
Alterations in Energy Metabolism, Mitochondrial Function and Redox Homeostasis in GK Diabetic Rat Tissues Treated with Aspirin

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
2.4. Measurement of Reactive Oxygen Species (ROS), NADPH Oxidase (NOX) Activity, Lipid Peroxidation (LPO) and Superoxide Dismutase (SOD) Activity
2.5. Measurement of Glutathione (GSH)-Redox Metabolism
2.6. Activities of Energy Metabolizing Enzymes
2.7. Measurement of Mitochondrial Respiratory Enzyme Complexes and Bioenergetics
2.8. Measurement of Cytochrome P450 (CYP 450)-Dependent 2E1 and 3A4 Enzymes
2.9. SDS-PAGE and Western Blot Analysis
2.10. Transmission Electron Microscopy (TEM)
2.11. Statistical Analysis
3. Results
3.1. Body Weights and Blood Glucose Levels after Treatment with ASA
3.2. Effect of Aspirin on Oxidative Stress
3.3. Effect of Aspirin on GSH-Redox Metabolism
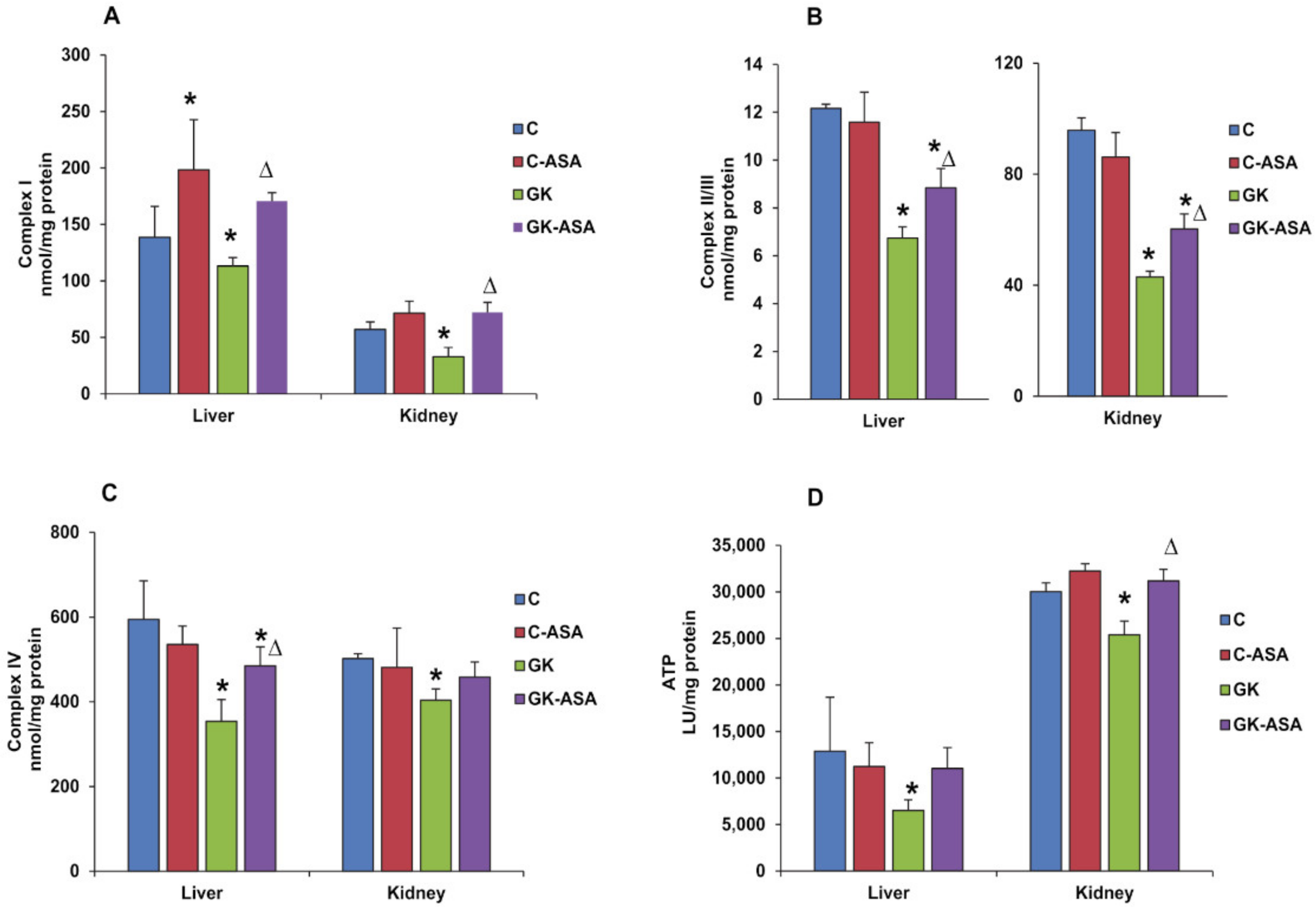
3.4. Effect of Aspirin on Energy Metabolism and Mitochondrial Function
3.5. Effect of Aspirin on CYP450 Enzyme Activities
3.6. Effect of Aspirin on Heme Oxygenase (HO-1) Expression
3.7. Effect of Aspirin on Cell/Insulin Signaling Akt/Glut-4/JNK Markers
3.8. Effect of Aspirin on Intracellular Structural Integrity
4. Discussion
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Östenson, C.-G.; Khan, A.; Abdel-Halim, S.M.; Guenifi, A.; Suzuki, K.; Goto, Y.; Efendic, S. Abnormal insulin secretion and glucose metabolism in pancreatic islets from the spontaneously diabetic GK rat. Diabetologia 1993, 36, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portha, B.; Giroix, M.-H.; Tourrel-Cuzin, C.; Le-Stunff, H.; Movassat, J. The GK Rat: A Prototype for the Study of Non-overweight Type 2 Diabetes. Methods Mol. Biol. 2012, 933, 125–159. [Google Scholar] [CrossRef] [PubMed]
- Tourrel, C.; Bailbe, D.; Lacorne, M.; Meile, M.-J.; Kergoat, M.; Portha, B. Persistent Improvement of Type 2 Diabetes in the Goto-Kakizaki Rat Model by Expansion of the β-Cell Mass During the Prediabetic Period with Glucagon-Like Peptide-1 or Exendin-4. Diabetes 2002, 51, 1443–1452. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, W.M.T.; Panveloski-Costa, A.C.; Yokota, C.N.F.; Pereira, J.N.B.; Filho, J.M.; Torres, R.P.; Hirabara, S.M.; Curi, R.; Alba-Loureiro, T.C. Comparison of Goto-Kakizaki rats and high fat diet-induced obese rats: Are they reliable models to study Type 2 Diabetes mellitus? PLoS ONE 2017, 12, e0189622. [Google Scholar] [CrossRef]
- Takeda, Y.; Shimomura, T.; Asao, H.; Wakabayashi, I. Relationship between Immunological Abnormalities in Rat Models of Diabetes Mellitus and the Amplification Circuits for Diabetes. J. Diabetes Res. 2017, 2017, 4275851. [Google Scholar] [CrossRef]
- Esser, N.; Paquot, N.; Scheen, A.J. Anti-inflammatory agents to treat or prevent type 2 diabetes, metabolic syndrome and cardiovascular disease. Expert Opin. Investig. Drugs 2015, 24, 283–307. [Google Scholar] [CrossRef]
- Raza, H.; John, A.; Howarth, F.C. Alterations in Glutathione Redox Metabolism, Oxidative Stress, and Mitochondrial Function in the Left Ventricle of Elderly Zucker Diabetic Fatty Rat Heart. Int. J. Mol. Sci. 2012, 13, 16241–16254. [Google Scholar] [CrossRef] [Green Version]
- Raza, H.; Prabu, S.K.; John, A.; Avadhani, N.G. Impaired Mitochondrial Respiratory Functions and Oxidative Stress in Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2011, 12, 3133–3147. [Google Scholar] [CrossRef] [Green Version]
- Raza, H.; Prabu, S.K.; Robin, M.-A.; Avadhani, N.G. Elevated mitochondrial cytochrome P450 2E1 and glutathione S-transferase A4-4 in streptozotocin-induced diabetic rats: Tissue-specific variations and roles in oxidative stress. Diabetes 2004, 53, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Raza, H.; John, A.; Shafarin, J.; Howarth, F.C. Exercise-induced alterations in pancreatic oxidative stress and mitochondrial function in type 2 diabetic Goto-Kakizaki rats. Physiol. Rep. 2016, 4, e12751. [Google Scholar] [CrossRef] [Green Version]
- Bellucci, P.N.; Bagnes, M.F.G.; Di Girolamo, G.; Gonzalez, C. Potential Effects of Nonsteroidal Anti-Inflammatory Drugs in the Prevention and Treatment of Type 2 Diabetes Mellitus. J. Pharm. Pr. 2017, 30, 549–556. [Google Scholar] [CrossRef]
- Khalil, M.A.M.; Khalil, M.S.U.D.; Khamis, S.S.A.; Alam, S.; Daiwajna, R.G.; Rajput, A.S.; Alhaji, M.M.; Chong, V.H.; Tan, J. Pros and Cons of Aspirin Prophylaxis for Prevention of Cardiovascular Events in Kidney Transplantation and Review of Evidence. Adv. Prev. Med. 2019, 2019, 6139253. [Google Scholar] [CrossRef]
- Diaz, T.; Trachtenberg, B.H.; Abraham, S.J.K.; Kosagisharaf, R.; Durant-Archibold, A.A. Aspirin Bioactivity for Prevention of Cardiovascular Injury in COVID-19. Front. Cardiovasc. Med. 2020, 7, 562708. [Google Scholar] [CrossRef]
- D’Agati, V. Does aspirin cause acute or chronic renal failure in experimental animals and in humans? Am. J. Kidney Dis. 1996, 28, S24–S29. [Google Scholar] [CrossRef]
- Perneger, T.V.; Whelton, P.K.; Klag, M.J. Risk of Kidney Failure Associated with the Use of Acetaminophen, Aspirin, and Nonsteroidal Antiinflammatory Drugs. New Engl. J. Med. 1994, 331, 1675–1679. [Google Scholar] [CrossRef] [Green Version]
- Somasundaram, S.; Sigthorsson, G.; Simpson, R.; Watts, J.; Jacob, M.; Tavares, I.A.; Rafi, S.; Roseth, A.; Foster, R.; Price, A.B.; et al. Uncoupling of intestinal mitochondrial oxidative phosphorylation and inhibition of cyclooxygenase are required for the development of NSAID-enteropathy in the rat. Aliment. Pharmacol. Ther. 2000, 14, 639–650. [Google Scholar] [CrossRef]
- Ligumsky, M.; Golanska, E.M.; Hansen, D.G.; Kauffman, G.L. Aspirin can inhibit gastric mucosal cyclo-oxygenase without causing lesions in rat. Gastroenterology 1983, 84, 756–761. [Google Scholar] [CrossRef]
- Mashita, Y.; Taniguchi, M.; Yokota, A.; Tanaka, A.; Takeuchi, K. Oral but Not Parenteral Aspirin Upregulates COX-2 Expression in Rat Stomachs. Digestion 2006, 73, 124–132. [Google Scholar] [CrossRef]
- Jacob, J.N.; Badyal, D.K.; Bala, S. Evaluation of theIn VivoAnti-Inflammatory and Analgesic Activity of a Highly Water-Soluble Aspirin Conjugate. Basic Clin. Pharmacol. Toxicol. 2013, 112, 171–174. [Google Scholar] [CrossRef]
- Amiri, L.; John, A.; Shafarin, J.; Adeghate, E.; Jayaprakash, P.; Yasin, J.; Howarth, F.C.; Raza, H. Enhanced Glucose Tolerance and Pancreatic Beta Cell Function by Low Dose Aspirin in Hyperglycemic Insulin-Resistant Type 2 Diabetic Goto-Kakizaki (GK) Rats. Cell. Physiol. Biochem. 2015, 36, 1939–1950. [Google Scholar] [CrossRef]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial dysfunction in type 2 diabetes mellitus: An organ-based analysis. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E268–E285. [Google Scholar] [CrossRef]
- John, A.; Amiri, L.; Shafarin, J.; Howarth, F.C.; Raza, H. Effect of Aspirin on Mitochondrial Dysfunction and Stress in the Pancreas and Heart of Goto-Kakizaki Diabetic Rats. Life 2021, 11, 902. [Google Scholar] [CrossRef]
- Abdin, A.A.; Baalash, A.A.; Hamooda, H.E. Effects of rosiglitazone and aspirin on experimental model of induced type 2 diabetes in rats: Focus on insulin resistance and inflammatory markers. J. Diabetes its Complicat. 2010, 24, 168–178. [Google Scholar] [CrossRef]
- Cao, Y.; Dubois, D.C.; Sun, H.; Almon, R.R.; Jusko, W.J. Modeling Diabetes Disease Progression and Salsalate Intervention in Goto-Kakizaki Rats. J. Pharmacol. Exp. Ther. 2011, 339, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Hammadi, S.H.; Al-Ghamdi, S.S.; Yassien, A.I.; Al-Hassani, S.D. Aspirin and Blood Glucose and Insulin Resistance. Open J. Endocr. Metab. Dis. 2012, 2, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Alnahdi, A.; John, A.; Raza, H. Augmentation of Glucotoxicity, Oxidative Stress, Apoptosis and Mitochondrial Dysfunction in HepG2 Cells by Palmitic Acid. Nutrients 2019, 11, 1979. [Google Scholar] [CrossRef] [Green Version]
- Birch-Machin, M.A.; Turnbull, D.M. Chapter 5 Assaying mitochondrial respiratory complex activity in mitochondria isolated from human cells and tissues. Methods Cell Biol. 2001, 65, 97–117. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeran, M.N.; Al Taee, H.; Azimullah, S.; Tariq, S.; Adeghate, E.; Ojha, S. β-Caryophyllene, a natural bicyclic sesquiterpene attenuates doxorubicin-induced chronic cardiotoxicity via activation of myocardial cannabinoid type-2 (CB2) receptors in rats. Chem.-Biol. Interact. 2019, 304, 158–167. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Bowe, J. Animal models for diabetes: Understanding the pathogenesis and finding new treatments. Biochem. Pharmacol. 2016, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; DuBois, D.C.; Cao, Y.; Jusko, W.J.; Almon, R. Diabetes disease progression in Goto-Kakizaki rats: Effects of salsalate treatment. Diabetes Metab. Syndr. Obesity Targets Ther. 2014, 7, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Griendling, K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.-R.; Harrison, D.G.; Bhatnagar, A. Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef]
- Zorov, D.B.; Filburn, C.R.; Klotz, L.O.; Zweier, J.L.; Sollott, S.J. Reactive oxygen species (ROS)-induced ROS release: A new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J. Exp. Med. 2000, 192, 1001–1014. [Google Scholar] [CrossRef] [Green Version]
- Newsholme, P.; Keane, K.N.; Carlessi, R.; Cruzat, V. Oxidative stress pathways in pancreatic β-cells and insulin-sensitive cells and tissues: Importance to cell metabolism, function, and dysfunction. Am. J. Physiol. Cell Physiol. 2019, 317, C420–C433. [Google Scholar] [CrossRef]
- Zaki, M.S.A.; Haidara, M.A.; Alghamd, M.A.; Shati, A.A.; Wares, A.; Eid, R.A. Protective Effect of Dietary Vitamin E (α Tocopherol) on Artemisinin Induced Oxidative Liver Tissue Damage in Rats. Int. J. Morphol. 2020, 38, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Chen, D. Liver regenerative capacity after partial hepatectomy in rats with nonalcoholic fatty liver disease. Zhonghua Gan Zang Bing Za Zhi 2006, 14, 597–601. [Google Scholar]
- Firneisz, G. Non-alcoholic fatty liver disease and type 2 diabetes mellitus: The liver disease of our age? World J. Gastroenterol. 2014, 20, 9072–9089. [Google Scholar] [CrossRef] [Green Version]
- Pagliassotti, M.J. Endoplasmic Reticulum Stress in Nonalcoholic Fatty Liver Disease. Annu. Rev. Nutr. 2012, 32, 17–33. [Google Scholar] [CrossRef]
- Elsaed, W.M.; Mohamed, H.A. Dietary zinc modifies diabetic-induced renal pathology in rats. Ren. Fail. 2016, 39, 246–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Li, Q.; Zhong, W.; Dong, J.; Wang, Z.; Wang, C. L-carnitine ameliorated fatty liver in high-calorie diet/STZ-induced type 2 diabetic mice by improving mitochondrial function. Diabetol. Metab. Syndr. 2011, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, B.; Klionsky, D.J. Development by Self-Digestion: Molecular Mechanisms and Biological Functions of Autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Abdelsadik, A.; Amin, M.M. Low-dose aspirin improves glucose uptake and attenuates inflammation in rats fed high-fat diet. Egypt. Pharm. J. 2018, 17, 171. [Google Scholar] [CrossRef]
- Dimitrovska, M.; Dervisevik, M.; Cipanovska, N.; Gerazova, K.; Dinevska-Kjovkarovska, S.; Miova, B. Physiological and pharmacological inductors of HSP70 enhance the antioxidative defense mechanisms of the liver and pancreas in diabetic rats. Can. J. Physiol. Pharmacol. 2018, 96, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Lapshina, E.; Sudnikovich, E.; Maksimchik, J.; Zabrodskaya, S.; Zavodnik, L.; Kubyshin, V.; Nocun, M.; Kazmierczak, P.; Dobaczewski, M.; Watala, C. Antioxidative enzyme and glutathione S-transferase activities in diabetic rats exposed to long-term ASA treatment. Life Sci. 2006, 79, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Dervisevik, M.; Dinevska-Kovkarovska, S.; Dimitrovska, M.; Cipanovska, N.; Miova, B. High dose of aspirin moderates diabetes-induced changes of heart glycogen/glucose metabolism in rats. J. Physiol. Sci. 2014, 64, 411–420. [Google Scholar] [CrossRef]
- Mollica, M.P.; Mattace Raso, G.; Cavaliere, G.; Trinchese, G.; De Filippo, C.; Aceto, S.; Prisco, M.; Pirozzi, C.; Di Guida, F.; Lama, A.; et al. Butyrate Regulates Liver Mitochondrial Function, Efficiency, and Dynamics in Insulin-Resistant Obese Mice. Diabetes 2017, 66, 1405–1418. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Kim, H.-S.; Yu, Y.-B.; Kang, H.-C.; Lee, D.-H.; Rhee, M.-H.; Cho, J.-Y.; Park, H.-J. The opposite correlation between calcium ion and cyclic-AMP regarding the activation of microsomal triglyceride transfer protein in rat liver. BMB Rep. 2009, 42, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Rai, U.; Kosuru, R.; Prakash, S.; Singh, S.; Birla, H.; Tiwari, V.; Singh, S. Tetramethylpyrazine prevents diabetes by activating PI3K/Akt/GLUT-4 signalling in animal model of type-2 diabetes. Life Sci. 2019, 236, 116836. [Google Scholar] [CrossRef]
- Whitehead, J.P.; Clark, S.F.; Ursø, B.; James, D.E. Signalling through the insulin receptor. Curr. Opin. Cell Biol. 2000, 12, 222–228. [Google Scholar] [CrossRef]
- Wang, L.-C.; Wu, J.; Zhang, W.; Zhi, Y.; Wu, Y.; Jiang, R.; Yang, R. Effects of aspirin on the ERK and PI3K/Akt signaling pathways in rats with acute pulmonary embolism. Mol. Med. Rep. 2013, 8, 1465–1471. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Fang, X.; Zhao, D.; Mu, Q.; Zuo, J.; Ma, Y.; Zhang, Y.; Mo, F.; Zhang, D.; Jiang, G.; et al. Anti-Diabetic Effects of Jiang Tang Xiao Ke Granule via PI3K/Akt Signalling Pathway in Type 2 Diabetes KKAy Mice. PLoS ONE 2017, 12, e0168980. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Zuberi, A.; Quon, M.J.; Dong, Z.; Ye, J. Aspirin Inhibits Serine Phosphorylation of Insulin Receptor Substrate 1 in Tumor Necrosis Factor-treated Cells through Targeting Multiple Serine Kinases. J. Biol. Chem. 2003, 278, 24944–24950. [Google Scholar] [CrossRef] [Green Version]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxidative Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are Oxidative Stress−Activated Signaling Pathways Mediators of Insulin Resistance and β-Cell Dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pessler, D.; Rudich, A.; Bashan, N. Oxidative stress impairs nuclear proteins binding to the insulin responsive element in the GLUT4 promoter. Diabetologia 2001, 44, 2156–2164. [Google Scholar] [CrossRef] [Green Version]
- Day, C.P. Genetic and Environmental Susceptibility to Non-Alcoholic Fatty Liver Disease. Dig. Dis. 2010, 28, 255–260. [Google Scholar] [CrossRef]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The Glutamate Dehydrogenase Pathway and Its Roles in Cell and Tissue Biology in Health and Disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Yraola, A.; Bakhit, S.M.; Franke, P.; Weise, C.; Schweiger, M.; Jorcke, D.; Ziegler, M. Regulation of glutamate dehydrogenase by reversible ADP-ribosylation in mitochondria. EMBO J. 2001, 20, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Göhring, I.; Mulder, H. Glutamate dehydrogenase, insulin secretion, and type 2 diabetes: A new means to protect the pancreatic β-cell? J. Endocrinol. 2012, 212, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Agil, A.; Chayah, M.; Visiedo, L.; Navarro-Alarcon, M.; Ferrer, J.M.R.; Tassi, M.; Reiter, R.J.; Fernández-Vázquez, G. Melatonin Improves Mitochondrial Dynamics and Function in the Kidney of Zücker Diabetic Fatty Rats. J. Clin. Med. 2020, 9, 2916. [Google Scholar] [CrossRef] [PubMed]
- Ahishali, E.; Demir, K.; Ahishali, B.; Akyuz, F.; Pinarbasi, B.; Poturoglu, S.; Ibrisim, D.; Gulluoglu, M.; Ozdil, S.; Besisik, F.; et al. Electron microscopic findings in non-alcoholic fatty liver disease: Is there a difference between hepatosteatosis and steatohepatitis? J. Gastroenterol. Hepatol. 2010, 25, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Dubinin, M.V. Diabetes Mellitus, Mitochondrial Dysfunction and Ca2+-Dependent Permeability Transition Pore. Int. J. Mol. Sci. 2020, 21, 6559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, M.; Dong, H.; Yu, X.; Zhang, J. Anti-hypoglycemic and hepatocyte-protective effects of hyperoside from Zanthoxylum bungeanum leaves in mice with high-carbohydrate/high-fat diet and alloxan-induced diabetes. Int. J. Mol. Med. 2017, 41, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamble, P.; Selvarajan, K.; Narasimhulu, C.A.; Nandave, M.; Parthasarathy, S. Aspirin may promote mitochondrial biogenesis via the production of hydrogen peroxide and the induction of Sirtuin1/PGC-1α genes. Eur. J. Pharmacol. 2013, 699, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemoto, S.; Fergusson, M.M.; Finkel, T. SIRT1 Functionally Interacts with the Metabolic Regulator and Transcriptional Coactivator PGC-1α. J. Biol. Chem. 2005, 280, 16456–16460. [Google Scholar] [CrossRef] [Green Version]
- Miwa, S.; Lawless, C.; von Zglinicki, T. Mitochondrial turnover in liver is fastin vivoand is accelerated by dietary restriction: Application of a simple dynamic model. Aging Cell 2008, 7, 920–923. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
John, A.; Amiri, L.; Shafarin, J.; Tariq, S.; Adeghate, E.; Howarth, F.C.; Raza, H. Alterations in Energy Metabolism, Mitochondrial Function and Redox Homeostasis in GK Diabetic Rat Tissues Treated with Aspirin. Life 2022, 12, 104. https://doi.org/10.3390/life12010104
John A, Amiri L, Shafarin J, Tariq S, Adeghate E, Howarth FC, Raza H. Alterations in Energy Metabolism, Mitochondrial Function and Redox Homeostasis in GK Diabetic Rat Tissues Treated with Aspirin. Life. 2022; 12(1):104. https://doi.org/10.3390/life12010104
Chicago/Turabian StyleJohn, Annie, Layla Amiri, Jasmin Shafarin, Saeed Tariq, Ernest Adeghate, Frank Christopher Howarth, and Haider Raza. 2022. "Alterations in Energy Metabolism, Mitochondrial Function and Redox Homeostasis in GK Diabetic Rat Tissues Treated with Aspirin" Life 12, no. 1: 104. https://doi.org/10.3390/life12010104
APA StyleJohn, A., Amiri, L., Shafarin, J., Tariq, S., Adeghate, E., Howarth, F. C., & Raza, H. (2022). Alterations in Energy Metabolism, Mitochondrial Function and Redox Homeostasis in GK Diabetic Rat Tissues Treated with Aspirin. Life, 12(1), 104. https://doi.org/10.3390/life12010104