
Two Retrotransposon Elements in Intron of Porcine BMPR1B Is Associated with Phenotypic Variation

 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and DNA Isolation
2.2. RIPs Excavation
2.3. RIPs Identification by PCR
2.4. Population Diversity Based on RIPs
2.5. qPCR
2.6. Dual-Luciferase Reporter Assay
2.7. Statistical Analysis
2.8. Animal Welfare
3. Results
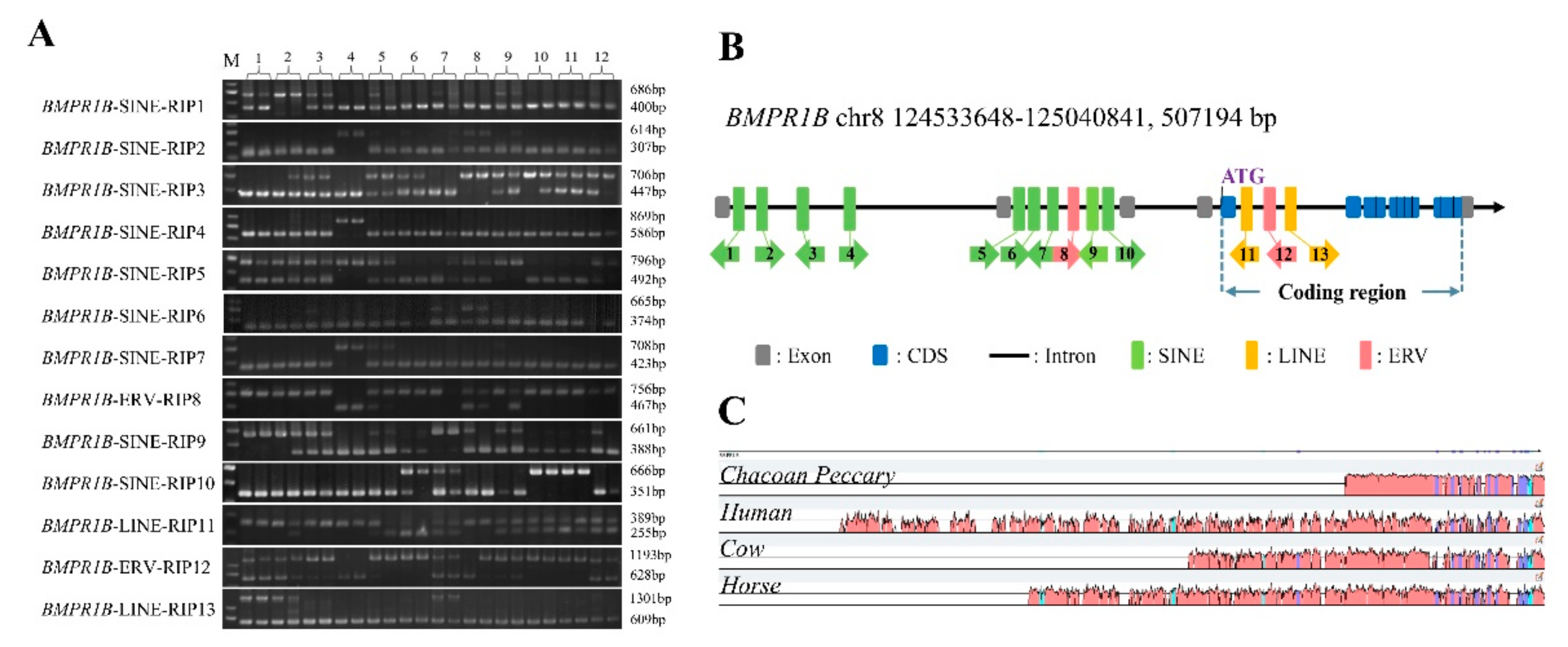
3.1. Thirteen RIPs Were Verified by PCR in the Pig BMPR1B Gene
3.2. Association between RIPs and Phenotype of Large White Pigs
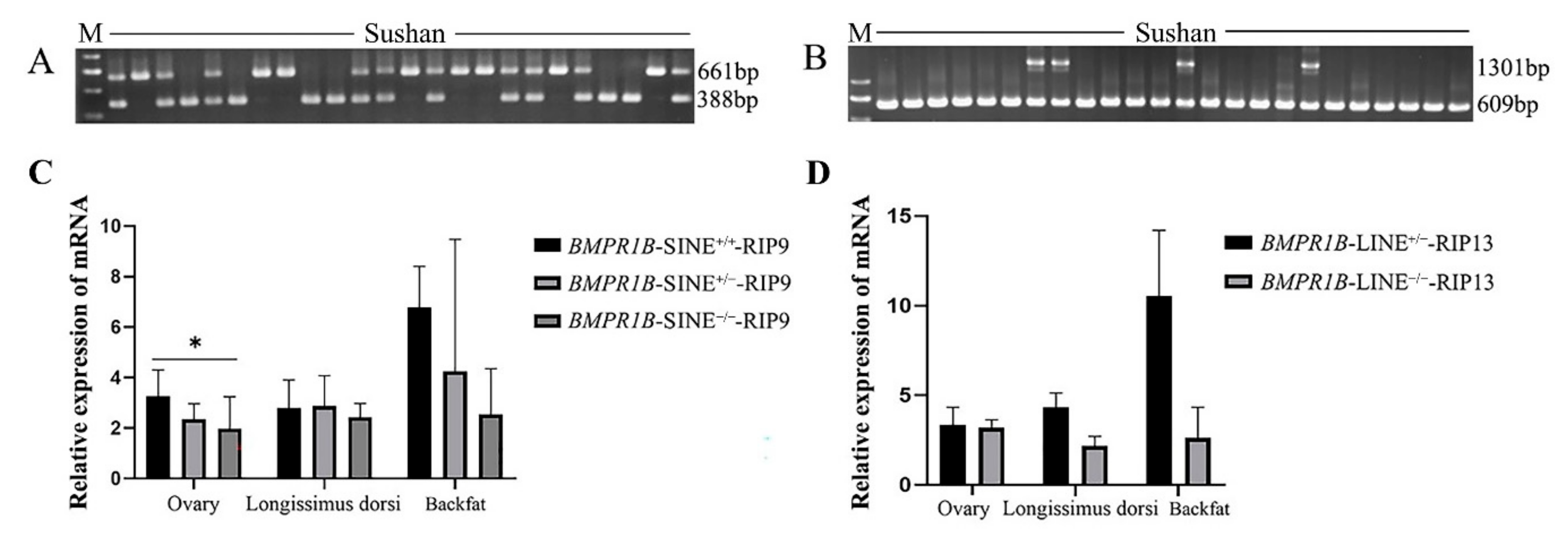
3.3. BMPR1B-SINE-RIP9 and BMPR1B-LINE-RIP13 Distribution in the Different Pig Breeds
3.4. The Expression Pattern of Pig BMPR1B in Sushan Pigs for Two RIPs
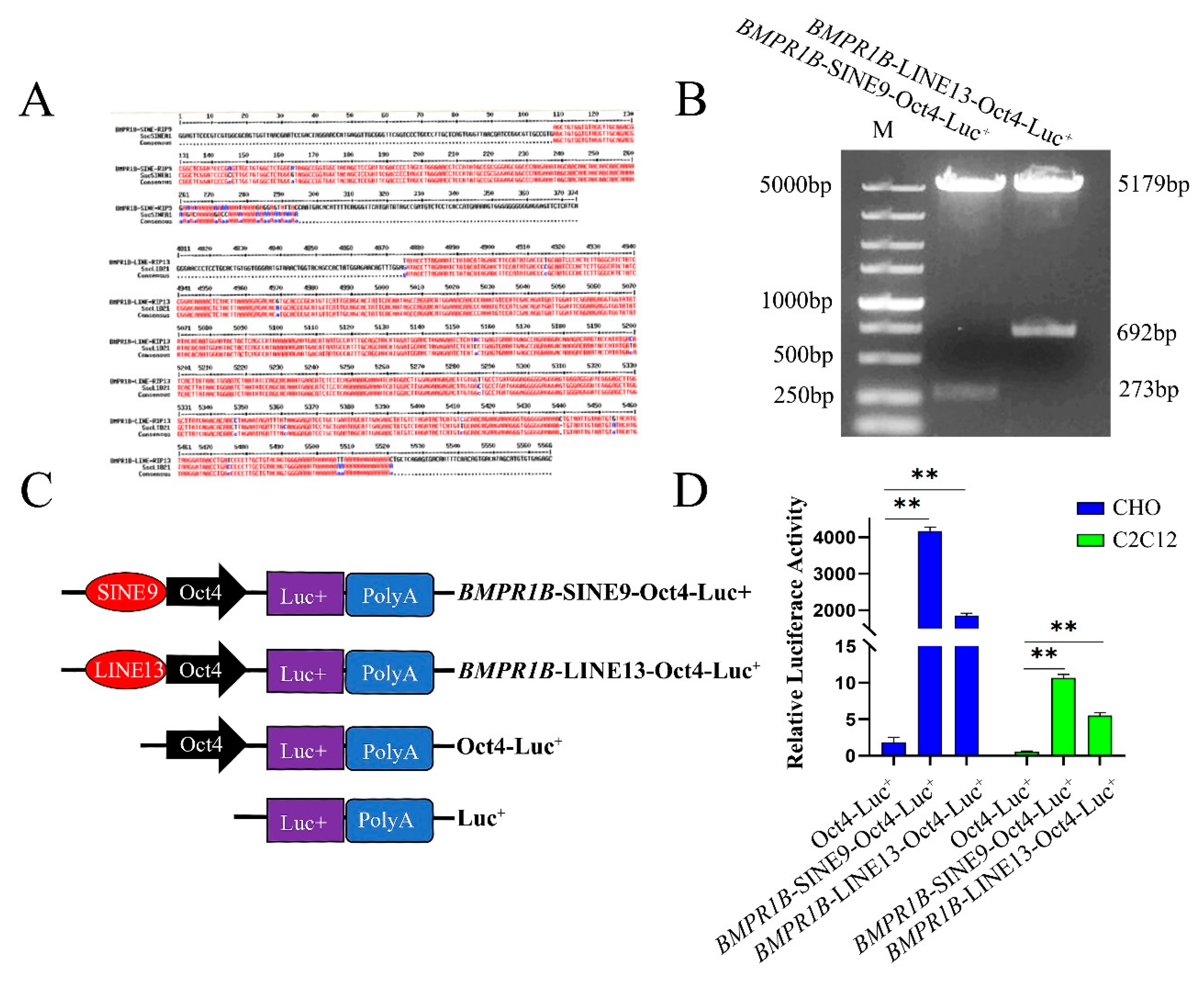
3.5. SINE and LINE Insertion Might Act as an Enhancer in CHO and C2C12 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamaji, N.; Celeste, A.J.; Thies, R.S.; Song, J.J.; Bernier, S.M.; Goltzman, D.; Lyons, K.M.; Nove, J.; Rosen, V.; Wozney, J.M. A mammalian serine/threonine kinase receptor specifically binds BMP-2 and BMP-4. Biochem. Biophys. Res. Commun. 1994, 205, 1944–1951. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.S.; Ovchinnikov, D.A.; Yoshii, I.; Mishina, Y.; Behringer, R.R.; Lyons, K.M. Bmpr1a and Bmpr1b have overlapping functions and are essential for chondrogenesis in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 5062–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartori, R.; Sandri, M. BMPs and the muscle–bone connection. Bone 2015, 80, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.J.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. Bone morphogenetic proteins and folliculogenesis: Lessons from the Booroola mutation. Reprod. Suppl. 2003, 61, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, W.; Yeh, L.C.; Gmyrek, A.; Lee, J.C.; Lee, J.C. Long-Range Communication Network in the Type 1B Bone Morphogenetic Protein Receptor. Biochemistry 2015, 54, 7079–7088. [Google Scholar] [CrossRef]
- Renault, L.; Patiño, L.C.; Magnin, F.; Delemer, B.; Young, J.; Laissue, P.; Binart, N.; Beau, I. BMPR1A and BMPR1B missense mutations cause primary ovarian insufficiency. J. Clin. Endocrinol. Metab. 2020, 105, e1449–e1457. [Google Scholar] [CrossRef]
- França, M.M.; Mendonca, B.B. Genetics of ovarian insufficiency and defects of folliculogenesis. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 36, 101594. [Google Scholar] [CrossRef]
- Yang, Y.; Yuan, J.; Yao, X.; Zhang, R.; Yang, H.; Zhao, R.; Guo, J.; Jin, K.; Mei, H.; Luo, Y.; et al. BMPR1B mutation causes Pierre Robin sequence. Oncotarget 2017, 8, 25864–25871. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Basit, S.; Khan, M.A.; Muhammad, N.; Ahmad, W. Genetics of human isolated acromesomelic dysplasia. Eur. J. Med. Genet. 2016, 59, 198–203. [Google Scholar] [CrossRef]
- Alshangiti, A.M.; Wyatt, S.L.; McCarthy, E.; Collins, L.M.; Hegarty, S.V.; Sullivan, A.M.; O′Keeffe, G.W. Association of distinct type 1 bone morphogenetic protein receptors with different molecular pathways and survival outcomes in neuroblastoma. Neuronal Signal 2020, 4, NS20200006. [Google Scholar] [CrossRef]
- Long, F.; Shi, H.; Li, P.; Guo, S.; Ma, Y.; Wei, S.; Li, Y.; Gao, F.; Gao, S.; Wang, M.; et al. A SMOC2 variant inhibits BMP signaling by competitively binding to BMPR1B and causes growth plate defects. Bone 2021, 142, 115686. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Merino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, C.J.H.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.; Wu, X.Y.; Juengel, J.L.; Ross, I.K.; Lumsden, J.M.; Lord, E.A.; Dodds, K.G.; Walling, G.A.; McEwan, J.C.; O′Connell, A.R.; et al. Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetic protein IB receptor (ALK-6) that is expressed in both oocytes and granulosa cells. Biol. Reprod. 2001, 64, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Fabre, S.; Pierre, A.; Mulsant, P.; Bodin, L.; Di Pasquale, E.; Persani, L.; Monget, P.; Monniaux, D. Regulation of ovulation rate in mammals: Contribution of sheep genetic models. Reprod. Biol. Endocrinol. 2006, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, G.H.; Yang, L.G. A review of research progress of FecB gene in Chinese breeds of sheep. Anim. Reprod. Sci. 2009, 116, 1–9. [Google Scholar] [CrossRef]
- Li, D.; Zhang, L.; Wang, Y.; Chen, X.; Li, F.; Yang, L.; Cui, J.; Li, R.; Cao, B.; An, X.; et al. FecB mutation and litter size are associated with a 90-base pair deletion in BMPR1B in East Friesian and Hu crossbred sheep. Anim. Biotechnol. 2022, 5, 1–10. [Google Scholar] [CrossRef]
- Tang, J.; Hu, W.; Di, R.; Liu, Q.; Wang, X.; Zhang, X.; Zhang, J.; Chu, M. Expression analysis of the prolific candidate genes, BMPR1B, BMP15, and GDF9 in Small Tail Han ewes with three fecundity (FecB gene) genotypes. Animals 2018, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Guo, X.; He, X.; Di, R.; Zhang, X.; Zhang, J.; Wang, X.; Chu, M. Insight into Pituitary lncRNA and mRNA at Two Estrous Stages in Small Tail Han Sheep with Different FecB Genotypes. Front. Endocrinol. 2021, 12, 789564. [Google Scholar] [CrossRef]
- Wen, Y.-L.; Guo, X.-F.; Ma, L.; Zhang, X.-S.; Zhang, J.-L.; Zhao, S.-G.; Chu, M.-X. The expression and mutation of BMPR1B and its association with litter size in small-tail Han sheep (Ovis aries). Arch. Anim. Breed. 2021, 64, 211–221. [Google Scholar] [CrossRef]
- Zhou, S.; Ding, Y.; Liu, J.; Liu, Y.; Zhao, X.; Li, G.; Zhang, C.; Li, C.; Wang, Y.; Kalds, P.; et al. Highly efficient generation of sheep with a defined FecB(B) mutation via adenine base editing. Genet. Sel. Evol. 2020, 52, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Q.; Zhao, K.; Jiang, C.; Ren, D.; Xu, P.; He, X.; Liao, R.; Jiang, K.; Ma, J.; et al. Production of Transgenic Pigs with an Introduced Missense Mutation of the Bone Morphogenetic Protein Receptor Type IB Gene Related to Prolificacy. Asian-Australas. J. Anim. Sci. 2016, 29, 925–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Chen, Q.; Gui, L.; Jin, J.; Li, Y.; Ru, Q.; Hou, S. Association of polymorphisms in bone morphogenetic protein receptor-1B gene exon-9 with litter size in Dorset, Mongolian, and Small Tail Han ewes. Asian-Australas. J. Anim. Sci. 2019, 32, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, H.; Akhatayeva, Z.; Liu, H.; Lin, C.; Han, X.; Lu, X.; Lan, X.; Zhang, Q.; Pan, C. Novel indel variations of the sheep FecB gene and their effects on litter size. Gene 2021, 767, 145176. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-T.; Zhang, M.-M.; Li, Q.-G.; Tang, H.; Zhang, L.-F.; Wang, K.-J.; Zhu, M.-Z.; Lu, Y.-F.; Bao, H.-G.; Zhang, Y.-M. Whole-genome resequencing reveals candidate mutations for pig prolificacy. Proc. R. Soc. B Biol. Sci. 2017, 284, 20172437. [Google Scholar] [CrossRef] [Green Version]
- Platt, R.N.; Vandewege, M.W.; Ray, D.A. Mammalian transposable elements and their impacts on genome evolution. Chromosome Res. 2018, 26, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Hermant, C.; Torres-Padilla, M.E. TFs for TEs: The transcription factor repertoire of mammalian transposable elements. Genes Dev. 2021, 35, 22–39. [Google Scholar] [CrossRef]
- Sundaram, V.; Wysocka, J. Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes. Philos. Trans. R. Soc. B 2020, 375, 20190347. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; He, M.; Tang, Q.; Tian, S.; Zhang, J.; Li, Y.; Wang, D.; Jin, L.; Ning, C.; Zhu, W. Comparative 3D genome architecture in vertebrates. BMC Biol. 2022, 20, 99. [Google Scholar] [CrossRef]
- Kalendar, R.; Schulman, A.H. Transposon-based tagging: IRAP, REMAP, and iPBS. In Molecular Plant Taxonomy; Springer: Berlin/Heidelberg, Germany, 2014; Volume 1115, pp. 233–255. [Google Scholar]
- Amiteye, S. Basic concepts and methodologies of DNA marker systems in plant molecular breeding. Heliyon 2021, 7, e08093. [Google Scholar] [CrossRef]
- Wang, Z.; Qu, L.; Yao, J.; Yang, X.; Li, G.; Zhang, Y.; Li, J.; Wang, X.; Bai, J.; Xu, G.; et al. An EAV-HP insertion in 5′ Flanking region of SLCO1B3 causes blue eggshell in the chicken. PLoS Genet. 2013, 9, e1003183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballif, B.; Ramirez, C.; Carl, C.; Sundin, K.; Krug, M.; Zahand, A.; Shaffer, L.; Flores-Smith, H. The PMEL Gene and Merle in the Domestic Dog: A Continuum of Insertion Lengths Leads to a Spectrum of Coat Color Variations in Australian Shepherds and Related Breeds. Cytogenet. Genome Res. 2018, 156, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Davis, B.W.; Jern, P.; Dorshorst, B.J.; Siegel, P.B.; Andersson, L. Characterization of the endogenous retrovirus insertion in CYP19A1 associated with henny feathering in chicken. Mob. DNA 2019, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dalirsefat, S.B.; Han, D.; Dong, X.; Hua, G.; Zheng, X.; Xia, T.; Shao, T.; Deng, X.; Wu, C. An EAV-HP insertion in the 5 flanking region of SLCO1B3 is associated with its tissue-expression profile in blue-eggshell Yimeng chickens (Gallus gallus)-ScienceDirect. Poult. Sci. 2020, 99, 6371–6377. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zheng, Y.; Wang, M.; Murani, E.; D’Alessandro, E.; Moawad, A.S.; Wang, X.; Wimmers, K.; Song, C. SINE Insertion in the Intron of Pig GHR May Decrease Its Expression by Acting as a Repressor. Animals 2021, 11, 1871. [Google Scholar] [CrossRef]
- Liang, D.; Zhao, P.; Si, J.; Fang, L.; Pairo-Castineira, E.; Hu, X.; Xu, Q.; Hou, Y.; Gong, Y.; Liang, Z. Genomic analysis revealed a convergent evolution of LINE-1 in coat color: A case study in water buffaloes (Bubalus bubalis). Mol. Biol. Evol. 2021, 38, 1122–1136. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.; Murani, E.; D′Alessandro, E.; An, Y.; Chen, C.; Li, K.; Galeano, G.; Wimmers, K.; Song, C. A 192 bp ERV fragment insertion in the first intron of porcine TLR6 may act as an enhancer associated with the increased expressions of TLR6 and TLR1. Mob. DNA 2021, 12, 20. [Google Scholar] [CrossRef]
- Du, Z.; D′Alessandro, E.; Zheng, Y.; Wang, M.; Chen, C.; Wang, X.; Song, C. Retrotransposon Insertion Polymorphisms (RIPs) in Pig Coat Color Candidate Genes. Animals 2022, 12, 969. [Google Scholar] [CrossRef]
- Wang, X.; D′Alessandro, E.; Chi, C.; Moawad, A.S.; Zong, W.; Chen, C.; Song, C. Genetic Evaluation and Population Structure of Jiangsu Native Pigs in China Revealed by SINE Insertion Polymorphisms. Animals 2022, 12, 1345. [Google Scholar] [CrossRef]
- Yeh, F.; Yang, R.; Boyle, T. POPGENE Version 1.32: Microsoft Windows–Based Freeware for Population Genetic Analysis, Quick User Guide; Center for International Forestry Research, University of Alberta: Edmonton, AB, Canada, 1999; pp. 1–29. [Google Scholar]
- Chan, S.; Shen, D.; Sang, Y.; Wang, S.; Wang, Y.; Chen, C.; Gao, B.; Song, C. Development of enhancer-trapping and-detection vectors mediated by the Tol2 transposon in zebrafish. PeerJ 2019, 7, e6862. [Google Scholar] [CrossRef]
- Wozney, J.M.; Rosen, V.; Celeste, A.J.; Mitsock, L.M.; Whitters, M.J.; Kriz, R.W.; Hewick, R.M.; Wang, E.A. Novel regulators of bone formation: Molecular clones and activities. Science 1988, 242, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Romanish, M.T.; Mager, D.L. Distributions of transposable elements reveal hazardous zones in mammalian introns. PLoS Comput. Biol. 2011, 7, e1002046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malicki, M.; Iliopoulou, M.; Hammann, C. Retrotransposon Domestication and Control in Dictyostelium discoideum. Front. Microbiol. 2017, 8, 1869. [Google Scholar] [CrossRef]
- Chen, C.; Wang, W.; Wang, X.; Shen, D.; Wang, S.; Wang, Y.; Gao, B.; Wimmers, K.; Mao, J.K.; Song, C. Retrotransposons evolution and impact on lncRNA and protein coding genes in pigs. Mob. DNA 2019, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Philippsen, G.S.; DeMarco, R. Impact of transposable elements in the architecture of genes of the human parasite Schistosoma mansoni. Mol. Biochem. Parasitol. 2019, 228, 27–31. [Google Scholar] [CrossRef]
- Hernandez, S.; Finlayson, H.; Ashworth, C.; Haley, C.; Archibald, A. A genome-wide linkage analysis for reproductive traits in F 2 L arge W hite× M eishan cross gilts. Anim. Genet. 2014, 45, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Legault, C. Selection of breeds, strains and individual pigs for prolificacy. J. Reprod. Fertil. Suppl. 1985, 33, 151–166. [Google Scholar] [CrossRef]
- Liu, X.; Xiong, X.; Yang, J.; Zhou, L.; Yang, B.; Ai, H.; Ma, H.; Xie, X.; Huang, Y.; Fang, S. Genome-wide association analyses for meat quality traits in Chinese Erhualian pigs and a Western Duroc×(Landrace× Yorkshire) commercial population. Genet. Sel. Evol. 2015, 47, 44. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, X.; Jing, X.; He, X.; Wang, L.; Liu, Y.; Liu, D. Transcriptomics analysis on excellent meat quality traits of skeletal muscles of the Chinese indigenous min pig compared with the large white breed. Int. J. Mol. Sci. 2017, 19, 21. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Li, P.; Zhu, M.; He, L.; Sui, S.; Gao, S.; Su, G.; Ding, N.; Huang, Y.; Lu, Z. Genome-wide association analysis reveals genomic regions on Chromosome 13 affecting litter size and candidate genes for uterine horn length in Erhualian pigs. Animal 2018, 12, 2453–2461. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, W.; Cai, J.; Ni, Y.; Xiao, L.; Zhang, J. Transcriptome analysis in comparing carcass and meat quality traits of Jiaxing Black Pig and Duroc× Duroc× Berkshire× Jiaxing Black Pig crosses. Gene 2022, 808, 145978. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Du, H.; Chen, J.; Yang, G.; Zhang, X. Porcine growth differentiation factor 9 gene polymorphisms and their associations with litter size. J. Genet. Genom. 2008, 35, 163–169. [Google Scholar] [CrossRef]
- Tao, H.; Mei, S.; Sun, X.; Peng, X.; Zhang, X.; Ma, C.; Wang, L.; Hua, L.; Li, F. Associations of TCF12, CTNNAL1 and WNT10B gene polymorphisms with litter size in pigs. Anim. Reprod. Sci. 2013, 140, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Charoensook, R.; Gatphayak, K.; Brenig, B.; Knorr, C. Genetic diversity analysis of Thai indigenous pig population using microsatellite markers. Asian-Australas. J. Anim. Sci. 2019, 32, 1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Seo, D.W.; Cho, E.S.; Choi, B.H.; Kim, Y.M.; Hong, J.K.; Han, H.D.; Jung, Y.B.; Kim, D.J.; Choi, T.J. Genetic diversity and ancestral study for Korean Native Pigs using 60K SNP chip. Animals 2020, 10, 760. [Google Scholar] [CrossRef] [PubMed]
- Mishina, Y.; Suzuki, A.; Ueno, N.; Behringer, R.R. Bmpr encodes a type I bone morphogenetic protein receptor that is essential for gastrulation during mouse embryogenesis. Genes Dev. 1995, 9, 3027–3037. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.Y.; Di, R.; Tang, Q.Q.; Jin, H.H.; Chu, M.X.; Huang, D.W.; He, J.N.; Liu, Q.Y.; Hu, W.P.; Wang, X.Y.; et al. Tissue-specific mRNA expression profiles of GDF9, BMP15, and BMPR1B genes in prolific and non-prolific goat breeds. Czech J. Anim. Sci. 2015, 60, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvak, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Loci | Insertion in Exon/Intron | Length (bp) | Direction | Type | Chromosome (Sus scrofa11.1) |
|---|---|---|---|---|---|
| BMPR1B-SINE-RIP1 | Intron1 | 286 | − | SINEA1 | 125,030,204–125,030,205 |
| BMPR1B-SINE-RIP2 | Intron1 | 307 | + | SINEA1 | 125,020,478–125,020,478 |
| BMPR1B-SINE-RIP3 | Intron1 | 259 | − | SINEA3 | 124,975,543–124,975,544 |
| BMPR1B-SINE-RIP4 | Intron1 | 283 | + | SINEA1 | 124,934,102–124,934,103 |
| BMPR1B-SINE-RIP5 | Intron2 | 304 | + | SINEA1 | 124,833,678–124,833,679 |
| BMPR1B-SINE-RIP6 | Intron2 | 291 | + | SINEA1 | 124,831,282–124,831,283 |
| BMPR1B-SINE-RIP7 | Intron2 | 285 | − | SINEA1 | 124,829,563–124,829,564 |
| BMPR1B-ERV-RIP8 | Intron2 | 289 | + | ERVIII | 124,804,706–124,804,995 |
| BMPR1B-SINE-RIP9 | Intron2 | 273 | − | SINEA1 | 124,800,210–124,800,483 |
| BMPR1B-SINE-RIP10 | Intron2 | 315 | + | SINEA1 | 124,788,936–124,788,937 |
| BMPR1B-LINE-RIP11 | Intron5 | 134 | − | L1D11 | 124,607,615–124,607,749 |
| BMPR1B-ERV-RIP12 | Intron5 | 565 | − | ERVIII | 124,606,427–124,606,428 |
| BMPR1B-LINE-RIP13 | Intron5 | 692 | + | L1D21 | 124,595,110–124,595,802 |
| RIPs | Genotype | Number | Thickness of Back Fat/mm | Age at 30 kg Body Weight/d | Age at 100 kg Body Weight/d |
|---|---|---|---|---|---|
| BMPR1B-SINE-RIP9 | SINE+\+ | 168 | 10.51 ± 0.20 | 74.99 ± 0.71 | 162.14 ± 0.55 a |
| SINE+\− | 286 | 10.87 ± 0.17 | 74.91 ± 0.52 | 163.71 ± 0.52 A | |
| SINE−\− | 75 | 11.25 ± 0.34 | 74.89 ± 1.27 | 160.80 ± 0.90 b | |
| BMPR1B-LINE-RIP13 | LINE+/+ | 259 | 10.65 ± 0.17 | 74.92 ± 0.52 a | 163.25 ± 0.56 a |
| LINE+/− | 115 | 10.67 ± 0.28 | 74.36 ± 0.94 a | 161.97 ± 0.76 ab | |
| LINE−/− | 21 | 11.62 ± 0.62 | 70.07 ± 2.58 b | 159.26 ± 1.48 b |
| Genotype | Number | First Litter Size | Number Born Alive | Born Weight/kg |
|---|---|---|---|---|
| SINE+/+ | 99 | 10.47 ± 0.28 a | 9.86 ± 0.26 | 13.42 ± 0.36 |
| SINE+/− | 118 | 10.03 ± 0.23 ab | 9.49 ± 0.24 | 12.73 ± 0.33 |
| SINE−/− | 26 | 9.19 ± 0.57 b | 8.88 ± 0.63 | 11.85 ± 0.90 |
| Polymorphic Site | Breeds | Number | Genotype Frequency | Allele Frequency | Hardy-Weinberg Equilibrium | Polymorphic Information Content | |||
|---|---|---|---|---|---|---|---|---|---|
| +/+ | +/− | −/− | + | − | |||||
| BMPR1B-SINE-RIP9 | Large White | 529 | 31.76 | 54.06 | 14.18 | 58.79 | 41.21 | 0.01 | 0.367 |
| Landrace | 24 | 50.00 | 37.50 | 12.50 | 68.75 | 31.25 | 0.53 | 0.337 | |
| Sushan | 24 | 33.33 | 41.67 | 25.00 | 54.17 | 45.83 | 0.43 | 0.373 | |
| Sujiang | 21 | 80.95 | 19.05 | 0.00 | 90.48 | 9.52 | 0.63 | 0.157 | |
| Wuzhishan | 24 | 4.17 | 20.83 | 75.00 | 14.58 | 85.42 | 0.42 | 0.218 | |
| BMPR1B-LINE-RIP13 | Large White | 395 | 66.57 | 29.11 | 5.32 | 80.13 | 19.87 | 0.09 | 0.268 |
| Duroc | 24 | 62.50 | 29.17 | 8.33 | 77.08 | 22.92 | 0.39 | 0.291 | |
| Landrace | 24 | 20.83 | 37.50 | 41.67 | 39.58 | 60.42 | 0.29 | 0.364 | |
| Sushan | 32 | 0 | 15.63 | 84.38 | 7.81 | 92.19 | 0.63 | 0.134 | |
| Sujiang | 21 | 4.76 | 19.05 | 76.19 | 14.29 | 85.71 | 0.31 | 0.215 | |
| Jinhua | 24 | 0 | 16.67 | 83.33 | 8.33 | 91.67 | 0.66 | 0.141 | |
| Erhualian | 24 | 0 | 4.17 | 95.83 | 2.08 | 97.92 | 0.92 | 0.040 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chi, C.; He, J.; Du, Z.; Zheng, Y.; D’Alessandro, E.; Chen, C.; Moawad, A.S.; Asare, E.; Song, C.; Wang, X. Two Retrotransposon Elements in Intron of Porcine BMPR1B Is Associated with Phenotypic Variation. Life 2022, 12, 1650. https://doi.org/10.3390/life12101650
Chi C, He J, Du Z, Zheng Y, D’Alessandro E, Chen C, Moawad AS, Asare E, Song C, Wang X. Two Retrotransposon Elements in Intron of Porcine BMPR1B Is Associated with Phenotypic Variation. Life. 2022; 12(10):1650. https://doi.org/10.3390/life12101650
Chicago/Turabian StyleChi, Chenglin, Jia He, Zhanyu Du, Yao Zheng, Enrico D’Alessandro, Cai Chen, Ali Shoaib Moawad, Emmanuel Asare, Chengyi Song, and Xiaoyan Wang. 2022. "Two Retrotransposon Elements in Intron of Porcine BMPR1B Is Associated with Phenotypic Variation" Life 12, no. 10: 1650. https://doi.org/10.3390/life12101650
APA StyleChi, C., He, J., Du, Z., Zheng, Y., D’Alessandro, E., Chen, C., Moawad, A. S., Asare, E., Song, C., & Wang, X. (2022). Two Retrotransposon Elements in Intron of Porcine BMPR1B Is Associated with Phenotypic Variation. Life, 12(10), 1650. https://doi.org/10.3390/life12101650