
Preliminary Evaluation of Protective Efficacy of Inactivated Senecavirus A on Pigs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Clinical Samples
2.2. RNA Extraction and RNA Detection for Pathogens
2.3. Virus Isolation
2.4. Sequencing and Phylogenetic Analyses
2.5. Pigs Challenge Experiment
2.6. Immunization-Challenge Experiment
3. Results
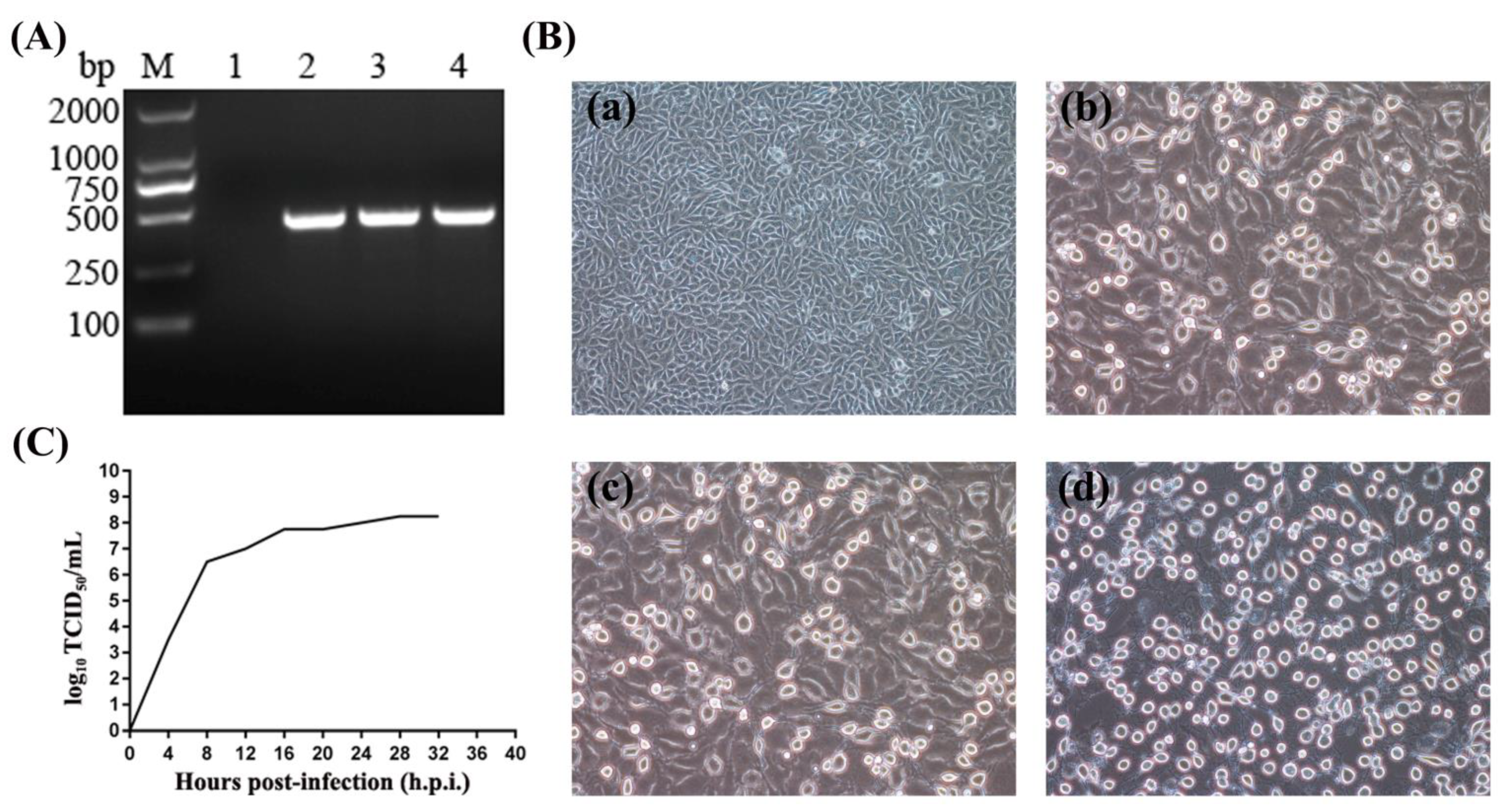
3.1. A SVA Strain Was Isolated from Pig Herd with Vesicular Symptoms
3.2. Characterization of the SVA Genome Sequence
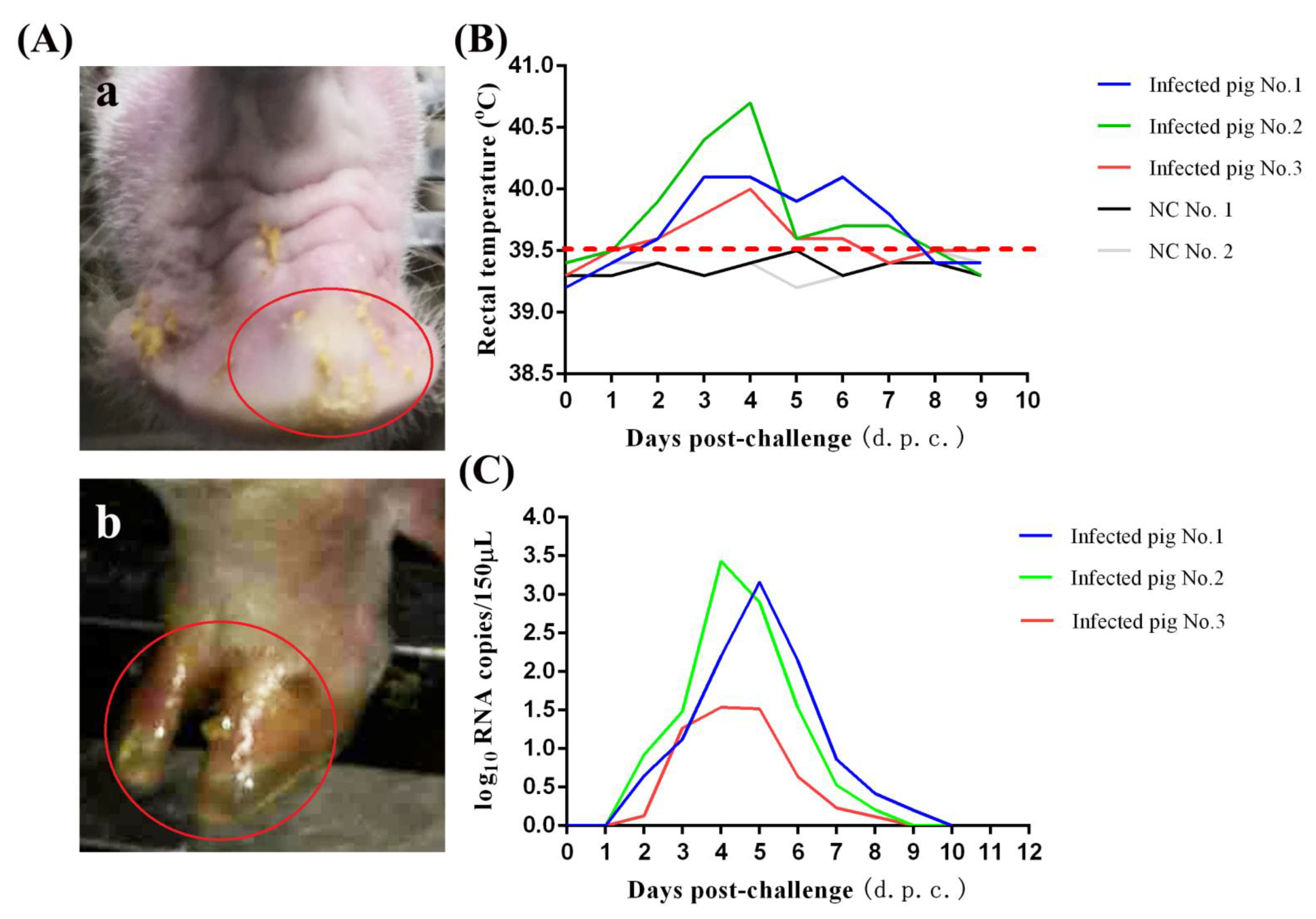
3.3. Pathogenicity of SVA GD-ZYY02-2018 Strain in Pigs
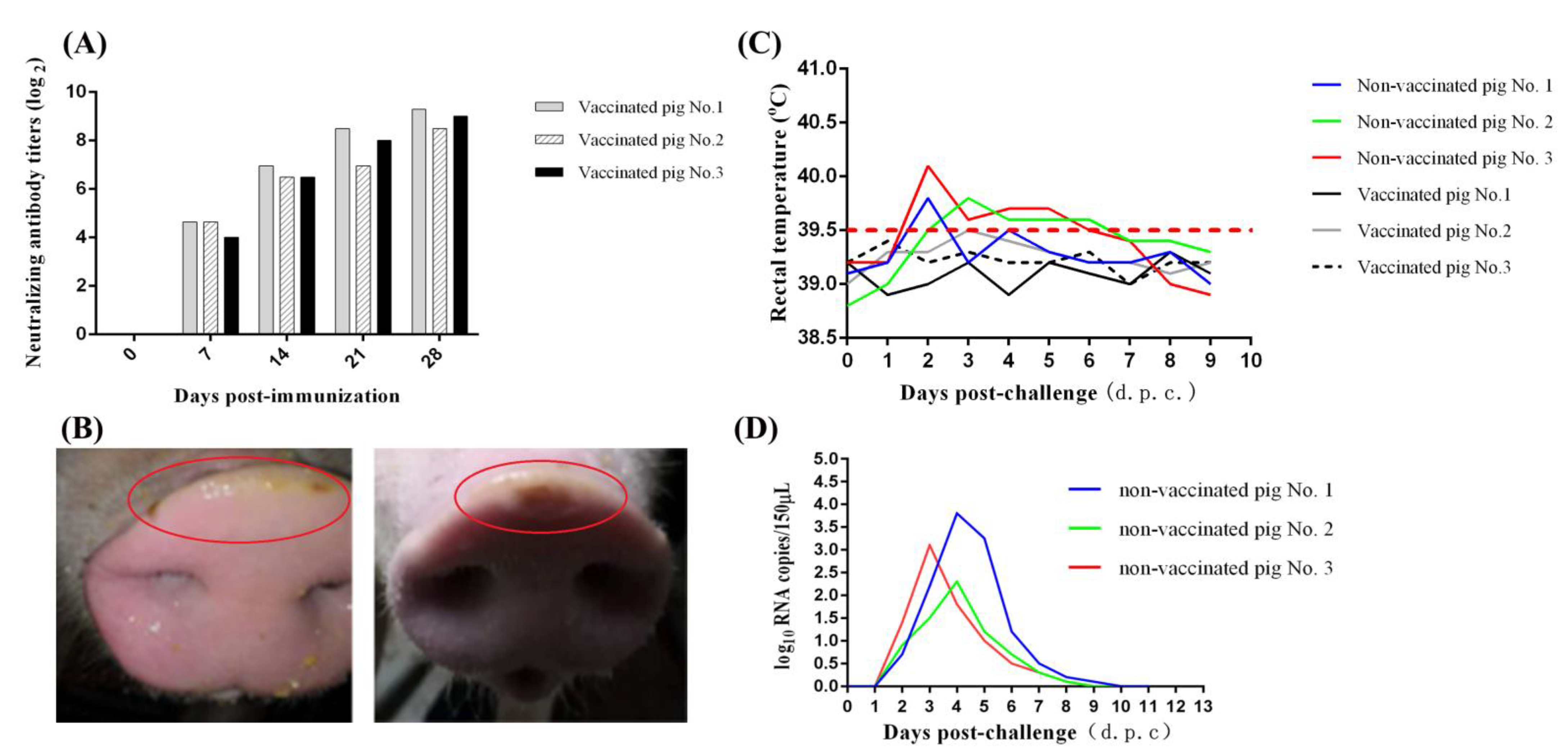
3.4. Protective Efficacy of Inactivated GD-ZYY02-2018 Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L. Complete genome sequence analysis of Seneca Valley virus-001, a novel oncolytic picornavirus. J. Gen. Virol. 2008, 89, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.; Canon, A.; Bates, J.L.; Gerardy, K.; Linhares, D.C.; Pineyro, P.E.; Schwartz, K.J.; Yoon, K.J.; Rademacher, C.J.; Holtkamp, D.; et al. Neonatal Mortality, Vesicular Lesions and Lameness Associated with Senecavirus A in a U.S. Sow Farm. Transbound. Emerg. Dis. 2016, 63, 373–378. [Google Scholar] [CrossRef]
- Pasma, T.; Davidson, S.; Shaw, S.L. Idiopathic vesicular disease in swine in Manitoba. Can. Vet. J. 2008, 49, 84–85. [Google Scholar]
- Vannucci, F.A.; Linhares, D.C.L.; Barcellos, D.E.S.N.; Lam, H.C.; Collins, J.; Marthaler, D. Identification and Complete Genome of Seneca Valley Virus in Vesicular Fluid and Sera of Pigs Affected with Idiopathic Vesicular Disease, Brazil. Transbound. Emerg. Dis. 2015, 62, 589–593. [Google Scholar] [CrossRef]
- Houston, E.; Temeeyasen, G.; Pineyro, P.E. Comprehensive review on immunopathogenesis, diagnostic and epidemiology of Senecavirus A. Virus Res. 2020, 286, 198038. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.L.; Mowrer, C.; Canon, A.; Linhares, D.C.; Rademacher, C.; Karriker, L.A.; Holtkamp, D.J. Systematic Epidemiological Investigations of Cases of Senecavirus A in US Swine Breeding Herds. Transbound. Emerg. Dis. 2017, 64, 11–18. [Google Scholar] [CrossRef]
- Leme, R.A.; Oliveira, T.E.; Alcantara, B.K.; Headley, S.A.; Alfieri, A.F.; Yang, M.; Alfieri, A.A. Clinical Manifestations of Senecavirus A Infection in Neonatal Pigs, Brazil, 2015. Emerg. Infect. Dis. 2016, 22, 1238–1241. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, X.; Bai, Y.; Sun, B.; Xie, Q.; Ma, J. The First Identification and Complete Genome of Senecavirus A Affecting Pig with Idiopathic Vesicular Disease in China. Transbound. Emerg. Dis. 2017, 64, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Fan, W.; Qian, P.; Chen, H.; Li, X. Isolation and full-genome sequencing of Seneca Valley virus in piglets from China, 2016. Virol. J. 2016, 13, 173. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Yang, F.; Chen, P.; Liu, H.; Cao, W.; Zhang, K.; Liu, X.; Zheng, H. Emergence of novel Seneca Valley virus strains in China, 2017. Transbound. Emerg. Dis. 2017, 64, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, J.; Wu, R.T.; Wu, Z.X.; Chen, J.W.; Luo, Y.; Xie, Q.M.; Ma, J.Y. Phylogenetic and Genome Analysis of 17 Novel Senecavirus A Isolates in Guangdong Province, 2017. Front. Vet. Sci. 2018, 5, 314. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Aguero, M.; Romero, L.; Sanchez, C.; Belak, S.; Arias, M.; Sanchez-Vizcaino, J.M. Rapid and differential diagnosis of foot-and-mouth disease, swine vesicular disease, and vesicular stomatitis by a new multiplex RT-PCR assay. J. Virol. Methods 2008, 147, 301–311. [Google Scholar] [CrossRef]
- Lung, O.; Fisher, M.; Beeston, A.; Hughes, K.B.; Clavijo, A.; Goolia, M.; Pasick, J.; Mauro, W.; Deregt, D. Multiplex RT-PCR detection and microarray typing of vesicular disease viruses. J. Virol. Methods 2011, 175, 236–245. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Joshi, L.R.; Mohr, K.A.; Clement, T.; Hain, K.S.; Myers, B.; Yaros, J.; Nelson, E.A.; Christopher-Hennings, J.; Gava, D.; Schaefer, R.; et al. Detection of the Emerging Picornavirus Senecavirus A in Pigs, Mice, and Houseflies. J. Clin. Microbiol. 2016, 54, 1536–1545. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Z.; Zhang, Q.; Sun, M.; Li, S.; Su, W.; Hu, X.; He, W.; Su, J. Efficacy assessment of an inactivated Tembusu virus vaccine candidate in ducks. Res. Vet. Sci. 2017, 110, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhu, Z.; Cao, W.; Liu, H.; Zhang, K.; Tian, H.; Liu, X.; Zheng, H. Immunogenicity and protective efficacy of an inactivated cell culture-derived Seneca Valley virus vaccine in pigs. Vaccine 2018, 36, 841–846. [Google Scholar] [CrossRef]
- Chen, P.; Yang, F.; Cao, W.; Liu, H.; Zhang, K.; Liu, X.; Xu, Z.; Zhu, Z.; Zheng, H. The Distribution of Different Clades of Seneca Valley Viruses in Guangdong Province, China. Virol. Sin. 2018, 33, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, P.; Hao, G.; Liu, W.; Chen, H.; Qian, P.; Li, X. Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China. Pathogens 2020, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yuan, F.; Li, Y.; Shang, P.; Schroeder, R.; Lechtenberg, K.; Henningson, J.; Hause, B.; Bai, J.; Rowland, R.R.R.; et al. Construction and characterization of a full-length cDNA infectious clone of emerging porcine Senecavirus A. Virology 2016, 497, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Fernandes, M.H.V.; de Lima, M.; Joshi, L.R.; Lawson, S.; Diel, D.G. A Novel Live Attenuated Vaccine Candidate Protects against Heterologous Senecavirus A Challenge. Front. Immunol. 2019, 10, 2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; Fan, H.; Zhou, E.; Li, L.; Li, S.; Yan, J.; Jiang, P. Pathogenesis of a senecavirus a isolate from swine in shandong Province, China. Vet. Microbiol. 2020, 242, 108606. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Wang, M.; Pan, S.; Mou, C.; Chen, Z. Pathogenicity of two Chinese Seneca Valley virus (SVV) strains in pigs. Microb. Pathog. 2019, 136, 103695. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) |
|---|---|
| SVA-0-F | TTTGAAATGGGGGGCTGG |
| SVA-0-R | CGAGCCGTGGATATTCAA |
| SVA-1-F | GCACAGAGGAGCAACATC |
| SVA-1-R | GATGTCCAGTCCAAGTTGT |
| SVA-2-F | TAACCGACCTCTTACAACTG |
| SVA-2-R | TGAGACCACCGTGACTTC |
| SVA-3-F | CAGCAGGACGATGGTTAC |
| SVA-3-R | GGAGGCGGTTCTACAGTA |
| SVA-4-F | ACCTGGAGGAAGTATGTGA |
| SVA-4-R | GTCTCTTCTCGGTCTGTATC |
| SVA-5-F | GGCGTTGGGTAGAGTTC |
| SVA-5-R | GAGTAGTCACCGTCTAAGAAT |
| SVA-6-F | GCAAGGACTGATGACTGA |
| SVA-6-R | CACCGTAGGCGATGATAT |
| SVA-7-F | CGCCAAGTTTCAATCCCATC |
| SVA-7-R | TCCAGGTCAGTCGAACAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, Y.; Liao, Y.; Sun, Y.; Ruan, Y.; Liu, C.; Zhang, M.; Li, F.; Li, X.; Fan, S.; et al. Preliminary Evaluation of Protective Efficacy of Inactivated Senecavirus A on Pigs. Life 2021, 11, 157. https://doi.org/10.3390/life11020157
Li Y, Zhang Y, Liao Y, Sun Y, Ruan Y, Liu C, Zhang M, Li F, Li X, Fan S, et al. Preliminary Evaluation of Protective Efficacy of Inactivated Senecavirus A on Pigs. Life. 2021; 11(2):157. https://doi.org/10.3390/life11020157
Chicago/Turabian StyleLi, Yuwan, Yangyi Zhang, Yingxin Liao, Yawei Sun, Yang Ruan, Chenchen Liu, Mengru Zhang, Fangfang Li, Xiaowen Li, Shuangqi Fan, and et al. 2021. "Preliminary Evaluation of Protective Efficacy of Inactivated Senecavirus A on Pigs" Life 11, no. 2: 157. https://doi.org/10.3390/life11020157
APA StyleLi, Y., Zhang, Y., Liao, Y., Sun, Y., Ruan, Y., Liu, C., Zhang, M., Li, F., Li, X., Fan, S., Yi, L., Ding, H., Zhao, M., Fan, J., & Chen, J. (2021). Preliminary Evaluation of Protective Efficacy of Inactivated Senecavirus A on Pigs. Life, 11(2), 157. https://doi.org/10.3390/life11020157