sanders, E.J.; Voon, F.C.T.; Wakely, J. The ultrastructure of polygonal networks in chick embryonic cells in vitro. Cell Biol. Int. Rep. 1983, 7, 679–688. [Google Scholar] [CrossRef]
- Mochizuki, Y.; Furukawa, K. Application of Coomassie brilliant blue staining to cultured hepatocytes. Cell Biol. Int. Rep. 1987, 11, 367–371. [Google Scholar] [CrossRef]
- Mochizuki, Y.; Furukawa, K.; Mitaka, T.; Yokoi, T.; Kodama, T. Polygonal networks, “geodomes”, of adult rat hepatocytes in primary culture. Cell Biol. Int. Rep. 1988, 12, 1–7. [Google Scholar] [CrossRef]
- Lin, Z.; Holtzer, S.; Schultheiss, T.; Murray, J.; Masaki, T.; Fischman, D.A.; Holtzer, H. Polygons and adhesion plaques and the disassembly and assembly of myofibrils in cardiac myocytes. J. Cell Biol. 1989, 108, 2355–2367. [Google Scholar] [CrossRef]
- Clark, A.F.; Wilson, K.; McCartney, M.D.; Miggans, S.T.; Kunkle, M.; Howe, W. Glucocorticoid-induced formation of cross-linked actin networks in cultured human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 1994, 35, 281–294. [Google Scholar]
- Entcheva, E.; Bien, H. Mechanical and spatial determinants of cytoskeletal geodesic dome formation in cardiac fibroblasts. Integr. Biol. 2009, 1, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.; Scholz, D.L.; Liou, W.; Rafferty, N.S. Polygonal arrays of actin filaments in human lens epithelial cells. An aging study. Investig. Ophthalmol. Vis. Sci. 1986, 27, 1535–1540. [Google Scholar]
- Rafferty, N.S.; Scholz, D.L. Comparative study of actin filament patterns in lens epithelial cells. Are these determined by the mechanisms of lens accommodation? Curr. Eye Res. 1989, 8, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Boal, D.H.; Boey, S.K. Simulations of the erythrocyte cytoskeleton at large deformation. II. Micropipette aspiration. Biophys. J. 1998, 75, 1584–1597. [Google Scholar] [CrossRef] [Green Version]
- Coughlin, M.F.; Stamenović, D. A Prestressed Cable Network Model of the Adherent Cell Cytoskeleton. Biophys. J. 2003, 84, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Vera, C.; Skelton, R.; Bossens, F.; Sung, L.A. 3-D Nanomechanics of an Erythrocyte Junctional Complex in Equibiaxial and Anisotropic Deformations. Ann. Biomed. Eng. 2005, 33, 1387–1404. [Google Scholar] [CrossRef]
- Ingber, D.E. Cellular tensegrity: Defining new rules of biological design that govern the cytoskeleton. J. Cell Sci. 1993, 104, 613–627. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (Forward and Reverse) | Product Size |
|---|---|---|---|
| ACTA2 | NM_001141945.2 | 5′-CTGAAGTACCCGATAGAACATGG-3′ 5′-TTGTAGAAAGAGTGGTGCCAGAT-3′ | 77 bp |
| CDH1 | NM_004360.4 | 5′-CCCGGGACAACGTTTATTAC-3′ 5′-GCTGGCTCAAGTCAAAGTCC-3′ | 71 bp |
| RPE65 | NM_000329.2 | 5′-CCCTCCTGCACAAGTTTGAC-3′ 5′-TCAGTCATTGCCCGTACGTA-3′ | 91 bp |
| GNB2L | NM_006098 | 5′-CTACAATGATCTTTCCCTCTAAATCC-3′ 5′-CCTAACCGCTACTGGCTGTG-3′ | 72 bp |
| Molecular Markers | Epithelioids p0 | Flat Epithelioids p2 and p3 | Fibroblast-Like Cells p6 |
|---|---|---|---|
| RPE65 | 1.00 ± 0.18 | 0.93 ± 0.23 | 0.00 |
| a-SMA | 1.00 ± 0.04 | 2.94 ± 0.74 | 3.91 ± 0.85 |
| E-cadherin | 1.00 ± 0.34 | 0.68 ± 0.31 | 0.02 ± 0.02 |
| Cell Phenotype | Membrane Characteristics | Rrms, nm | SD, nm | Number of Cells |
|---|---|---|---|---|
| Cuboidal cells in monolayer | microvillous structure | 333.0 | 95.3 | 6 |
| Cuboidal cells in clusters | microvillous structure | |||
| cluster 1 | 245.8 | 41.5 | 7 | |
| cluster 2 | 266.0 | 99.1 | 4 | |
| Flat epithelioids | ruffles and rod-like structures | 85.0 | 30.9 | 7 |
| Flat epithelioids | geodomes | 43.5 | 6.2 | 9 |
| Fibroblast-like cells | stress fibers | 50.2 | 21.2 | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chtcheglova, L.A.; Ohlmann, A.; Boytsov, D.; Hinterdorfer, P.; Priglinger, S.G.; Priglinger, C.S. Nanoscopic Approach to Study the Early Stages of Epithelial to Mesenchymal Transition (EMT) of Human Retinal Pigment Epithelial (RPE) Cells In Vitro. Life 2020, 10, 128. https://doi.org/10.3390/life10080128
Chtcheglova LA, Ohlmann A, Boytsov D, Hinterdorfer P, Priglinger SG, Priglinger CS. Nanoscopic Approach to Study the Early Stages of Epithelial to Mesenchymal Transition (EMT) of Human Retinal Pigment Epithelial (RPE) Cells In Vitro. Life. 2020; 10(8):128. https://doi.org/10.3390/life10080128
Chicago/Turabian StyleChtcheglova, Lilia A., Andreas Ohlmann, Danila Boytsov, Peter Hinterdorfer, Siegfried G. Priglinger, and Claudia S. Priglinger. 2020. "Nanoscopic Approach to Study the Early Stages of Epithelial to Mesenchymal Transition (EMT) of Human Retinal Pigment Epithelial (RPE) Cells In Vitro" Life 10, no. 8: 128. https://doi.org/10.3390/life10080128
APA StyleChtcheglova, L. A., Ohlmann, A., Boytsov, D., Hinterdorfer, P., Priglinger, S. G., & Priglinger, C. S. (2020). Nanoscopic Approach to Study the Early Stages of Epithelial to Mesenchymal Transition (EMT) of Human Retinal Pigment Epithelial (RPE) Cells In Vitro. Life, 10(8), 128. https://doi.org/10.3390/life10080128