
Collagenase-Induced Mouse Model of Osteoarthritis—A Thorough Flow Cytometry Analysis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Antibodies
2.3. Induction of Osteoarthritis
2.4. Isolation of Cells from Spleen and Synovium
2.5. Synovial Extracts
2.6. Isolation of Cells from Bone Marrow (BM)
2.7. Flow Cytometry Analysis
2.8. Histological Analysis
2.9. Statistical Analysis
3. Results
3.1. Histopathological Analysis of the Experimental Mouse Model of OA during the 7th, 14th, and 30th Day of Development
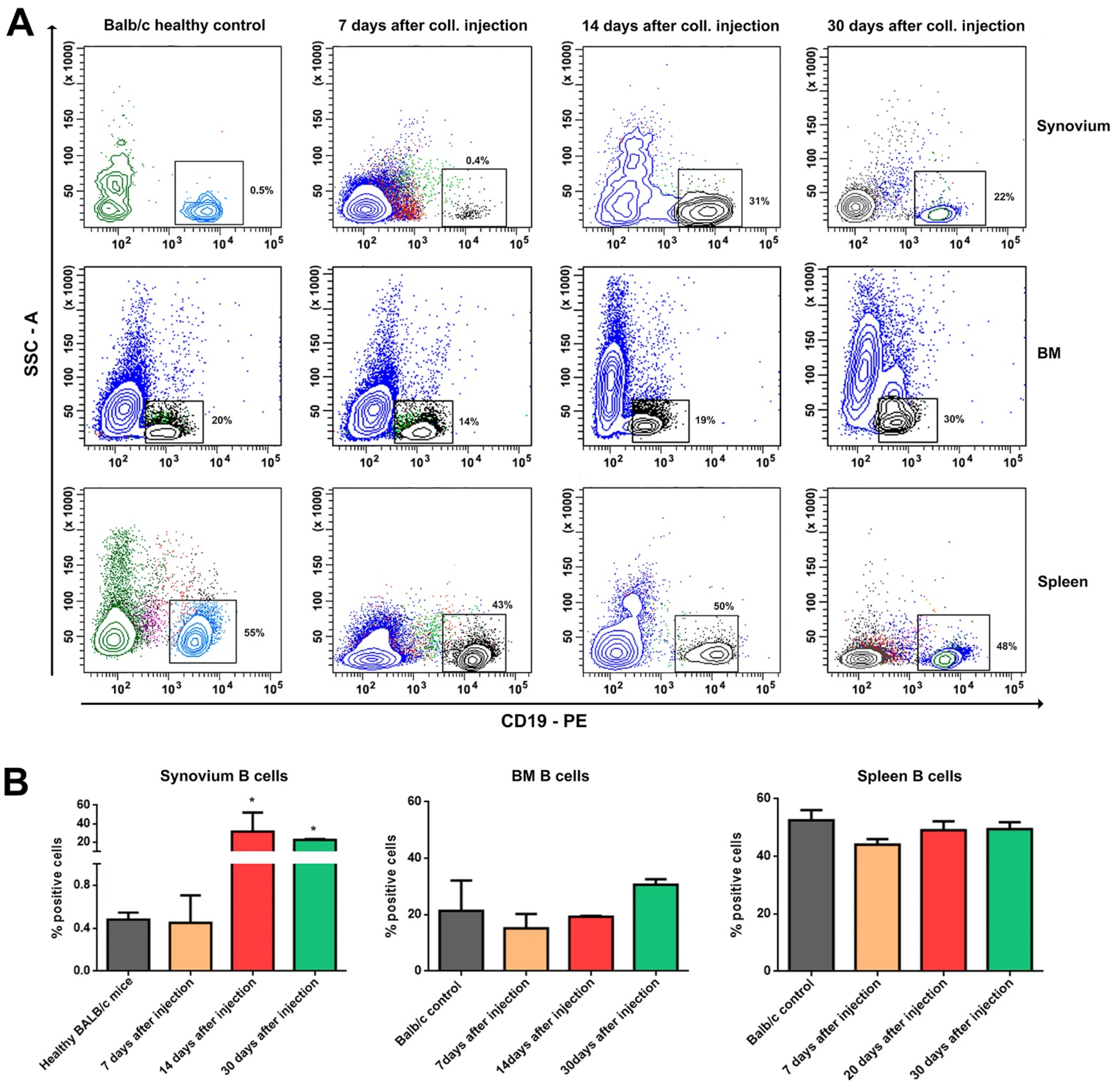
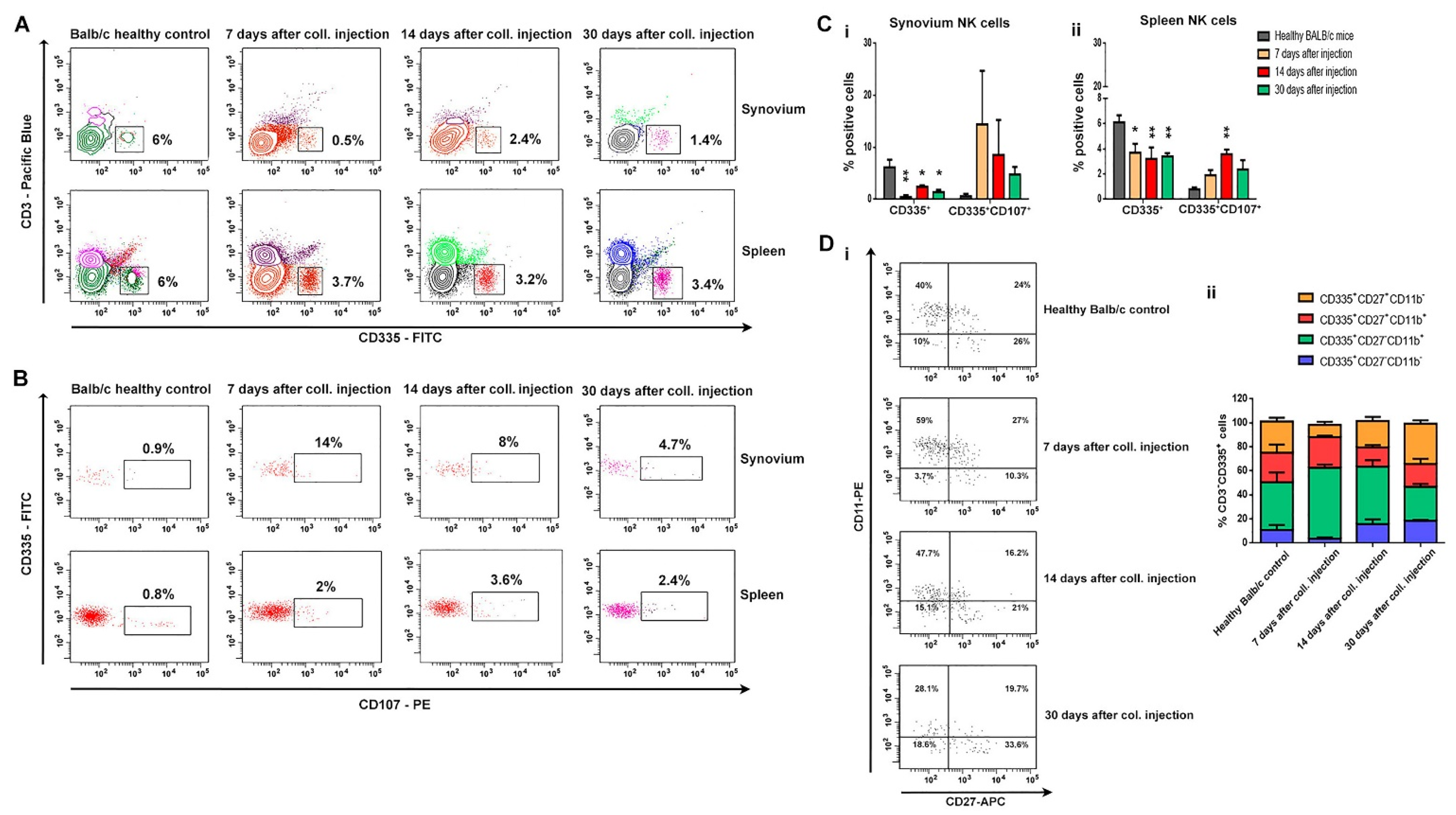
3.2. Distribution of B, T, and NK Cells in the Synovium, BM, and Spleen during the Experimental OA Development
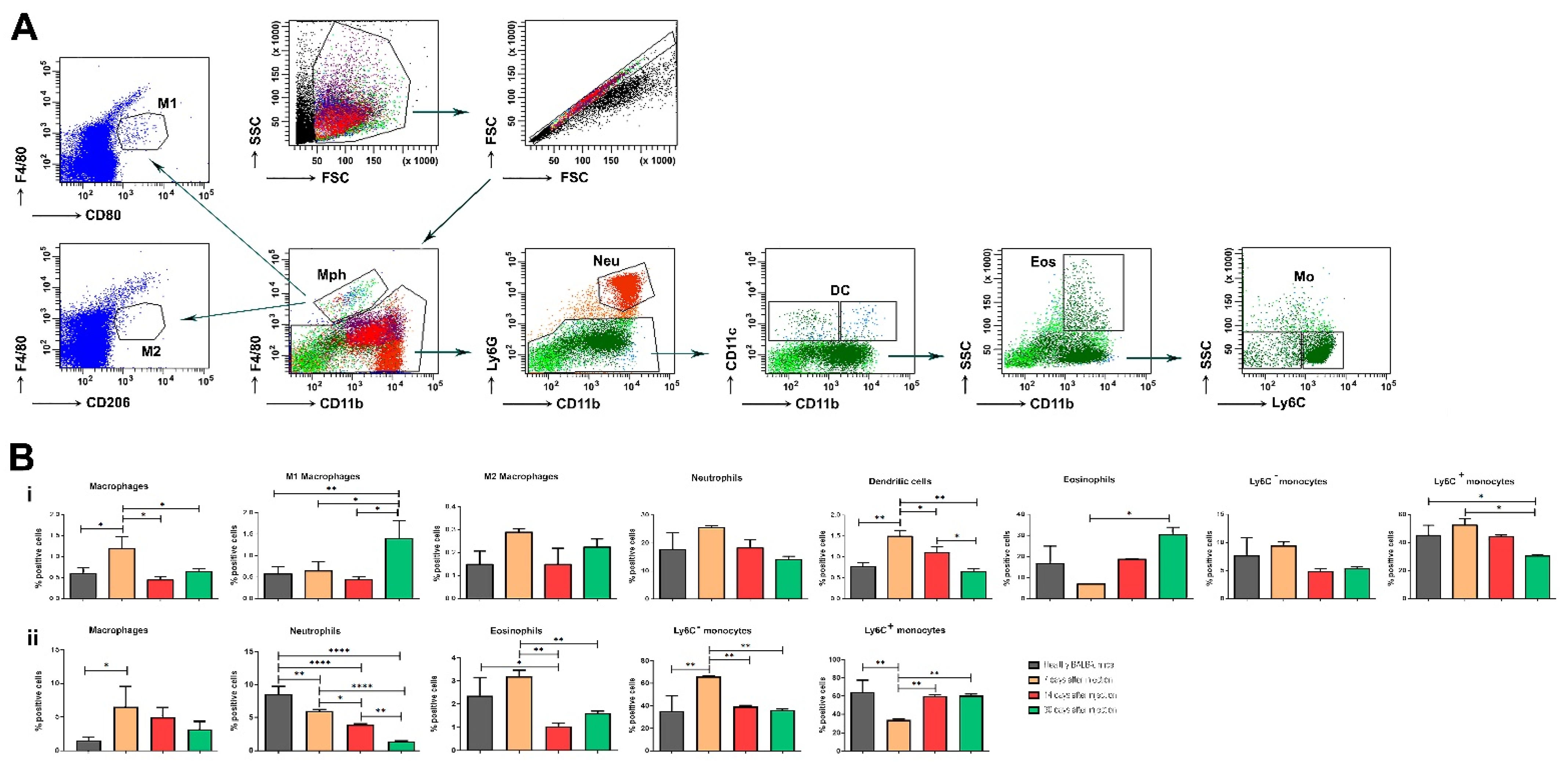
3.3. Distribution of Myeloid Cells in the Synovium and BM during OA Development
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adães, S.; Mendonça, M.; Santos, T.N.; Castro-Lopes, J.M.; Ferreira-Gomes, J.; Neto, F.L. Intra-articular injection of collagenase in the knee of rats as an alternative model to study nociception associated with osteoarthritis. Arthritis Res. Ther. 2014, 16, R10. [Google Scholar] [CrossRef] [Green Version]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]
- Benigni, G.; Dimitrova, P.; Antonangeli, F.; Sanseviero, E.; Milanova, V.; Blom, A.; van Lent, P.; Morrone, S.; Satoni, A.; Bernardini, G. CXCR3/CXCL10 Axis Regulates Neutrophil–NK Cell Cross-Talk Determining the Severity of Experimental Osteoarthritis. J. Immunol. 2017, 198, 2115–2124. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieza, A.; Causey, K.; Kamenov, K.; Hanson, S.; Chatterji, S.; Vos, T. Global estimates of the need for rehabilitation based on the Global Burden of Disease study 2019: A systematic analysis for the Global Burden of Disease Study 2019. The Lancet 2020, 396, 2006–2017. [Google Scholar] [CrossRef]
- Cope, P.J.; Ourradi, K.; Li, Y.; Sharif, M. Models of osteoarthritis: The good, the bad and the promising. Osteoarthr. Cartil. 2019, 27, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culemann, S.; Grüneboom, A.; Krönke, G. Chapter Three—Origin and function of synovial macrophage subsets during inflammatory joint disease. Adv. Immunol. 2019, 143, 75–98. [Google Scholar] [PubMed]
- Da, R.-R.; Qin, Y.; Baeten, D.; Zhang, Y. B Cell Clonal Expansion and Somatic Hypermutation of Ig Variable Heavy Chain Genes in the Synovial Membrane of Patients with Osteoarthritis. J. Immunol. 2007, 178, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Dalbeth, N.; Callan, M.F. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2002, 46, 1763–1772. [Google Scholar] [CrossRef]
- Dalbeth, N.; Gundle, R.; Davies, R.J.; Lee, Y.G.; McMichael, A.J.; Callan, M.F. CD56bright NK cells are enriched at inflammatory sites and can engage with monocytes in a reciprocal program of activation. J. Immunol. 2004, 173, 6418–6426. [Google Scholar] [CrossRef]
- Du, J.; Chen, S.; Shi, J.; Zhu, X.; Ying, H.; Zhang, Y.; Chen, S.; Shen, B.; Li, J. The association between the lymphocyte-monocyte ratio and disease activity in rheumatoid arthritis. Clin. Rheumatol. 2017, 36, 2689–2695. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Zhu, W.; Liu, W.; Ma, D.; Li, H.; Yu, W.; Wang, L.; Cao, Y.; Jiang, Y. Diagnostic value of the blood monocyte–lymphocyte ratio in knee osteoarthritis. J. Int. Med. Res. 2019, 47, 4413–4421. [Google Scholar] [CrossRef] [PubMed]
- Guerassimov, A.; Zhang, Y.; Cartman, A.; Rosenberg, L.C.; Esdaile, J.; Fitzcharles, M.-A.; Poole, A.R. Immune responses to cartilage link protein and the G1 domain of proteoglycan aggrecan in patients with osteoarthritis. Arthritis Rheum. 1999, 42, 527–533. [Google Scholar] [CrossRef]
- Haseeb, A.; Haqqi, T.M. Immunopathogenesis of osteoarthritis. Clin. Immunol. 2013, 146, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Haubruck, P.; Pinto, M.M.; Moradi, B.; Little, C.B.; Gentek, R. Monocytes, Macrophages, and Their Potential Niches in Synovial Joints—Therapeutic Targets in Post-Traumatic Osteoarthritis? Front. Immunol. 2021, 12, 763702. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, M.-F.; Zhang, X.; Wellman, S.S.; Bolognesi, M.P.; Kraus, V.B. Synergistic Roles of Macrophages and Neutrophils in Osteoarthritis Progression. Arthritis Rheumatol. 2021, 73, 89–99. [Google Scholar] [CrossRef]
- Huang, Y.; Deng, W.; Zheng, S.; Feng, F.; Huang, Z.; Huang, Q.; Guo, X.; Huang, Z.; Huang, X.; Pan, X.; et al. Relationship between monocytes to lymphocytes ratio and axial spondyloarthritis. Int. Immunopharmacol. 2018, 57, 43–46. [Google Scholar] [CrossRef]
- Hussein, M.R.; Fathi, N.A.; El-Din, A.M.E.; Hassan, H.I.; Abdullah, F.; Al-Hakeem, E. Alterations of the CD4+, CD8+ T Cell Subsets, Interleukins-1β, IL-10, IL-17, Tumor Necrosis Factor-α and Soluble Intercellular Adhesion Molecule-1 in Rheumatoid Arthritis and Osteoarthritis: Preliminary Observations. Pathol. Oncol. Res. 2008, 14, 321–328. [Google Scholar] [CrossRef]
- Jaime, P.; García-Guerrero, N.; Estella, R.; Pardo, J.; García-Álvarez, F.; Martinez-Lostao, L. CD56+/CD16− Natural Killer cells expressing the inflammatory protease granzyme A are enriched in synovial fluid from patients with osteoarthritis. Osteoarthr. Cartil. 2017, 25, 1708–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampropoulou-Adamidou, K.; Lelovas, P.; Karadimas, E.V.; Liakou, C.; Triantafillopoulos, I.K.; Dontas, I.; Papaioannou, N.A. Useful animal models for the research of osteoarthritis. Eur. J. Orthop. Surg. Traumatol. 2014, 24, 263–271. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, J.; Grässel, S. Experimental Osteoarthritis Models in Mice. In Mouse Genetics; Springer: Berlin/Heidelberg, Germany, 2014; pp. 401–419. [Google Scholar]
- Nees, T.A.; Rosshirt, N.; Zhang, J.A.; Platzer, H.; Sorbi, R.; Tripel, E.; Reiner, T.; Walker, T.; Schiltenwolf, M.; Lorenz, H.-M.; et al. T Helper Cell Infiltration in Osteoarthritis-Related Knee Pain and Disability. J. Clin. Med. 2020, 9, 2423. [Google Scholar] [CrossRef] [PubMed]
- Orlowsky, E.W.; Kraus, V.B. The Role of Innate Immunity in Osteoarthritis: When Our First Line of Defense Goes On the Offensive. J. Rheumatol. 2015, 42, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Poulet, B. Non-invasive loading model of murine osteoarthritis. Curr. Rheumatol. Rep. 2016, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Jin, H.-Z.; Li, Y.-J.; Chen, Z. Emerging Role of Eosinophils in Resolution of Arthritis. Front. Immunol. 2021, 12, 4260. [Google Scholar] [CrossRef] [PubMed]
- Rosshirt, N.; Trauth, R.; Platzer, H.; Tripel, E.; Nees, T.A.; Lorenz, H.-M.; Tretter, T.; Moradi, B. Proinflammatory T cell polarization is already present in patients with early knee osteoarthritis. Arthritis Res. Ther. 2021, 23, 37. [Google Scholar] [CrossRef]
- Sargin, G.; Senturk, T.; Yavasoglu, I.; Kose, R. Relationship between neutrophil-lymphocyte, platelet-lymphocyte ratio and disease activity in rheumatoid arthritis treated with rituximab. Int. J. Rheum. Dis. 2018, 21, 2122–2127. [Google Scholar] [CrossRef]
- Smith, M.D.; Triantafillou, S.; Parker, A.; Youssef, P.P.; Coleman, M. Synovial membrane inflammation and cytokine production in patients with early osteoarthritis. J. Rheumatol. 1997, 24, 365–371. [Google Scholar]
- Tak, P.P.; Kummer, J.A.; Hack, C.E.; Daha, M.R.; Smeets, T.J.; Erkelens, G.W.; Meinders, A.E.; Kluin, P.M.; Breedveld, F.C. Granzyme-positive cytotoxic cells are specifically increased in early rheumatoid synovial tissue. Arthritis Rheum. 1994, 37, 1735–1743. [Google Scholar] [CrossRef]
- van de Loo, F.A.; Arntz, O.J.; van Enckevort, F.H.; van Lent, P.L.; van den Berg, W.B. Reduced cartilage proteoglycan loss during zymosan-induced gonarthritis in NOS2-deficient mice and in anti-interleukin-1-treated wild-type mice with unabated joint inflammation. Arthritis Rheum. 1998, 41, 634–646. [Google Scholar] [CrossRef]
- van der Kraan, P.M.; Vitters, E.L.; van Beuningen, H.M.; van de Putte, L.B.; van den Berg, W.B. Degenerative knee joint lesions in mice after a single intra-articular collagenase injection. A new model of osteoarthritis. J. Exp. Pathol. 1990, 71, 19–31. [Google Scholar]
- van Osch, G.J.V.M.; van der Kraan, P.M.; Vitters, E.L.; Blankevoort, L.; van den Berg, W.B. Induction of osteoarthritis by intra-articular injection of collagenase in mice. Strain and sex related differences. Osteoarthr. Cartil. 1993, 1, 171–177. [Google Scholar] [CrossRef]
- Wang, E.C.; Newton, Z.; Hayward, O.A.; Clark, S.R.; Collins, F.; Perks, W.V.; Singh, R.K.; Twohig, J.P.; Williams, A.S. Regulation of early cartilage destruction in inflammatory arthritis by death receptor 3. Arthritis Rheumatol. 2014, 66, 2762–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildbaum, G.; Netzer, N.; Karin, N. Plasmid DNA encoding IFN-γ-inducible protein 10 redirects antigen-specific T cell polarization and suppresses experimental autoimmune encephalomyelitis. J. Immunol. 2002, 168, 5885–5892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Zhang, X.; Jiang, Y.; Liu, X.; Huang, L.; Wei, Q.; Huang, Y.; Wu, W.; Gu, J. Alterations in peripheral T cell and B cell subsets in patients with osteoarthritis. Clin. Rheumatol. 2020, 39, 523–532. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boneva, B.; Ralchev, N.; Ganova, P.; Tchorbanov, A.; Mihaylova, N. Collagenase-Induced Mouse Model of Osteoarthritis—A Thorough Flow Cytometry Analysis. Life 2022, 12, 1938. https://doi.org/10.3390/life12111938
Boneva B, Ralchev N, Ganova P, Tchorbanov A, Mihaylova N. Collagenase-Induced Mouse Model of Osteoarthritis—A Thorough Flow Cytometry Analysis. Life. 2022; 12(11):1938. https://doi.org/10.3390/life12111938
Chicago/Turabian StyleBoneva, Blagovesta, Nikola Ralchev, Petya Ganova, Andrey Tchorbanov, and Nikolina Mihaylova. 2022. "Collagenase-Induced Mouse Model of Osteoarthritis—A Thorough Flow Cytometry Analysis" Life 12, no. 11: 1938. https://doi.org/10.3390/life12111938
APA StyleBoneva, B., Ralchev, N., Ganova, P., Tchorbanov, A., & Mihaylova, N. (2022). Collagenase-Induced Mouse Model of Osteoarthritis—A Thorough Flow Cytometry Analysis. Life, 12(11), 1938. https://doi.org/10.3390/life12111938