

MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy
Abstract
:1. Introduction
2. Results
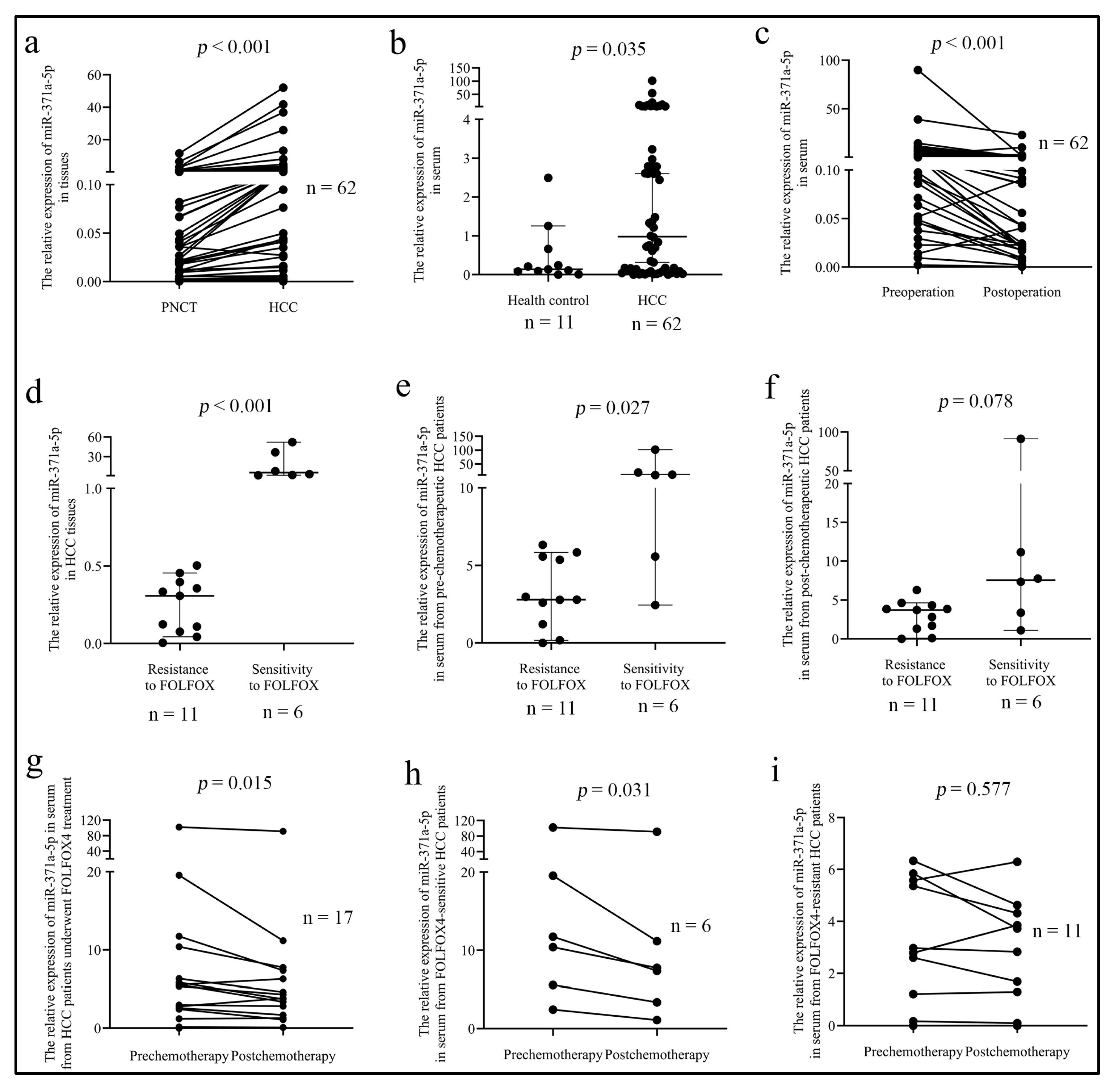
2.1. MiR-371a-5p Is Associated with HCC Malignant Characteristics and the Response of Patients to FOLFOX4 Therapy
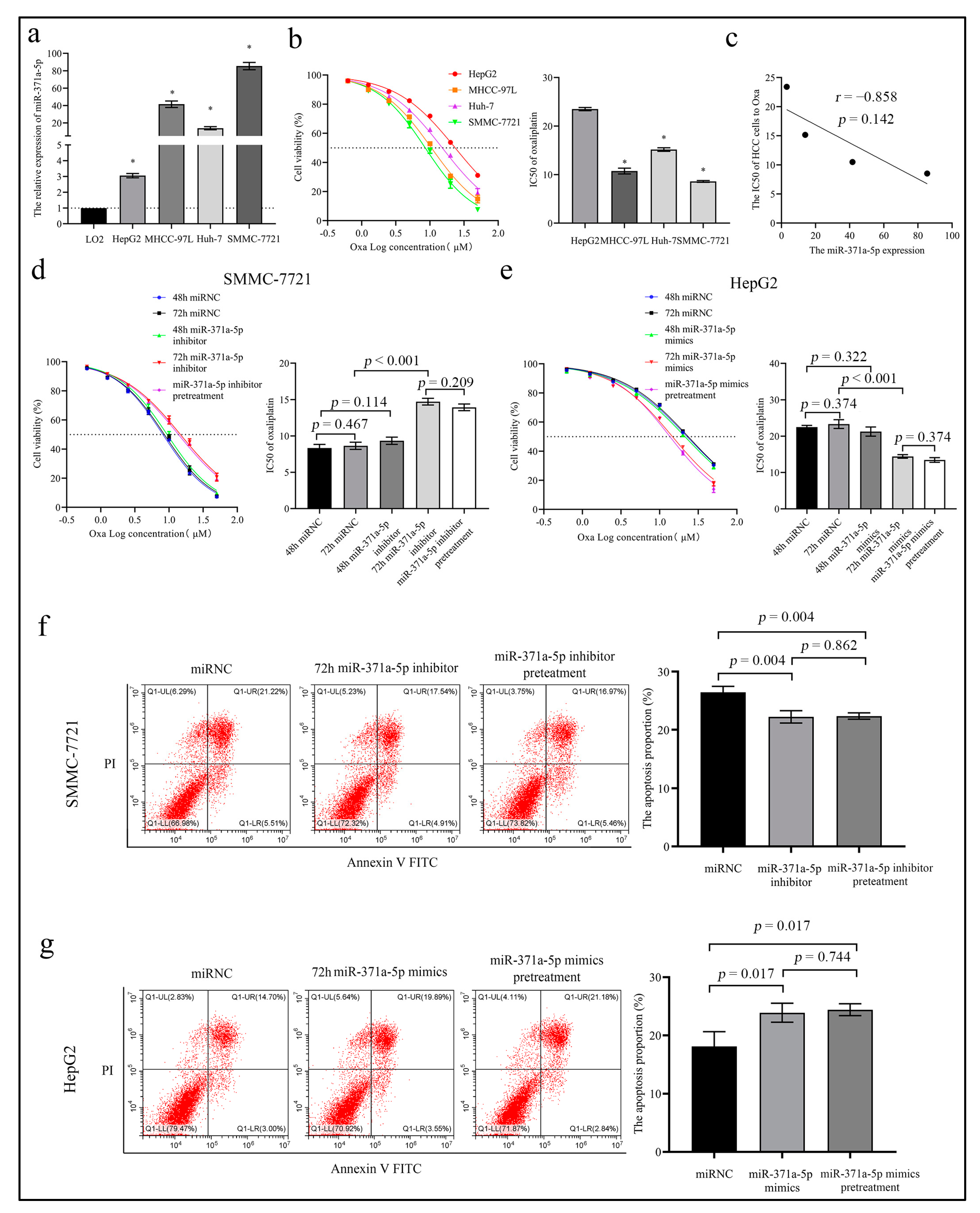
2.2. MiR-371a-5p Enhances the Response of HCC Cells to Oxaliplatin In Vitro
2.3. MiR-371a-5p Represses HCC Cell Autophagy by Target Suppression of BECN1
2.4. Inhibition of BECN1-Dependent Autophagy Is Essential for the miR-371a-5p-Caused Sensitivity of HCC Cells to Oxaliplatin In Vitro
2.5. BECN1 Is Down-Regulated in HCC Tissues and Associated with HCC Malignant Characteristics
3. Discussion
4. Material and Methods
4.1. Clinical Samples
4.2. Cell Lines and Culture
4.3. Vectors, Oligonucleotides and Cell Transfection
4.4. Drug Treatment and Grouping
4.5. Quantitative Real-Time Polymerase Chain Reaction (QPCR)
4.6. Western-Blot (WB) and Immunohistochemistry (IHC)
4.7. Luciferase Reporter Assay
4.8. Transmission Electron Microscopy (TEM)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Pinero, F.; Dirchwolf, M.; Pessoa, M.G. Biomarkers in Hepatocellular Carcinoma: Diagnosis, Prognosis and Treatment Response Assessment. Cells 2020, 9, 1370. [Google Scholar] [CrossRef]
- Konyn, P.; Ahmed, A.; Kim, D. Current epidemiology in hepatocellular carcinoma. Expert Rev. Gastroenterol. Hepatol. 2021, 15, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.W.; Li, D.D.; Chen, X.; Li, X.L.; He, Y.P.; Guo, L.H.; Liu, L.N.; Sun, L.P.; Zhang, X.P. Correction to: MicroRNA-125b reverses oxaliplatin resistance in hepatocellular carcinoma by negatively regulating EVA1A mediated autophagy. Cell Death Dis. 2018, 9, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Mei, J.; Wang, Q.; Shi, F.; Liu, H.; Zhao, M.; Lu, L.; Ling, Y.; Guo, Z.; Guo, Y.; et al. Transarterial infusion chemotherapy with FOLFOX for advanced hepatocellular carcinoma: A multi-center propensity score matched analysis of real-world practice. Hepatobiliary Surg. Nutr. 2021, 10, 631–645. [Google Scholar] [CrossRef]
- Goyal, L.; Zheng, H.; Abrams, T.A.; Miksad, R.; Bullock, A.J.; Allen, J.N.; Yurgelun, M.B.; Clark, J.W.; Kambadakone, A.; Muzikansky, A.; et al. A Phase II and Biomarker Study of Sorafenib Combined with Modified FOLFOX in Patients with Advanced Hepatocellular Carcinoma. Clin. Cancer Res. 2019, 25, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Croce, C.M. MicroRNA and ER stress in cancer. Semin. Cancer Biol. 2021, 75, 3–14. [Google Scholar] [CrossRef]
- Li, Y.; Lv, Z.; He, G.; Wang, J.; Zhang, X.; Lu, G.; Ren, X.; Wang, F.; Zhu, X.; Ding, Y.; et al. The SOX17/miR-371-5p/SOX2 axis inhibits EMT, stem cell properties and metastasis in colorectal cancer. Oncotarget 2015, 6, 9099–9112. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Su, F.; Xie, R.; Tang, W. miR-371-5p suppresses the proliferative and migratory capacity of human nasopharyngeal carcinoma by targeting BCL2. Oncol. Lett. 2018, 15, 9209–9215. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Miao, H.; Xu, Y.; Xiong, L.; Wang, Y.; Xiang, H.; Zhang, H.; Zhang, Z. MiR-371-5p facilitates pancreatic cancer cell proliferation and decreases patient survival. PLoS ONE 2014, 9, e112930. [Google Scholar] [CrossRef]
- Liu, R.Y.; Diao, C.F.; Zhang, Y.; Wu, N.; Wan, H.Y.; Nong, X.Y.; Liu, M.; Tang, H. miR-371-5p down-regulates pre mRNA processing factor 4 homolog B (PRPF4B) and facilitates the G1/S transition in human hepatocellular carcinoma cells. Cancer Lett. 2013, 335, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Visalli, M.; Bartolotta, M.; Polito, F.; Oteri, R.; Barbera, A.; Arrigo, R.; Di Giorgio, R.M.; Navarra, G.; Aguennouz, M. miRNA expression profiling regulates necroptotic cell death in hepatocellular carcinoma. Int. J. Oncol. 2018, 53, 771–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wen, L.; Chen, S.; Zhang, J.; Ma, Y.; Hu, J.; Yue, T.; Wang, J.; Zhu, J.; Bu, D.; et al. The novel long noncoding RNA CRART16 confers cetuximab resistance in colorectal cancer cells by enhancing ERBB3 expression via miR-371a-5p. Cancer Cell Int. 2020, 20, 68. [Google Scholar] [CrossRef] [Green Version]
- Gungormez, C.; Teker, E.; Atmanoglu, S.; Borazan, E. Correction to: miRNA Profile and Bioinformatic Analysis for Diagnosis in Patients with Stage IIIA Colon Cancer. Biochem. Genet. 2022, 60, 452. [Google Scholar] [CrossRef] [PubMed]
- Ernst, S.; Heinzelmann, J.; Bohle, R.M.; Weber, G.; Stockle, M.; Junker, K.; Heinzelbecker, J. The metastatic potential of seminomatous germ cell tumours is associated with a specific microRNA pattern. Andrology 2020, 8, 1687–1698. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in major human diseases. EMBO J. 2021, 40, e108863. [Google Scholar] [CrossRef]
- Yazdani, H.O.; Huang, H.; Tsung, A. Autophagy: Dual Response in the Development of Hepatocellular Carcinoma. Cells 2019, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Liu, C.; Yang, Q.; Xin, C.; Du, J.; Sun, F.; Zhou, L. MIR145-3p promotes autophagy and enhances bortezomib sensitivity in multiple myeloma by targeting HDAC4. Autophagy 2020, 16, 683–697. [Google Scholar] [CrossRef]
- Li, H.; Chen, L.; Li, J.J.; Zhou, Q.; Huang, A.; Liu, W.W.; Wang, K.; Gao, L.; Qi, S.T.; Lu, Y.T. miR-519a enhances chemosensitivity and promotes autophagy in glioblastoma by targeting STAT3/Bcl2 signaling pathway. J. Hematol. Oncol. 2018, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.P.; Liu, J.P.; Feng, J.F.; Zhu, C.P.; Yang, Y.; Zhou, W.P.; Ding, J.; Huang, C.K.; Cui, Y.L.; Ding, C.H.; et al. miR-541 potentiates the response of human hepatocellular carcinoma to sorafenib treatment by inhibiting autophagy. Gut 2020, 69, 1309–1321. [Google Scholar] [CrossRef]
- Wu, W.; Lu, P.; Huang, Y.; Zhu, Z.; Li, C.; Liu, Y. Emodin regulates the autophagy via the miR-371a-5p/PTEN axis to inhibit hepatic malignancy. Biochem. Biophys. Res. Commun. 2022, 619, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xu, A.; Kang, L.; Cong, R.; Fan, Z.; Zhu, X.; Huo, N.; Liu, W.; Xue, C.; Ji, Q.; et al. LSD1-Demethylated LINC01134 Confers Oxaliplatin Resistance Through SP1-Induced p62 Transcription in HCC. Hepatology 2021, 74, 3213–3234. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.S.; Hou, P.; Kong, Y. Hepatitis B virus promotes proliferation and metastasis in male Chinese hepatocellular carcinoma patients through the LEF-1/miR-371a-5p/SRCIN1/pleiotrophin/Slug pathway. Exp. Cell Res. 2018, 370, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Bronckaers, A.; Gago, F.; Balzarini, J.; Liekens, S. The dual role of thymidine phosphorylase in cancer development and chemotherapy. Med. Res. Rev. 2009, 29, 903–953. [Google Scholar] [CrossRef]
- Hawkins, D.S.; Demers, G.W.; Galloway, D.A. Inactivation of p53 enhances sensitivity to multiple chemotherapeutic agents. Cancer Res. 1996, 56, 892–898. [Google Scholar]
- Kreso, A.; O’Brien, C.A.; van Galen, P.; Gan, O.I.; Notta, F.; Brown, A.M.; Ng, K.; Ma, J.; Wienholds, E.; Dunant, C.; et al. Variable clonal repopulation dynamics influence chemotherapy response in colorectal cancer. Science 2013, 339, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Vera-Ramirez, L.; Vodnala, S.K.; Nini, R.; Hunter, K.W.; Green, J.E. Autophagy promotes the survival of dormant breast cancer cells and metastatic tumour recurrence. Nat. Commun. 2018, 9, 1944. [Google Scholar] [CrossRef] [Green Version]
- Redshaw, Z.; Sweetman, D.; Loughna, P.T. The effects of age upon the expression of three miRNAs in muscle stem cells isolated from two different porcine skeletal muscles. Differentiation 2014, 88, 117–123. [Google Scholar] [CrossRef]
- Meder, B.; Backes, C.; Haas, J.; Leidinger, P.; Stahler, C.; Grossmann, T.; Vogel, B.; Frese, K.; Giannitsis, E.; Katus, H.A.; et al. Influence of the confounding factors age and sex on microRNA profiles from peripheral blood. Clin. Chem. 2014, 60, 1200–1208. [Google Scholar] [CrossRef]
- Fafian-Labora, J.; Lesende-Rodriguez, I.; Fernandez-Pernas, P.; Sangiao-Alvarellos, S.; Monserrat, L.; Arntz, O.J.; van de Loo, F.J.; Mateos, J.; Arufe, M.C. Effect of age on pro-inflammatory miRNAs contained in mesenchymal stem cell-derived extracellular vesicles. Sci. Rep. 2017, 7, 43923. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.L.; Chatterjee, N.; Chittur, S.V.; Welsh, J.; Tenniswood, M.P. Effects of 1alpha,25 dihydroxyvitamin D3 and testosterone on miRNA and mRNA expression in LNCaP cells. Mol. Cancer 2011, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, K.; Liu, X.; Liu, H.T.; Liang, X.M.; Liang, G.L.; Tang, S.T.; Huo, R.R.; Ma, L.; Xiang, B.B.; et al. Analysis of Clinicopathological Characteristics and Prognosis of Young Patients with Hepatocellular Carcinoma after Hepatectomy. J. Clin. Transl. Hepatol. 2020, 8, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Namieno, T.; Kawata, A.; Sato, N.; Kondo, Y.; Uchino, J. Age-related, different clinicopathologic features of hepatocellular carcinoma patients. Ann. Surg. 1995, 221, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.H.; Ding, Z.B.; Zhou, J.; Qiu, S.J.; Fan, J. Prognostic significance of Beclin 1-dependent apoptotic activity in hepatocellular carcinoma. Autophagy 2009, 5, 380–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z.B.; Shi, Y.H.; Zhou, J.; Qiu, S.J.; Xu, Y.; Dai, Z.; Shi, G.M.; Wang, X.Y.; Ke, A.W.; Wu, B.; et al. Association of autophagy defect with a malignant phenotype and poor prognosis of hepatocellular carcinoma. Cancer Res. 2008, 68, 9167–9175. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.M.; Wang, G.L.; Chen, L.; Xu, Y.Y.; He, S.; Cao, X.L.; Qin, J.; Zhou, J.M.; Zhang, Y.X.; E, Q. The expression of beclin-1, an autophagic gene, in hepatocellular carcinoma associated with clinical pathological and prognostic significance. BMC Cancer 2014, 14, 327. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Yao, Y.; Xu, G.; Zhou, C.; Zhang, Y.; Sun, J.; Jiang, R.; Shao, Q.; Chen, Y. CD24 regulates sorafenib resistance via activating autophagy in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 646. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.; Wei, J.; You, W.; Wang, R.; Shang, J.; Xiong, Y.; Yang, H.; Yang, X.; Fu, Z. Disruption of the c-Myc/miR-200b-3p/PRDX2 regulatory loop enhances tumor metastasis and chemotherapeutic resistance in colorectal cancer. J. Transl. Med. 2017, 15, 257. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Events | n | miR-371a-5p in HCC Tissues (Median) a | U/H Value | p Value | miR-371a-5p in HCC Serum (Median) b | U/H Value | p Value |
|---|---|---|---|---|---|---|---|
| Gender | 237 | 0.434 | 269 | 0.842 | |||
| Male | 51 | 0.356 | 1.210 | ||||
| female | 11 | 0.376 | 0.757 | ||||
| Age | 180 | 0.004 | 151 | 0.001 | |||
| ≤50 | 15 | 0.934 | 5.580 | ||||
| >50 | 47 | 0.253 | 0.616 | ||||
| HBsAg | 415 | 0.561 | 368 | 0.208 | |||
| Negative | 24 | 0.229 | 0.261 | ||||
| Positive | 38 | 0.383 | 1.270 | ||||
| Serum AFP (μg/L) | 248 | 0.001 | 391 | 0.221 | |||
| ≤400 | 33 | 0.109 | 0.697 | ||||
| >400 | 29 | 0.797 | 1.370 | ||||
| Liver cirrhosis | 459 | 0.855 | 461 | 0.877 | |||
| Absence | 35 | 0.356 | 1.000 | ||||
| Presence | 27 | 0.376 | 0.964 | ||||
| Tumor size (cm) | 239 | 0.023 | 122 | < 0.001 | |||
| ≤5.0 | 45 | 0.347 | 0.616 | ||||
| >5.0 | 17 | 0.455 | 5.580 | ||||
| Tumor number | 338 | 0.820 | 256 | 0.115 | |||
| Single | 47 | 0.361 | 0.723 | ||||
| Multiple | 15 | 0.356 | 2.790 | ||||
| Tumor differentiation (grading) | 9.45 | 0.009 | 13.7 | 0.001 | |||
| Well (1) | 11 | 0.347 | 0.174 | ||||
| Moderate (2) | 30 | 0.117 | 0.246 | ||||
| Poor (3) | 21 | 1.070 * | 2.79 # * | ||||
| Microvascular invasion | 374 | 0.607 | 322 | 0.191 | |||
| Yes | 19 | 0.356 | 2.600 | ||||
| No | 43 | 0.376 | 0.838 |
| Events | n | HCC BECN1 IHC Scores (Mean ± SD) | F/t Value | p Value |
|---|---|---|---|---|
| PNCT | 62 | 2.560 ± 0.617 | 12.30 | <0.001 |
| HCC | 1.100 ± 0.762 | |||
| Gender | 1.100 | 0.278 | ||
| Male | 51 | 1.180 ± 0.740 | ||
| female | 11 | 0.909 ± 0.701 | ||
| Age | 0.026 | 0.978 | ||
| ≤50 | 15 | 1.130 ± 0.915 | ||
| >50 | 47 | 1.130 ± 0.679 | ||
| HBsAg | 0.386 | 0.701 | ||
| Negative | 24 | 1.080 ± 0.654 | ||
| Positive | 38 | 1.160 ± 0.789 | ||
| Serum AFP (μg/L) | 0.949 | 0.347 | ||
| ≤400 | 33 | 1.210 ± 0.650 | ||
| >400 | 29 | 1.030 ± 0.823 | ||
| Liver cirrhosis | 0.514 | 0.609 | ||
| Absence | 35 | 1.170 ± 0.747 | ||
| Presence | 27 | 1.070 ± 0.730 | ||
| Tumor size (cm) | 2.060 | 0.043 | ||
| ≤5.0 | 45 | 1.240 ± 0.743 | ||
| >5.0 | 17 | 0.824 ± 0.636 | ||
| Tumor number | 0.026 | 0.980 | ||
| Single | 47 | 1.130 ± 0.769 | ||
| Multiple | 15 | 1.130 ± 0.640 | ||
| Tumor differentiation (grading) | 29.10 | <0.001 | ||
| Well (1) | 11 | 2.000 ± 0.632 | ||
| Moderate (2) | 30 | 1.230 ± 0.430 | ||
| Poor (3) | 21 | 0.524 ± 0.602 # * | ||
| Microvascular invasion | 0.541 | 0.591 | ||
| Yes | 19 | 1.050 ± 0.705 | ||
| No | 43 | 1.160 ± 0.754 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Qiu, X.; Jin, P.; Li, Z.; Zhang, Y.; Lv, L.; Song, F. MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy. Life 2022, 12, 1651. https://doi.org/10.3390/life12101651
Lv Z, Qiu X, Jin P, Li Z, Zhang Y, Lv L, Song F. MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy. Life. 2022; 12(10):1651. https://doi.org/10.3390/life12101651
Chicago/Turabian StyleLv, Zhenbing, Xin Qiu, Pu Jin, Zhaodong Li, Yan Zhang, Lei Lv, and Fangzhou Song. 2022. "MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy" Life 12, no. 10: 1651. https://doi.org/10.3390/life12101651
APA StyleLv, Z., Qiu, X., Jin, P., Li, Z., Zhang, Y., Lv, L., & Song, F. (2022). MiR-371a-5p Positively Associates with Hepatocellular Carcinoma Malignancy but Sensitizes Cancer Cells to Oxaliplatin by Suppressing BECN1-Dependent Autophagy. Life, 12(10), 1651. https://doi.org/10.3390/life12101651