Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress


 ,
,  , ,
, ,
Abstract
:1. Introduction
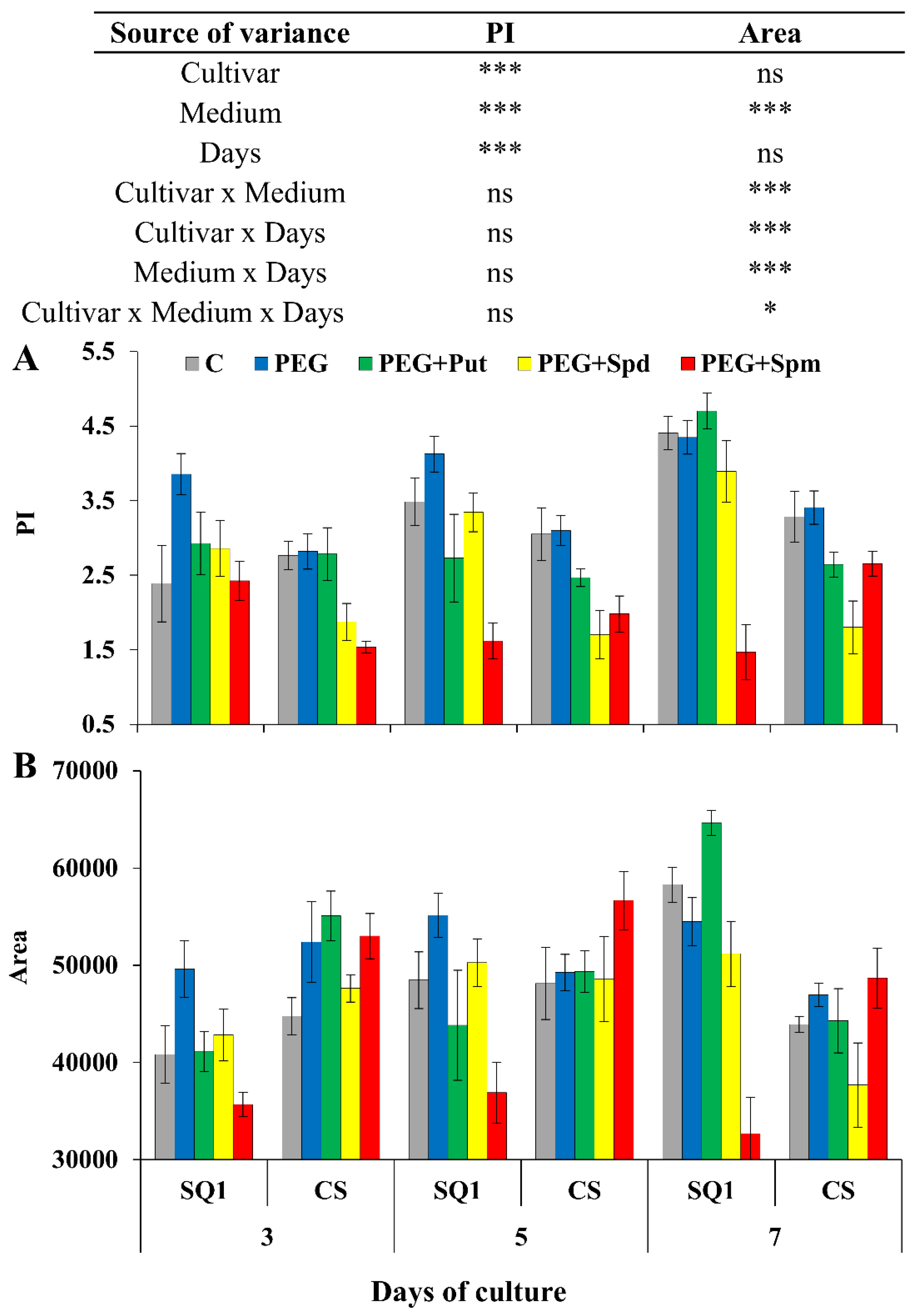
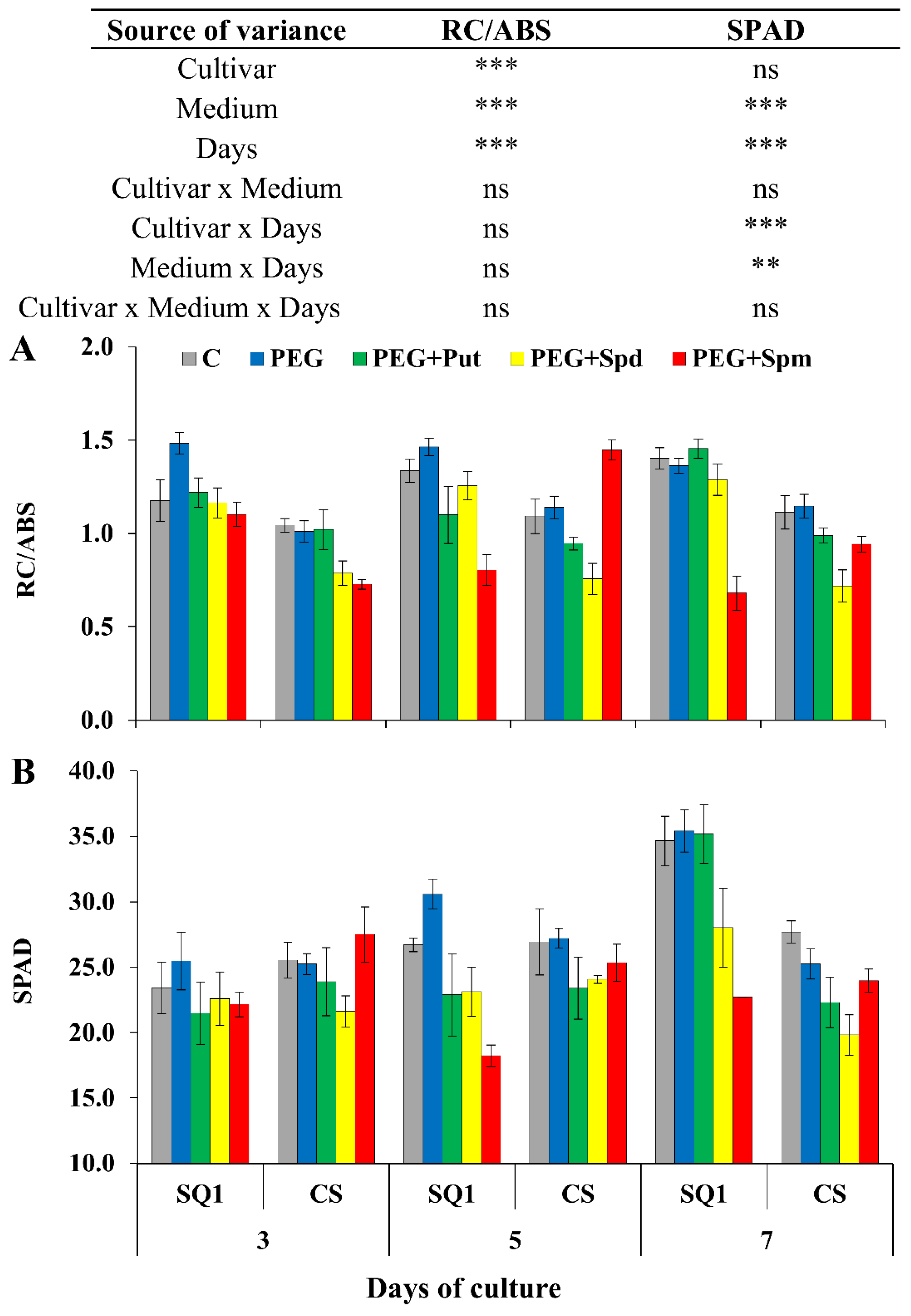
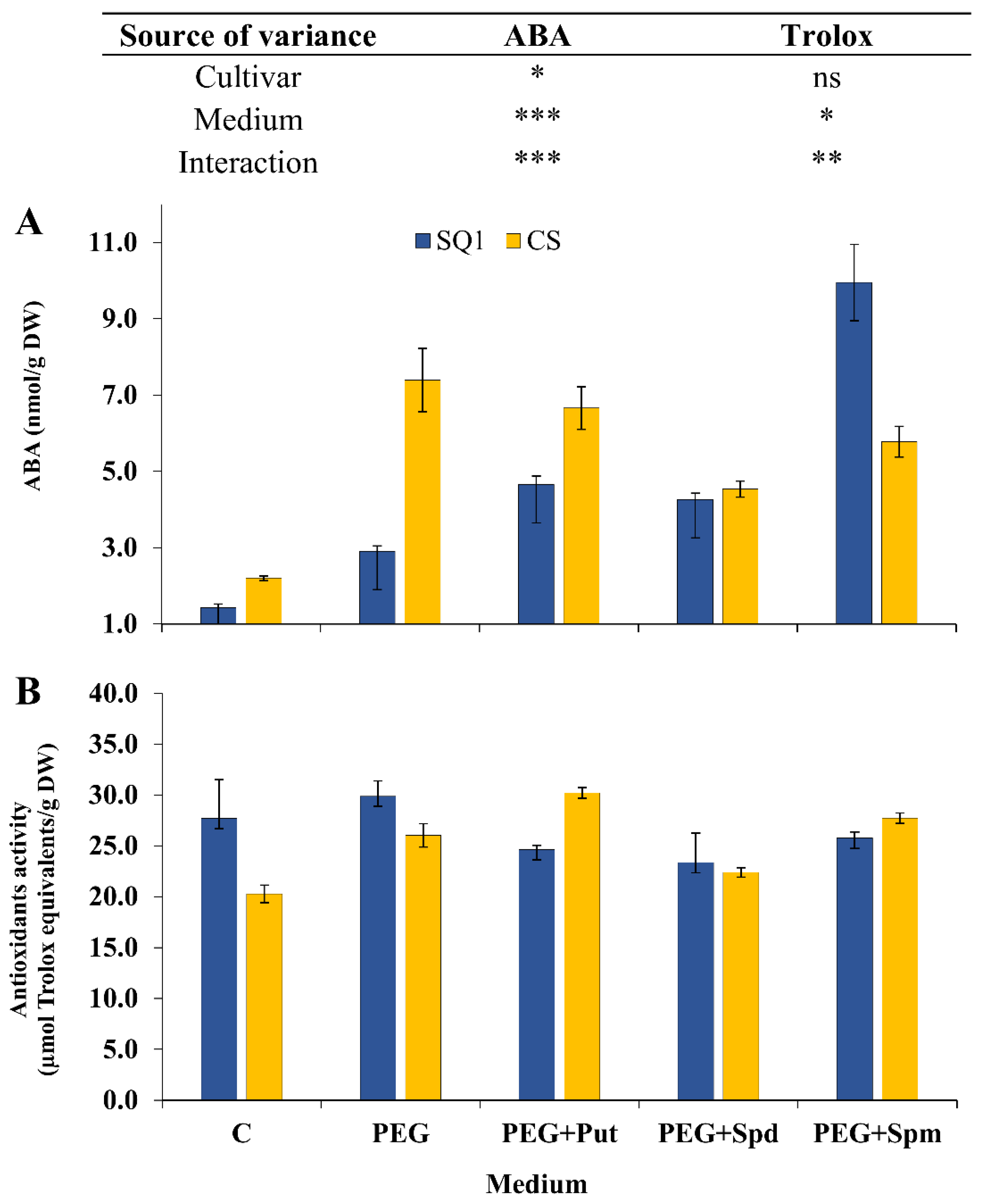
2. Results
2.1. Physiological Parameters
2.2. Biochemical Parameters
2.3. Water Content and Yield Components
3. Discussion
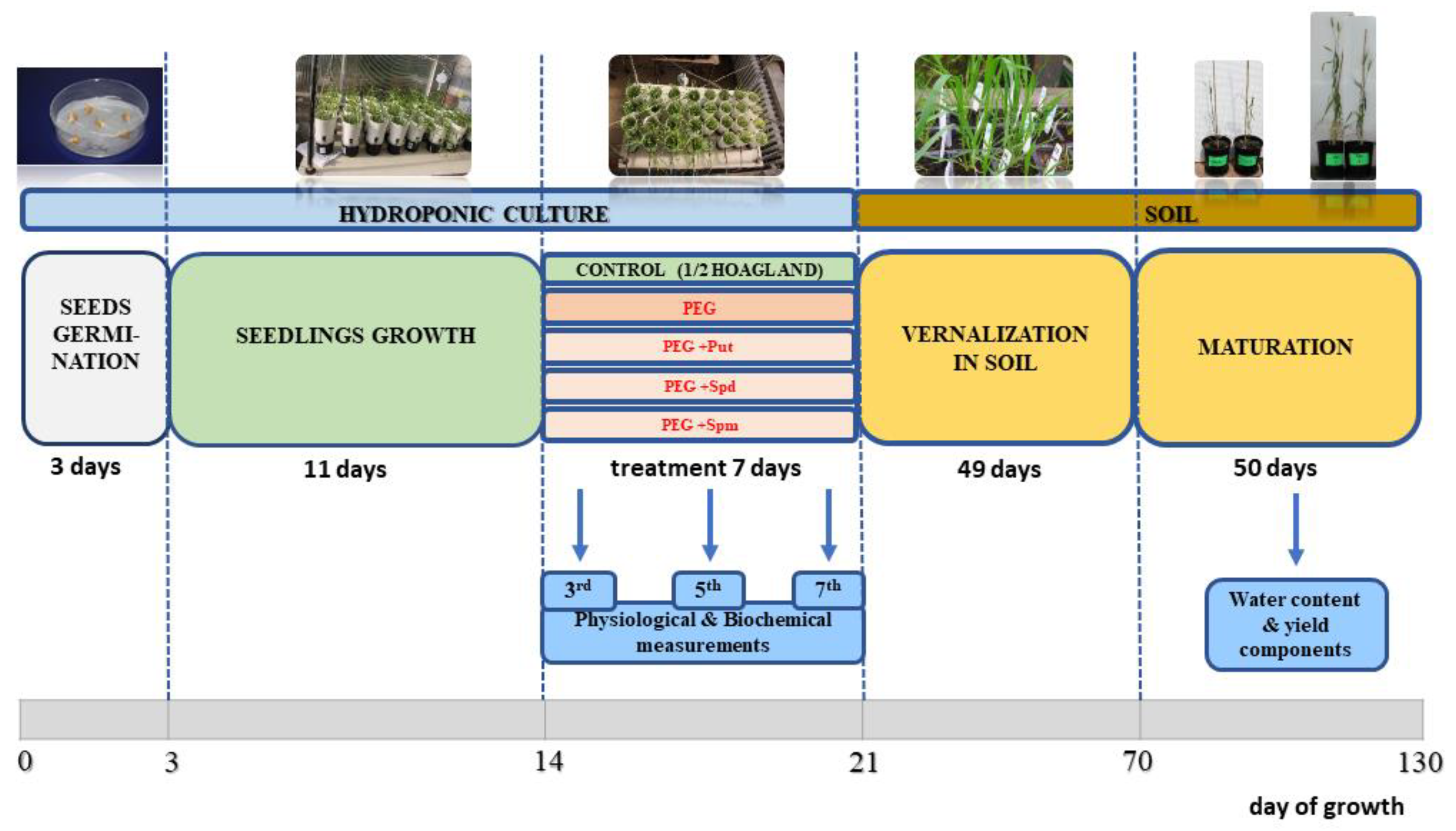
4. Materials and Methods
4.1. Plant Material
4.2. Growth Conditions
4.3. Physiological Parameters
4.4. Biochemical Parameters
4.4.1. Soluble Carbohydrates
4.4.2. Total Phenolics
4.4.3. Proline
4.4.4. Salicylic Acid
4.4.5. Abscisic Acid
4.4.6. Total Low Molecular Weight Antioxidant Activity
4.5. Water Content and Yield Components
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| Area | area over the chlorophyll a fluorescence induction curve |
| C | control |
| CF | chlorophyll a fluorescence |
| CS | Chinese Spring |
| DW | dry weight |
| Fv/Fm | maximal efficiency of PSII photochemistry |
| Fv/Fo | maximum primary yield of PSII photochemistry |
| FW | fresh weight |
| NG | number of grains |
| Pas | polyamines |
| PEG | polyethylene glycol |
| PI | performance index as an essential indicator of sample vitality |
| Put | putrescine |
| RC/ABS | reaction centers per antenna |
| SA | salicylic acid |
| SPAD | chlorophyll content units |
| Spd | spermidine |
| Spm | spermine |
| SQ1 | wheat cultivar |
| WC | water content |
| WG | weight of grains |
References
- Capell, T.; Bassie, L.; Christou, P. Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9909–9914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Nada, K.; Honda, C.; Kitashiba, H.; Wen, X.-P.; Pang, X.-M.; Moriguchi, T. Polyamine biosynthesis of apple callus under salt stress: Importance of the arginine decarboxylase pathway in stress response. J. Exp. Bot. 2006, 57, 2589–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, G.K.; Sabat, S.C. Changes in growth, pigment content and antioxidants in the root and leaf tissues of wheat plants under the influence of exogenous salicylic acid. Braz. J. Plant Physiol. 2011, 23, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining mysteries of molecular biology: The role of polyamines in the cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Gupta, M.; da Silva, E.F.; Svendsen, H.F. Computational study of thermodynamics of polyamines with regard to CO2 capture. Energy Procedia 2012, 23, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Ponappa, T.; Scheerens, J.C.; Miller, A.R. Vacuum infiltration of polyamines increases firmness of strawberry slices under various storage-conditions. J. Food. Sci. 1993, 58, 361–364. [Google Scholar] [CrossRef]
- Cohen, S.S. Introduction to the Polyamines; Prentice-Hall: Englewood Cliffs, NJ, USA, 1971. [Google Scholar]
- Mueller, P.; Schier, A.F. Extracellular movement of signaling molecules. Dev. Cell 2011, 21, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.-I.; Zhu, K.; Sun, Y.-H.; Hegyi, B.; Zeng, Q.; Murphy, C.J.; Small, J.V.; Ye, C.-I.; Izumiya, Y.; Penninger, J.M.; et al. KCNJ15/Kir4.2 couples with polyamines to sense weak extracellular electric fields in galvanotaxis. Nat. Commun. 2015, 6, 8532. [Google Scholar] [CrossRef] [Green Version]
- Moschou, P.N.; Roubelakis-Angelakis, K.A. Polyamines and programmed cell death. J. Exp. Bot. 2014, 65, 1285–1296. [Google Scholar] [CrossRef] [Green Version]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; Roubelakis-Angelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabor, H.; Tabor, C.W. Biosynthesis and metabolism of 1,4-diaminobutane, spermidine, spermine, and related amines. Adv. Enzym. Relat. Areas Mol. Biol. 1972, 36, 203–268. [Google Scholar]
- Maiale, S.; Sanchez, D.H.; Guirado, A.; Vidal, A.; Ruiz, O.A. Spermine accumulation under salt stress. J. Plant Physiol. 2004, 161, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Galston, A.W.; Sawhney, R.K. Polyamines in plant physiology. Plant Physiol. 1990, 94, 406–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navakoudis, E.; Lutz, C.; Langebartels, C.; Lutz-Meindl, U.; Kotzabasis, K. Ozone impact on the photosynthetic apparatus and the protective role of polyamines. BBA Gen. Subj. 2003, 1621, 160–169. [Google Scholar] [CrossRef]
- Dernetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus–The protective role of polyamines. BBA Bioenerg. 2007, 1767, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.H.; Li, J.; Guo, S.R.; Tezuka, T. Effects of exogenous putrescine on gas-exchange characteristics and chlorophyll fluorescence of NaCl-stressed cucumber seedlings. Photosynth. Res. 2009, 100, 155–162. [Google Scholar] [CrossRef]
- Terui, Y.; Yoshida, T.; Sakamoto, A.; Saito, D.; Oshima, T.; Kawazoe, M.; Yokoyama, S.; Igarashi, K.; Kashiwagi, K. Polyamines protect nucleic acids against depurination. Int. J. Biochem. Cell Biol. 2018, 99, 147–153. [Google Scholar] [CrossRef]
- Besford, R.T.; Richardson, C.M.; Campos, J.L.; Tiburcio, A.F. Effect of polyamines on stabilization of molecular-complexes in thylakoid membranes of osmotically stressed oat leaves. Planta 1993, 189, 201–206. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, Z. Effects of endogenous abscisic acid, jasmonic acid, polyamines, and polyamine oxidase activity in tomato seedlings under drought stress. Sci. Hortic. 2013, 159, 172–177. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, H.; Wen, X.-X.; Liao, Y.-C. Effect of polyamine on seed germination of wheat under drought stress is related to changes in hormones and carbohydrates. J. Integr. Agric. 2016, 15, 2759–2774. [Google Scholar] [CrossRef] [Green Version]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirbet, A.; Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photoch. Photobio. B 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Rastogi, A.; Zivcak, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypinski, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Singh, G.P.; Pandey, R.; Ramya, P.; Singh, P.K.; Nivedita; Prabhu, K.V. Chlorophyll fluorescence kinetics and response of wheat (Triticum aestivum L.) under high temperature stress. Indian J. Exp. Biol. 2018, 56, 194–201. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: London, UK, 2000; pp. 445–483. [Google Scholar]
- Singh, R.; Malhotra, S.P. Carbon fixation, sucrose synthesis and its transport to storage tissues. In Carbohydrate Reserves in Plants: Synthesis and Regulation; Gupta, A.K., Kaur, N., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2000; pp. 1–34. [Google Scholar]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Belbachir, O.A.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2017, 218, 26–34. [Google Scholar] [CrossRef]
- Quarrie, S.A. Implications of genetic differences in ABA accumulation for crop production. In Abscisic Acid: Physiology and Biochemistry; Davies, W.J., Jones, H.G., Eds.; BIOS Scientific Publishers: Oxford, UK, 1991; pp. 227–243. [Google Scholar]
- Wang, L.-J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.-J.; Cheng, J.-S.; Luo, H.-B.; Li, S.-H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Kusajima, M.; Okumura, Y.; Fujita, M.; Nakashita, H. Abscisic acid modulates salicylic acid biosynthesis for systemic acquired resistance in tomato. Biosci. Biotechnol. Biochem. 2017, 81, 1850–1853. [Google Scholar] [CrossRef] [Green Version]
- Akashi, K.; Nishimura, N.; Ishida, Y.; Yokota, A. Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon. Biochem. Biophys. Res. Commun. 2004, 323, 72–78. [Google Scholar] [CrossRef]
- Pal, M.; Tajti, J.; Szalai, G.; Peeva, V.; Vegh, B.; Janda, T. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef] [PubMed]
- Bignon, E.; Chan, C.-H.; Morell, C.; Monari, A.; Ravanat, J.-L.; Dumont, E. Molecular dynamics insights into polyamine-DNA binding modes: Implications for cross-link selectivity. Chem. Eur. J. 2017, 23, 12845–12852. [Google Scholar] [CrossRef] [PubMed]
- Bagni, N.; Calzoni, G.L.; Speranza, A. Polyamines as sole nitrogen-sources for helianthus-tuberosus explants invitro. New Phytol. 1978, 80, 317–323. [Google Scholar] [CrossRef]
- Quarrie, S.A.; Steed, A.; Calestani, C.; Semikhodskii, A.; Lebreton, C.; Chinoy, C.; Steele, N.; Pljevljakusic, D.; Waterman, E.; Weyen, J.; et al. A high-density genetic map of hexaploid wheat (Triticum aestivum L.) from the cross Chinese Spring X SQ1 and its use to compare QTLs for grain yield across a range of environments. Appl. Genet. 2005, 110, 865–880. [Google Scholar] [CrossRef]
- Marcinska, I.; Czyczylo-Mysza, I.; Skrzypek, E.; Grzesiak, M.T.; Janowiak, F.; Filek, M.; Dziurka, M.; Dziurka, K.; Waligorski, P.; Juzon, K.; et al. Alleviation of osmotic stress effects by exogenous application of salicylic or abscisic acid on wheat seedlings. Int. J. Mol. Sci. 2013, 14, 13171–13193. [Google Scholar] [CrossRef] [Green Version]
- Marcinska, I.; Czyczylo-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka, M.; Dziurka, K.; et al. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol. Plant 2013, 35, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Ting, S.; Rouseff, R. Proline content in Florida frozen concentrated orange juice and canned grapefruit juice. Proc. Fla. State Hortic. Soc. 1979, 92, 143–145. [Google Scholar]
- Wilbert, S.M.; Ericsson, L.H.; Gordon, M.P. Quantification of jasmonic acid, methyl jasmonate, and salicylic acid in plants by capillary liquid chromatography electrospray tandem mass spectrometry. Anal. Biochem. 1998, 257, 186–194. [Google Scholar] [CrossRef]
- Dziurka, M.; Janeczko, A.; Juhasz, C.; Gullner, G.; Oklestkova, J.; Novak, O.; Saja, D.; Skoczowski, A.; Tobias, I.; Barna, B. Local and systemic hormonal responses in pepper leaves during compatible and incompatible pepper-tobamovirus interactions. Plant Physiol. Biochem. 2016, 109, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Walker-Simmons, M.K.; Abrams, S.R. Use of ABA immunoassays. In Abscisic Acid, Physiology and Biochemistry; Davies, W.J., Jones, H.G., Eds.; Bios Scientific Publishers: Oxford, UK, 1991; pp. 53–63. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Żur, I.; Dubas, E.; Krzewska, M.; Zieliński, K.; Fodor, J.; Janowiak, F. Glutathione provides antioxidative defence and promotes microspore-derived embryo development in isolated microspore cultures of triticale (× Triticosecale Wittm.). Plant Cell Rep. 2019, 38, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Tarchoune, I.; Degl’Innocenti, E.; Kaddour, R.; Guidi, L.; Lachaal, M.; Navari-Izzo, F.; Ouerghi, Z. Effects of NaCl or Na2SO4 salinity on plant growth, ion content and photosynthetic activity in Ocimum basilicum L. Acta Physiol. Plant. 2012, 34, 607–615. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Medium | Water Content (WC) | Number of Grains (NG) | Weight of Grains (WG) | Biomass |
|---|---|---|---|---|---|
| SQ1 | Control | 7.5 ± 0.2 | 82.3 ± 9.9 | 3.5 ± 0.5 | 6.0 ± 0.8 |
| PEG | 4.7 ± 0.4 | 79.6 ± 4.6 | 3.5 ± 0.2 | 6.0 ± 0.4 | |
| PEG + Put | 3.4 ± 0.2 | 60.6 ± 5.3 | 2.3 ± 0.1 | 4.1 ± 0.1 | |
| PEG + Spd | 3.3 ± 0.6 | 82.6 ± 4.7 | 3.4 ± 0.2 | 5.3 ± 0.4 | |
| PEG + Spm | 3.8 ± 0.3 | 89.3 ± 5.4 | 3.5 ± 0.7 | 6.0 ± 1.0 | |
| CS | Control | 5.3 ± 0.3 | 222.6 ± 21.9 | 5.7 ± 0.3 | 13.2 ± 1.0 |
| PEG | 4.2 ± 0.2 | 215.0 ± 5.20 | 5.8 ± 0.5 | 11.6 ± 0.8 | |
| PEG + Put | 3.3 ± 0.2 | 229.3 ± 18.8 | 5.9 ± 0.5 | 10.8 ± 1.4 | |
| PEG + Spd | 3.2 ± 0.3 | 159.0 ± 24.9 | 4.1 ± 0.4 | 8.7 ± 1.0 | |
| PEG + Spm | 2.2 ± 0.4 | 180.6 ± 10.9 | 4.9 ± 0.1 | 10.5 ± 0.0 | |
| Source of Variance | WC | NG | WG | Biomass | |
| Cultivar | ** | *** | *** | *** | |
| Medium | *** | ns | ns | * | |
| Cultivar × Medium | ns | * | * | ns | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcińska, I.; Dziurka, K.; Waligórski, P.; Janowiak, F.; Skrzypek, E.; Warchoł, M.; Juzoń, K.; Kapłoniak, K.; Czyczyło-Mysza, I.M. Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress. Life 2020, 10, 151. https://doi.org/10.3390/life10080151
Marcińska I, Dziurka K, Waligórski P, Janowiak F, Skrzypek E, Warchoł M, Juzoń K, Kapłoniak K, Czyczyło-Mysza IM. Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress. Life. 2020; 10(8):151. https://doi.org/10.3390/life10080151
Chicago/Turabian StyleMarcińska, Izabela, Kinga Dziurka, Piotr Waligórski, Franciszek Janowiak, Edyta Skrzypek, Marzena Warchoł, Katarzyna Juzoń, Kamila Kapłoniak, and Ilona Mieczysława Czyczyło-Mysza. 2020. "Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress" Life 10, no. 8: 151. https://doi.org/10.3390/life10080151
APA StyleMarcińska, I., Dziurka, K., Waligórski, P., Janowiak, F., Skrzypek, E., Warchoł, M., Juzoń, K., Kapłoniak, K., & Czyczyło-Mysza, I. M. (2020). Exogenous Polyamines Only Indirectly Induce Stress Tolerance in Wheat Growing in Hydroponic Culture under Polyethylene Glycol-Induced Osmotic Stress. Life, 10(8), 151. https://doi.org/10.3390/life10080151