
Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Description and Experimental Protocol
2.2. Data Analysis
2.3. Thermal Time (TT)
2.4. Hydrotime ()
2.5. Hydrothermal Time Model (HTT)
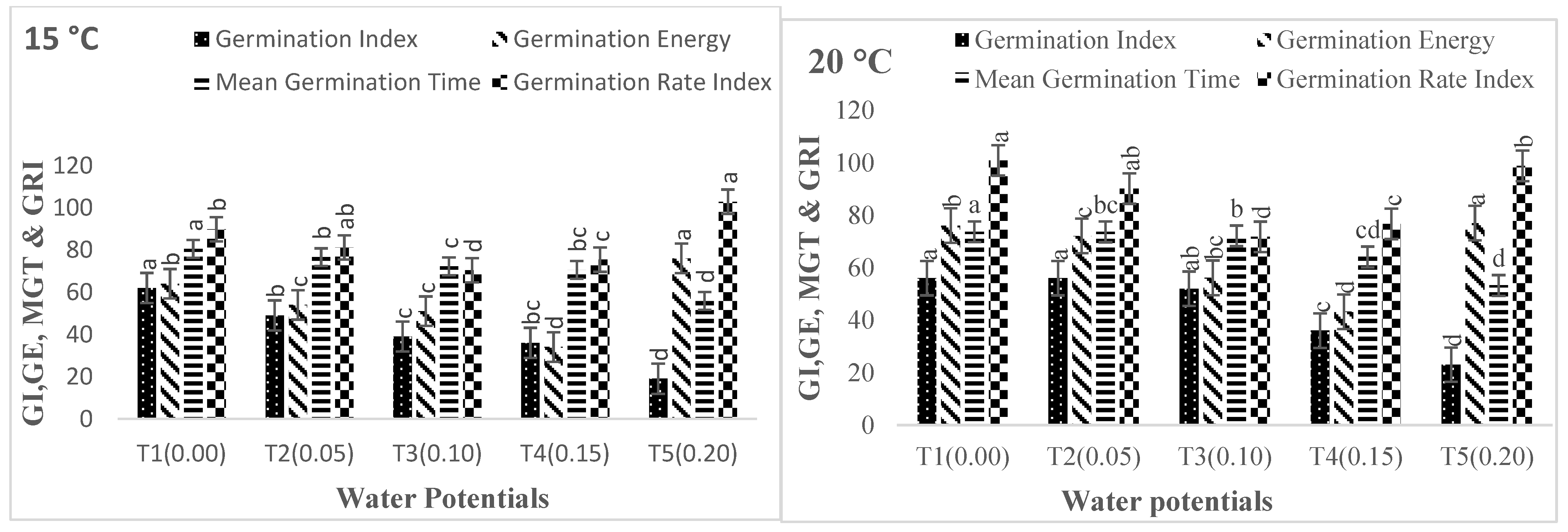
2.6. Germination Attributes
2.6.1. Mean Germination Time (MGT)
2.6.2. Germination Rate Index (GRI)
2.6.3. Germination Index (GI)
2.6.4. Coefficient of Velocity of Germination (CVG)
2.6.5. Germination Energy (GE)
2.6.6. Seed Vigor Index-I (SVI-I)
2.6.7. Seed Vigor Index-II (SVI-II)
2.7. Statistical Analysis
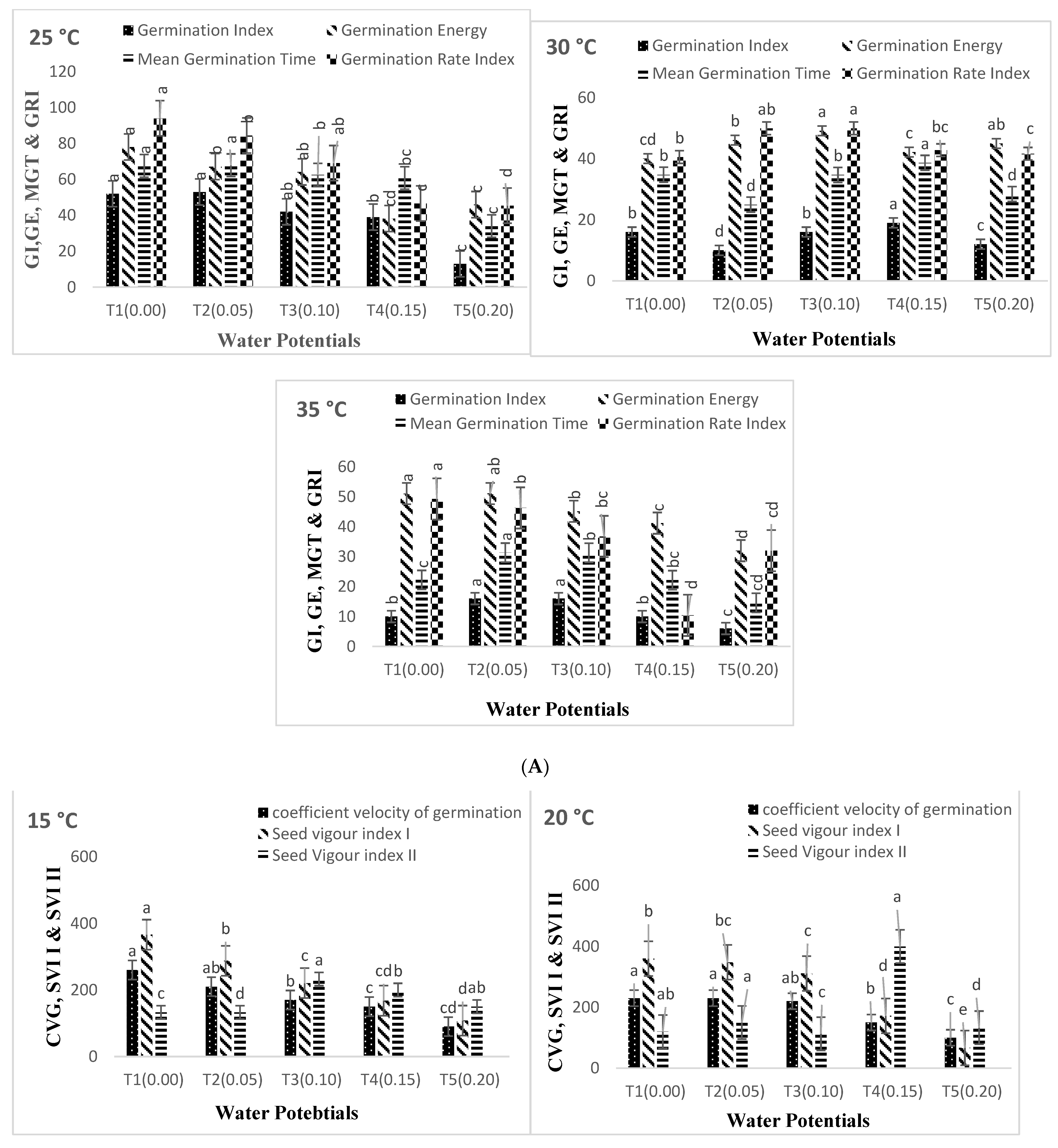
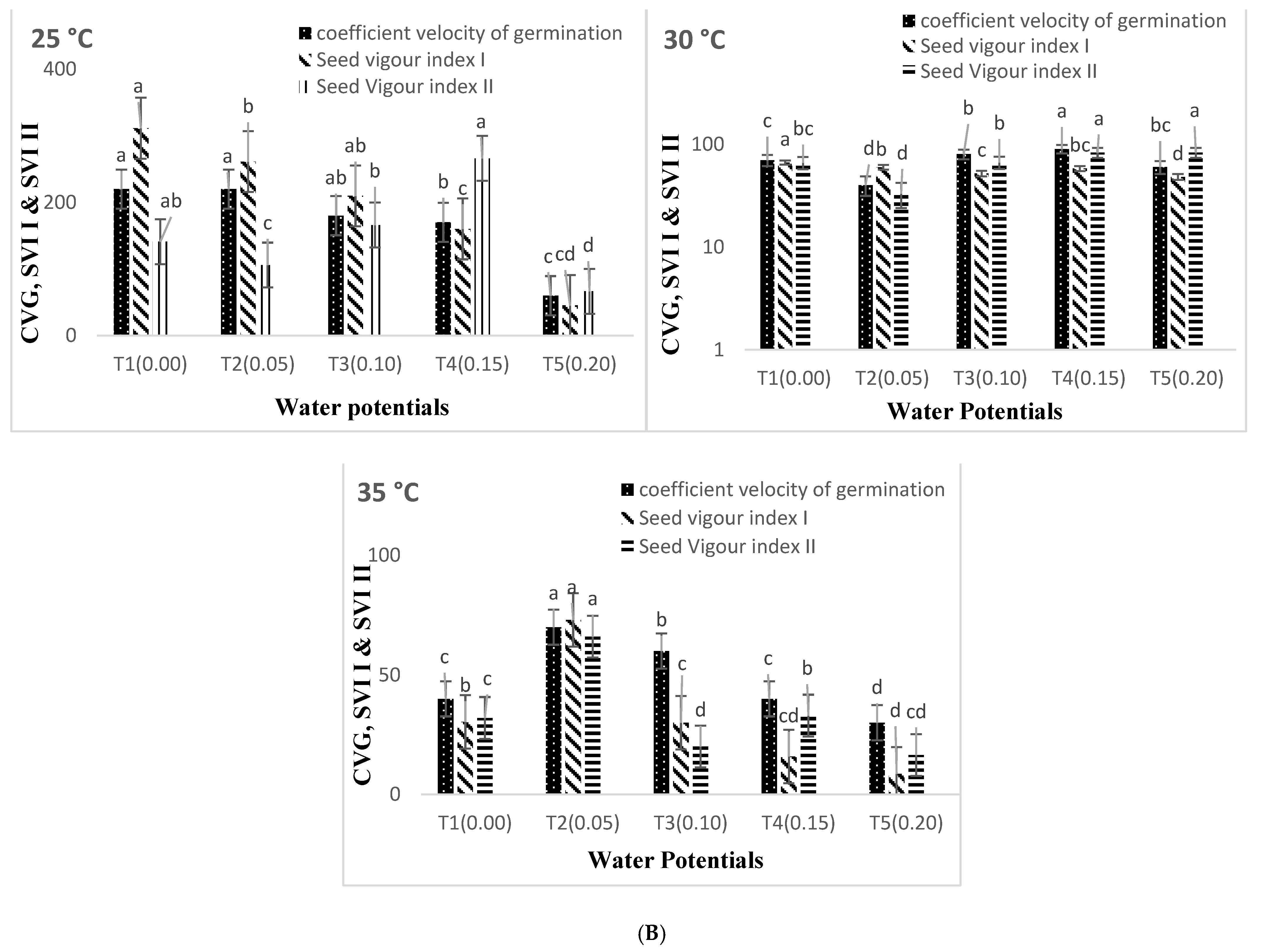
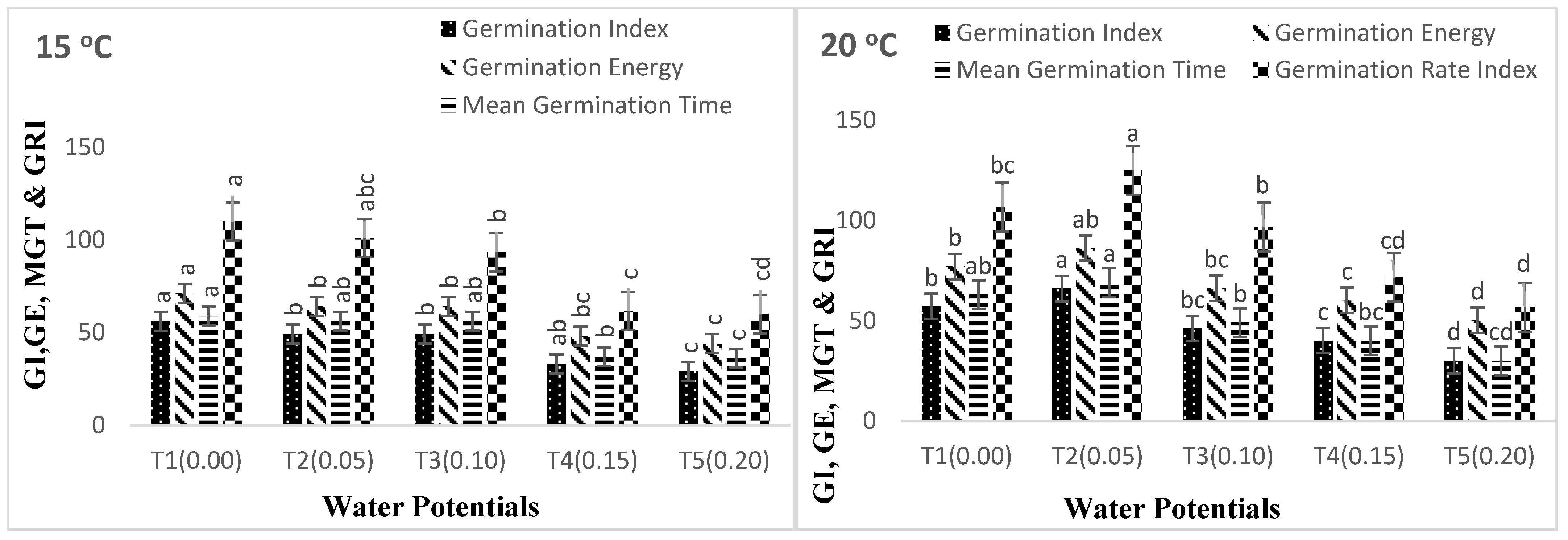
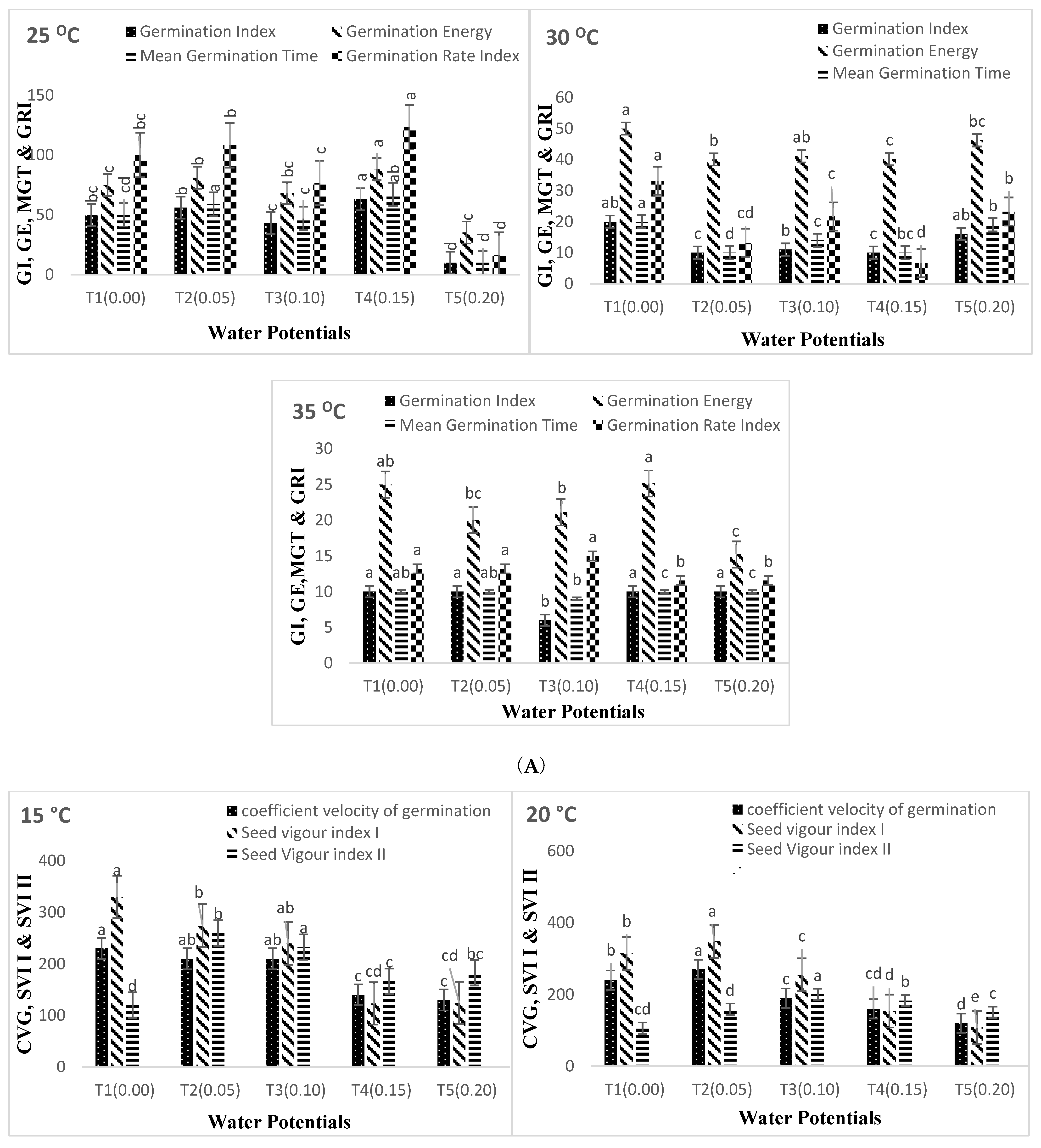
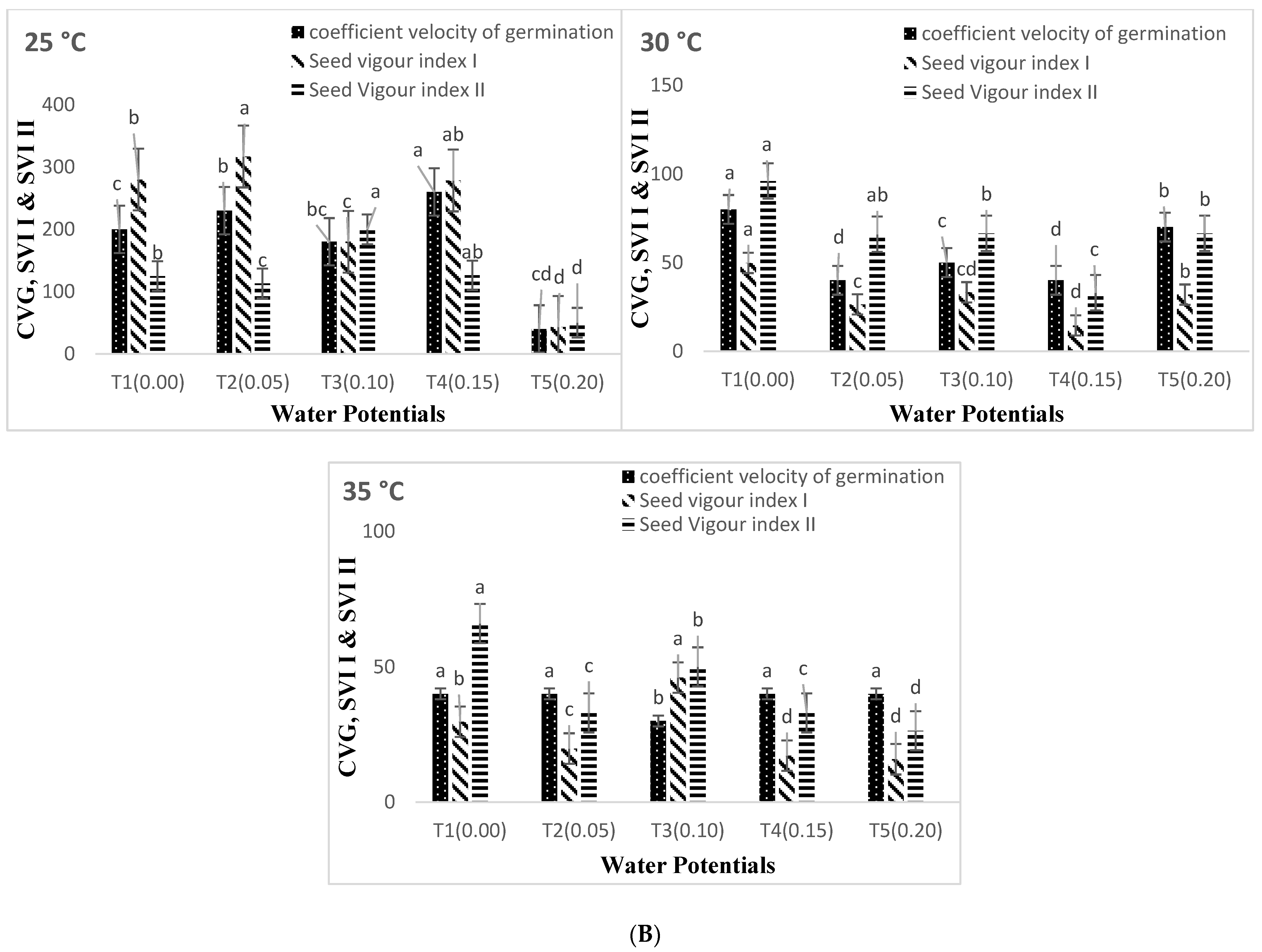
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tack, J.; Barkley, A.; Nalley, L.L. Effect of Warming Temperatures on US Wheat Yields. Proc. Natl. Acad. Sci. USA 2015, 112, 6931–6936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.; Jamshed, M.; Samuel, M.A. Abiotic Stress Signaling in Wheat–an Inclusive Overview of Hormonal Interactions during Abiotic Stress Responses in Wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Zaib-Un-Nisa; Ullah, I.; Amaral Júnior, A.T.; et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A Meta-Analysis of Crop Yield under Climate Change and Adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W. Rising Temperatures Reduce Global Wheat Production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Noor, J.; Ullah, A.; Saleem, M.H.; Tariq, A.; Ullah, S.; Waheed, A.; Okla, M.K.; Al-Hashimi, A.; Chen, Y.; Ahmed, Z. Effect of Jasmonic Acid Foliar Spray on the Morpho-Physiological Mechanism of Salt Stress Tolerance in Two Soybean Varieties (Glycine max L.). Plants 2022, 11, 651. [Google Scholar] [CrossRef]
- Khan, S.; Ullah, A.; Ullah, S.; Saleem, M.H.; Okla, M.K.; Al-Hashimi, A.; Chen, Y.; Ali, S. Quantifying Temperature and Osmotic Stress Impact on Seed Germination Rate and Seedling Growth of Eruca sativa Mill. via Hydrothermal Time Model. Life 2022, 12, 400. [Google Scholar] [CrossRef]
- Ullah, A.; Li, M.; Noor, J.; Tariq, A.; Liu, Y.; Shi, L. Effects of Salinity on Photosynthetic Traits, Ion Homeostasis and Nitrogen Metabolism in Wild and Cultivated Soybean. PeerJ 2019, 7, e8191. [Google Scholar] [CrossRef] [Green Version]
- Blum, A.; Shpiler, L.; Golan, G.; Mayer, J. Yield Stability and Canopy Temperature of Wheat Genotypes under Drought-Stress. Field Crops Res. 1989, 22, 289–296. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 1998; ISBN 0-08-054086-4. [Google Scholar]
- Chen, P.; Jiang, L.; Yang, W.; Wang, L.; Wen, Z. Seed Germination Response and Tolerance to Different Abiotic Stresses of Four Salsola Species Growing in an Arid Environment. Front. Plant Sci. 2022, 13, 892667. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Jamali, M. Halothermal and hydrothermal time models describe germination responses of canola seeds to ageing. Plant Biol. 2021, 23, 621–629. [Google Scholar] [CrossRef]
- Ullah, A.; Sadaf, S.; Ullah, S.; Alshaya, H.; Okla, M.K.; Alwasel, Y.A.; Tariq, A. Using Halothermal Time Model to Describe Barley (Hordeumvulgare L.) Seed Germination Response to Water Potential and Temperature. Life 2022, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Saberali, S.F.; Shirmohamadi-Aliakbarkhani, Z. Quantifying Seed Germination Response of Melon (Cucumis melo L.) to Temperature and Water Potential: Thermal Time, Hydrotime and Hydrothermal Time Models. S. Afr. J. Bot. 2020, 130, 240–249. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Pirdashti, H.; Vahabinia, F.; Gholamhossieni, M. Quantification of the Effect of Environmental Factors on Seed Germination and Seedling Growth of Eruca (Eruca sativa) Using Mathematical Models. J. Plant Growth Regul. 2020, 39, 190–204. [Google Scholar] [CrossRef]
- Gummerson, R.J. The Effect of Constant Temperatures and Osmotic Potentials on the Germination of Sugar Beet. J. Exp. Bot. 1986, 37, 729–741. [Google Scholar] [CrossRef]
- Alvarado, V.; Bradford, K.J. A Hydrothermal Time Model Explains the Cardinal Temperatures for Seed Germination. Plant Cell Environ. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Atashi, S.; Bakhshandeh, E.; Zeinali, Z.; Yassari, E.; Teixeira da Silva, J.A. Modeling Seed Germination in Melisa officinalis L. in Response to Temperature and Water Potential. Acta Physiol. Plant. 2014, 36, 605–611. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Bradford, K.J.; Pirdashti, H.; Vahabinia, F.; Abdellaoui, R. A New Halothermal Time Model Describes Seed Germination Responses to Salinity across Both Sub-and Supra-Optimal Temperatures. Acta Physiol. Plant. 2020, 42, 1–15. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.; Hilhorst, H. Seeds: Physiology of Development, Germination and Dormancy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 1-4614-4692-9. [Google Scholar]
- Parmoon, G.; Moosavi, S.A.; Akbari, H.; Ebadi, A. Quantifying Cardinal Temperatures and Thermal Time Required for Germination of Silybum Marianum Seed. Crop J. 2015, 3, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi-Far, F.; Bakhshandeh, E.; Ghadirian, R. Evaluating Seed Quality in Sesame (Sesamum indicum L.) by the Accelerated Ageing Test. Seed Technol. 2010, 32, 69–72. [Google Scholar]
- Odoba, A.; Odiaka, N.I.; Gbanguba, A.U.; Bashiru, M. Germination Characteristics of Twenty Varieties of Soybean (Glycine max (L.) Merr) Stored for Seven Months. Sci. Agric. 2016, 13, 151–155. [Google Scholar]
- Rowse, H.R.; Finch-Savage, W.E. Hydrothermal Threshold Models Can Describe the Germination Response of Carrot (Daucus carota) and Onion (Allium cepa) Seed Populations across Both Sub-and Supra-optimal Temperatures. New Phytol. 2003, 158, 101–108. [Google Scholar] [CrossRef]
- Tabrizi, L.; Nasiri, M.M.; Kouchaki, A.R. Investigations on the Cardinal Temperatures for Germination of Plantago ovata and Plantago psyllium. Fall Winter 2005, 2, 143–150. [Google Scholar]
- Bidgoly, R.O.; Balouchi, H.; Soltani, E.; Moradi, A. Effect of Temperature and Water Potential on Carthamus tinctorius L. Seed Germination: Quantification of the Cardinal Temperatures and Modeling Using Hydrothermal Time. Ind. Crops Prod. 2018, 113, 121–127. [Google Scholar] [CrossRef]
- Atashi, S.; Bakhshandeh, E.; Mehdipour, M.; Jamali, M.; Teixeira da Silva, J.A. Application of a Hydrothermal Time Seed Germination Model Using the Weibull Distribution to Describe Base Water Potential in Zucchini (Cucurbita pepo L.). J. Plant Growth Regul. 2015, 34, 150–157. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of Hydrothermal Time to Quantifying and Modeling Seed Germination and Dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 1-4899-1002-6. [Google Scholar]
- Kader, M.A. A Comparison of Seed Germination Calculation Formulae and the Associated Interpretation of Resulting Data. J. Proc. R. Soc. Wales 2005, 138, 65–75. [Google Scholar]
- Swaminathan, C.; Revathy, R. Improving Seed Germination in Sapindus Emarginatus Vahl. Pinnacle Agric. Res. Manag. 2013, 2013, 3. [Google Scholar]
- Bradford, K.J.; Still, D.W. Applications of Hydrotime Analysis in Seed Testing. Seed Technol. 2004, 26, 75–85. [Google Scholar]
- Khaliliaqdam, N.; Soltani, A.; Latifi, N.; Ghaderi Far, F. Soybean Seed Aging and Environmental Factors on Seedling Growth. Commun. Soil Sci. Plant Anal. 2013, 44, 1786–1799. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Atashi, S.; Hafeznia, M.; Pirdashti, H.; Teixeira da Silva, J.A. Hydrothermal Time Analysis of Watermelon (Citrullus vulgaris Cv.‘Crimson Sweet’) Seed Germination. Acta Physiol. Plant. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Dahal, P.; Bradford, K.J. Effects of Priming and Endosperm Integrity on Seed Germination Rates of Tomato Genotypes: II. Germination at Reduced Water Potential. J. Exp. Bot. 1990, 41, 1441–1453. [Google Scholar] [CrossRef]
- Wang, R. Modeling Seed Germination and Seedling Emergence in Winterfat (Krascheninnikovia lanata (Pursh) ADJ Meeuse & Smit): Physiological Mechanisms and Ecological Relevance. Ph.D. Thesis, University of Saskatchewan Saskatoon, Saskatoon, SK, Canada, 2005. [Google Scholar]
- Buriro, M.; Oad, F.C.; Keerio, M.I.; Tunio, S.; Gandahi, A.W.; Hassan, S.W.U.; Oad, S.M. Wheat Seed Germination under the Influence of Temperature Regimes. Sarhad J. Agric. 2011, 27, 539–543. [Google Scholar]
- Porter, J.R.; Gawith, M. Temperatures and the Growth and Development of Wheat: A Review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar] [CrossRef]
- Nuttall, J.G.; Barlow, K.M.; Delahunty, A.J.; Christy, B.P.; O’Leary, G.J. Acute High Temperature Response in Wheat. Agron. J. 2018, 110, 1296–1308. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
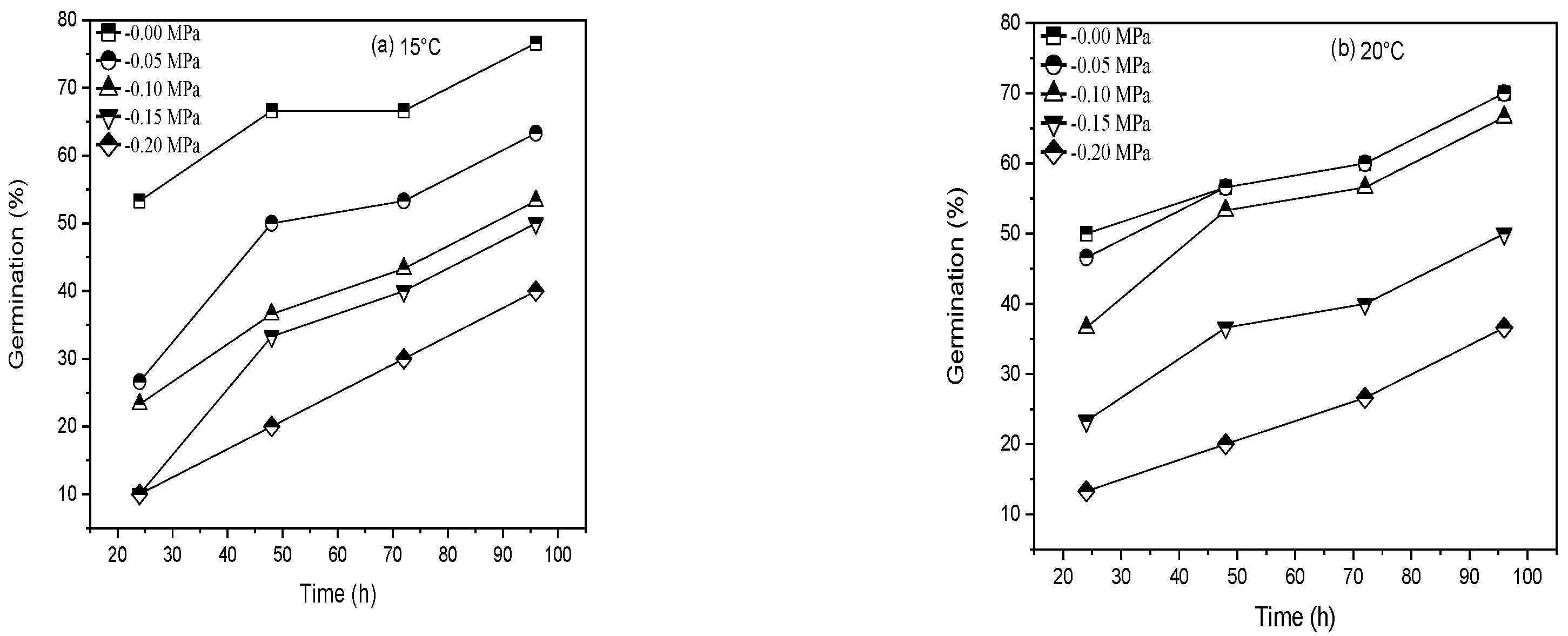
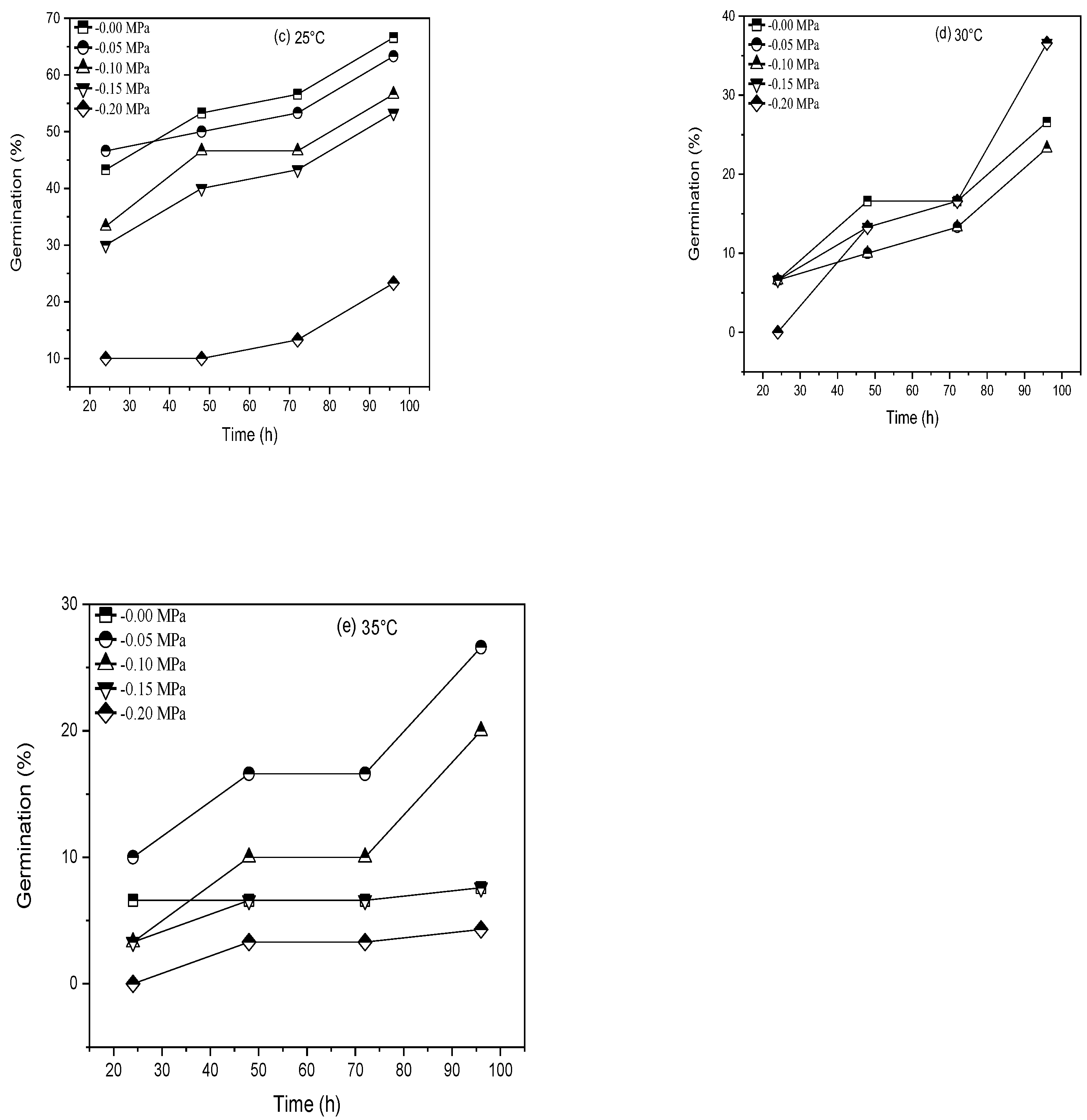
| Cultivar | T (°C) | θH (MPah−1) | Ψb(50) (MPa) | σψb (MPa) | R2 | SE |
|---|---|---|---|---|---|---|
| Pirsabak 15 | 15 °C | 6.24 | −0.11 | 0.161 | 0.997 | 0.0089 |
| 20 °C | 8.61 | −0.12 | 0.148 | 0.856 | 0.0600 | |
| 25 °C | 6.72 | −0.04 | 0.077 | 0.862 | 0.0588 | |
| 30 °C | 2.4 | −0.015 | 0.031 | 0.510 | 0.1167 | |
| 35 °C | 2.4 | −0.03 | 0.063 | 0.775 | 0.0750 | |
| Shahkar 13 | 15 °C | 7.68 | −0.10 | 0.154 | 0.853 | 0.0606 |
| 20 °C | 6.72 | −0.12 | 0.148 | 0.921 | 0.0444 | |
| 25 °C | 8.16 | −0.06 | 0.1 | 0.542 | 0.1070 | |
| 30 °C | 0.96 | −0.02 | 0.063 | 0.800 | 0.0707 | |
| 35 °C | 0.96 | −0.03 | 0.044 | 0.133 | 0.1040 |
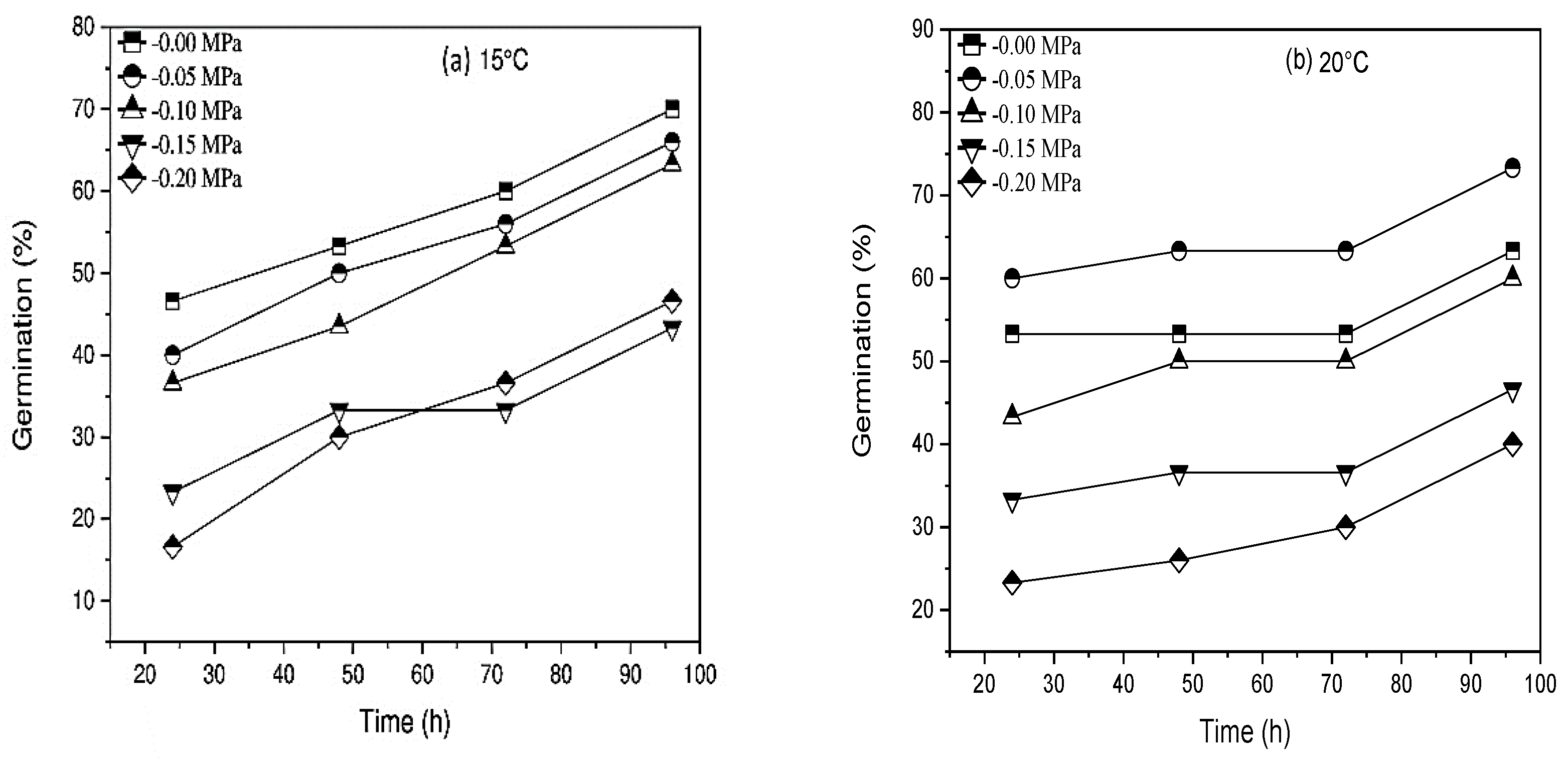
| Cultivar | Pirsabak 15 | Shahkar 13 | Averaged Values |
|---|---|---|---|
| HTT parameter | |||
| Ψb(50) (MPa) | −0.120 | −0.120 | −0.120 |
| σψb (MPa) | 0.148 | 0.148 | 0.148 |
| θHTT (MPa °Ch−1) | 43.20 | 38.40 | 40.80 |
| kT (MPa °Ch−1) | 0.104 | 0.1041 | 0.104 |
| Cardinal Temperature | |||
| Tb (°C) | 15 | 15 | 15 |
| To (°C) | 20 | 20 | 20 |
| Tc (°C) | 35 | 35 | 35 |
| R2 | 0.86 | 0.97 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, S.; Ullah, A.; Ullah, S.; Noor, J.; Ali, B.; Khan, M.N.; Hashem, M.; Mostafa, Y.S.; Alamri, S. Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model. Life 2022, 12, 983. https://doi.org/10.3390/life12070983
Saeed S, Ullah A, Ullah S, Noor J, Ali B, Khan MN, Hashem M, Mostafa YS, Alamri S. Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model. Life. 2022; 12(7):983. https://doi.org/10.3390/life12070983
Chicago/Turabian StyleSaeed, Saleha, Abd Ullah, Sami Ullah, Javaria Noor, Baber Ali, Muhammad Nauman Khan, Mohamed Hashem, Yasser S. Mostafa, and Saad Alamri. 2022. "Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model" Life 12, no. 7: 983. https://doi.org/10.3390/life12070983
APA StyleSaeed, S., Ullah, A., Ullah, S., Noor, J., Ali, B., Khan, M. N., Hashem, M., Mostafa, Y. S., & Alamri, S. (2022). Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model. Life, 12(7), 983. https://doi.org/10.3390/life12070983