
Effects of Different Growth Regulators on the Rooting of Catalpa bignonioides Softwood Cuttings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials, Cutting Treatments, and Sample Collection
2.2. Cuttings and Post-Management Processes
2.3. Determination of Rooting Indicators
2.4. Determination of Physiological and Biochemical Indicators
2.5. Data Processing
3. Results and Analysis
3.1. Rooting of C. bignonioides Softwood Cuttings
3.2. Comparison of Rooting Rate and Callus Production Rate in Response to Different Growth Regulators
3.2.1. Influence of Growth Regulator on the Rooting Rate and Callus Production Rate of C. bignonioides Softwood Cuttings
3.2.2. Effects of Growth Regulator Concentration on the Rooting Rate and Callus Production Rate of C. bignonioides Softwood Cuttings
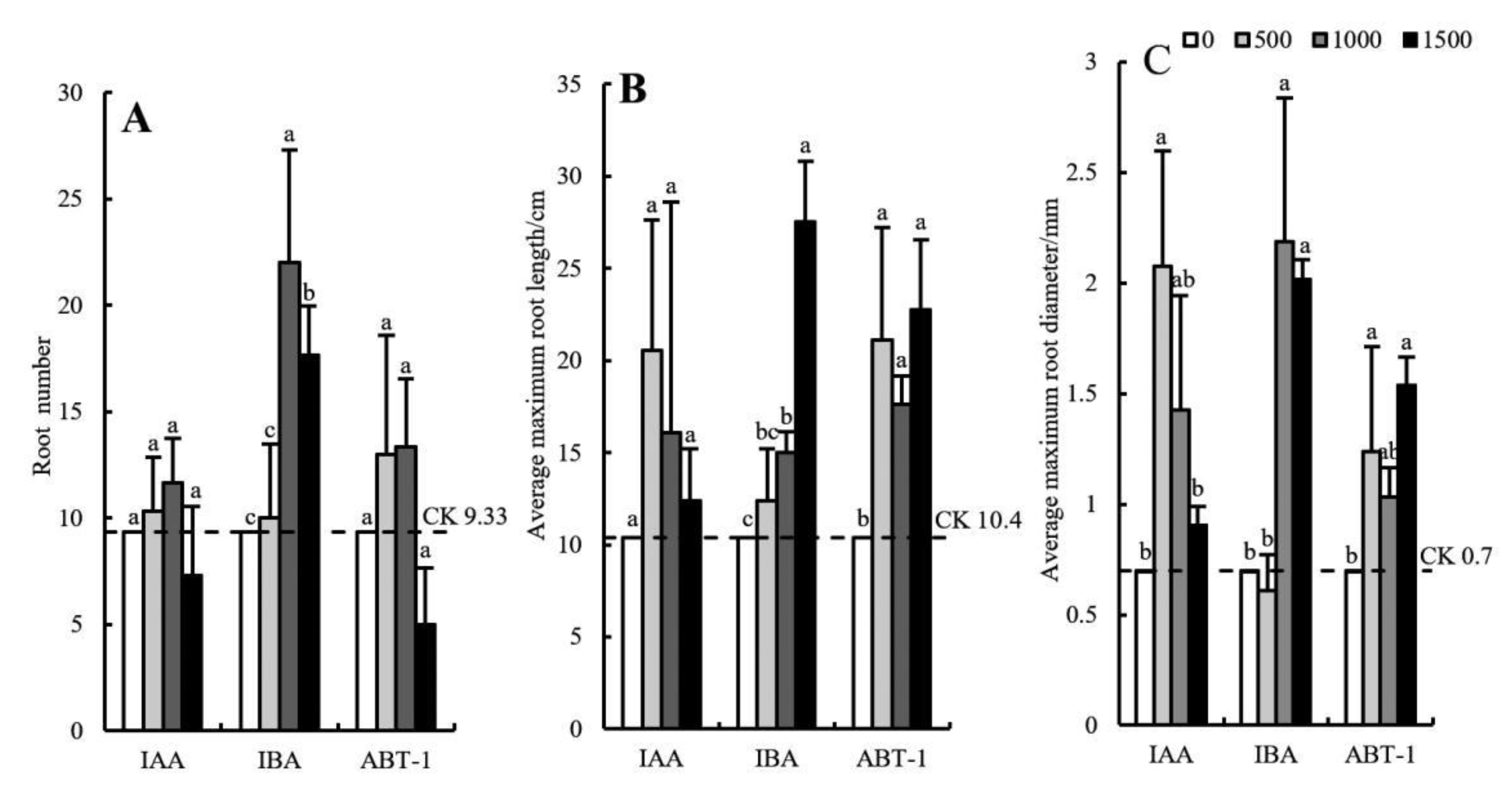
3.3. Comparison of Root Morphology in Response to Different Growth Regulator Treatments
3.3.1. Effects of Growth Regulators on Root Morphology of C. bignonioides Softwood Cuttings
3.3.2. Effects of Growth Regulator Concentration on the Root Morphology of C. bignonioides Softwood Cuttings
3.4. Changes in the Contents of Hormones in the Phloem of C. bignonioides Cuttings under Different Treatments
3.4.1. Changes in the Content of IAA in the Phloem of C. bignonioides Cuttings
3.4.2. Changes in GA3 Content in the Phloem of C. bignonioides Cuttings
3.4.3. Changes in Content of ABA in the Phloem of C. bignonioides Cuttings
3.4.4. Changes in the ZR Content of the Phloem of Cuttings of C. bignonioides
3.4.5. Changes in the IAA/ABA Value of the Phloem of C. bignonioides Cuttings
3.4.6. Changes in the IAA/ZR Value of the Phloem of C. bignonioides Cuttings
4. Discussion
4.1. Effects of Growth Regulators on the Rooting Indexes of C. bignonioides Cuttings
4.2. Changes in the Contents of Endogenous Hormones Associated with Rooting
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, W.C. The Editorial board of “Flora of China” of the Chinese Academy of Sciences was established. Chin. Sci. Bull. 1959, 22, 772. [Google Scholar]
- Zhu, G.F.; Jiang, W.B.; Wen, M.L.; Han, J. Cultural and Historical Connotations of Catalpa ovata and Its Ways of Application in Landscape Greening. Chin. Agric. Sci. Bull. 2010, 26, 301–305. [Google Scholar]
- Wei, Z.P. Evaluation, Screening and Propagation Characteristics of Garden Plants in Hohhot; Inner Mongolia Agricultural University: Hohhot, China, 2008. [Google Scholar]
- Bi, H.T.; Wang, Y.G.; Shen, J.M.; Quan, J.E. Effects of IBA treatment on rooting of cutting and physiological and biochemical twig of Catalpa bignonioides. J. Henan Agric. Univ. 2020, 54, 44–52. [Google Scholar]
- Chen, S.C.; Wang, X.J.; Xiao, Z.D.; Sun, Q.; Xiao, D.J. Primary Report on the Propagation Experiment of Catalpa bungei by Twig Cutting. J. Anhui Agric. Sci. 2008, 36, 7635-7636, 7901. [Google Scholar]
- Liang, Y.W.; Peng, F.R.; Wang, S.C. Preliminary report on cutting experiment of Catalpa bungeana twig. J. For. Eng. J. 2006.
- Zhao, X.Y.; Wang, J.H.; Zhang, J.F.; Zhang, S.F.; Zhang, J.G.; Ma, J.W. Study on phenotypic traits and germination characters of four taxons of catalpa genus seed. J. Northwest AF Univ. (Nat. Sci. Ed.) 2008, 12, 31. [Google Scholar]
- Peng, C.; Li, Z.F.; Xu, X.H.; Huang, G.W.; Chen, H.L. Effect of Different Phytohormone and Their Concentraions and Presoaking Time on Seeds Germination of Catalpa ovata Seed. Seed 2016, 35. [Google Scholar]
- Cui, L.J.; Chen, S.L.; Liu, Y.X.; Lin, J.; Shi, K.M. Germination response of catalpa bungei seed to exogenous hormones treatment under different temperatures. J. Northeast For. Univ. 2020, 2, 12–16. [Google Scholar]
- Hu, T.; Cao, Y.; Zhang, G.X. Softwood Cutting of Chionanthus virginicus and Changes of Endogenous Hormone Content during Rooting Period. J. Northwest For. Univ. 2019, 34, 115–121. [Google Scholar]
- Ren, X.L. Research on cutting techniques and cuttings rooting type in American Nandina domestica‘Alba’. Non-Wood For. Res. 2019, 37, 188–192. [Google Scholar]
- Zhu, Y.C.; Li, B.; Liao, W.B. Effects of three auxins treatments on cutting root ting of Lonicera korolkowi ‘Zabclii’. Pratacult. Sci. 2016, 33, 61–66. [Google Scholar]
- Tong, Z.Q.; Wei, J. Cuttage Propagation Technique and Rooting Me chanism of Quercus mongolica with Softwood Cuttinng. J. Northwest For. Univ. 2017, 32, 116–121. [Google Scholar]
- Bai, L.; Li, R.S.; Yi, G.T.; Yang, J.; Zou, W. Rooting factors and optimization for propagation of Mytilaria laosensis cuttings. J. Zhejiang AF Univ. 2016, 33, 543–550. [Google Scholar]
- Quan, J.E.; Meng, S.; Guo, E.H.; Zhang, S.; Zhao, Z.; Yang, X. De novo sequencing and comparative transcriptome analysis of adventitious root development induced by exogenous indole-3-butyric acid in cuttings of tetraploid black locust. BMC Genom. 2017, 18, 179. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Zhao, Z.; Quan, J.E.; Zhang, X.; Zhang, B. Rooting and Correlative Enzyme Activities of Hardwood Cuttings of Tetraploid Robinia pseudoacacia. Acta Bot. Boreali-Occident. Sin. 2011, 31, 116–122. [Google Scholar]
- Wang, Q.; Zhang, J.; Zhong, C.L.; Zhang, Y.; Wei, Y.C.; Meng, J.X. Variation of endogenesis hormone and nutritive matter concentration in Chukrasia tabularis cuttings during rooting. J. Cent. South Univ. For. Technol. 2020, 40, 111–119. [Google Scholar]
- Zhai, Y.F.; Liu, X.D.; Lv, D.; Zhao, M.; Zhao, H.; Zhao, X.P.; Wang, Y.L.; Zhao, Y.H. Effects of plant growth regulators on the rooting and enzyme activities changes of Lonicera tatarica cuttings. J. Cent. South Univ. For. Technol. 2021, 41, 52–61. [Google Scholar]
- Wang, X.L. Physiological and Biochemical Basis of Adventitious Roots in Tetraploid Locust Cutting; Northwest A&F. University: Xi’an, China, 2011. [Google Scholar]
- Zhang, E.L.; Ma, L.L.; Yang, R.T.; Li, L.; Wang, Q.; Li, Y.; Wang, P. Transcriptome Profiling of IBA-Induced Adventitious Root Formation in Softwood Cuttings of Catalpa bungei‘Yu-1’4. Sci. Silvae Sin. 2018, 54, 48–61. [Google Scholar]
- Ma, L.L.; Wang, P.; Zhang, Z.Y.; Li, L.F.; Yang, R.T.; Li, Y. Evaluation on Rooting Ability of Softwood Cuttings on Catalpa. North. Hortic. 2014, 15, 72–77. [Google Scholar]
- Wang, G.P.; Wang, L.G.; Wang, X.C.; Zhang, C.; Zhang, L.; Liu, B. Dynamic characteristics of cutting rooting of Catalpa bungei with tender branches. J. Nanjing For. Univ. 2020, 44, 94–102. [Google Scholar]
- Zhen, J.H.; Dong, Q.; Duan, H.C. Quality evaluation on Crateva unilocularis cuttings treated with three hormones. Non-Wood For. Res. 2020, 38, 230–237. [Google Scholar]
- Wang, X.S.; Bao, Z.L.; Chen, L.; Cui, L.; Li, F.; Zhang, Y.B. Green Cutting Propagation of Cold Resistant Prunus salicina Linn. J. Northeast For. Univ. 2021, 49, 13–16. [Google Scholar]
- Liang, X.C.; Huang, S.D.; Jiang, Y.M.; Zhao, C.M. Effects of Exogenous Hormones and Substrate on Cutting Rooting of Cryptocarya concinna. Chin. Agric. Sci. Bull. 2021, 37, 24–29. [Google Scholar]
- Yan, Z.J.; Li, H.M.; Yang, P.; Gao, W.S.; Jian, H.Y. Effects of Plant Growth Regulators on Cutting Propagation of Rosa odorata var. Odorata. Acta Agric. Jiangxi 2020, 32, 41–44. [Google Scholar]
- Wu, K.Y.; Gong, B.C.; Xu, Y.; Zhang, P. Effects of different plant growth regulators on hardwood cuttage of persimmon rootstock Yalin 6. Acta Agric. Zhejiangensis 2021, 33, 1256–1263. [Google Scholar]
- Liu, M.G.; Wang, L.; Dong, S.J.; Cai, X. Endogenous hormone variation in cuttings of Thuja occidentalis L. in the period of adventitious root formation. J. Shenyang Agric. Univ. 2010, 41, 555–559. [Google Scholar]
- He, C.D.; Yao, R.; Huang, R.C.; Mi, X.Q.; Zhang, C.; Tan, Y.M.; Wang, Y.Q. Effects of K-IBA treatments on adventitious rooting and endogenous hormones contents of shoot cuttings of Thuja occidentals L. J. Cent. South Univ. For. Technol. 2017, 37, 7–12. [Google Scholar]
- Zhang, X.P.; Fang, Y.M.; Huang, S.H. The endogenous hormone’s variation during theadventitious roots formation of hybrid Tuliptrees by cutting. J. Nanjing For. Univ. 2004, 28, 79–82. [Google Scholar]
- Li, C.C.; Zhao, Y.L.; Zhang, D.L.; Wu, H.; Chen, X. Changes of endogenous hormones and anatomical structure of Rhododendron stamineum during cutting propagation. For. Res. 2012, 25, 360–365. [Google Scholar]
- Zhao, Y.L.; Chen, X.; Li, C.C. Dynamic of physiology and biochemistry duringwild Rhododendron scabrifolium cutting propagation. Sci. Silvae Sin. 2013, 49, 45–51. [Google Scholar]
- Li, Y.X.; Zeng, H.J.; Wang, X.M.; Cai, N. Changes of endogenous hormones during Swida wilsoniana wanger cutting. Chin. Agric. Sci. Bull. 2010, 26, 247–251. [Google Scholar]
- Dong, S.J.; Liu, M.G.; Dai, F.; Wu, Y.L.; Shan, S.T.; Ding, R.J. Variation of endogenous hormone contents in softwood cuttings of Armeniaca sibirica during adventitious root formation. Non-Wood For. Res. 2013, 31, 108–114. [Google Scholar]
- Shi, F.H.; Zhao, R.; Luo, S.; Zhen, C.; Shen, Y.B. Study on plant hormone changes in soft cuttings of Tiliamiqueliana during rooting. J. Cent. South Univ. For. Technol. 2019, 39, 21–26. [Google Scholar]
- Sun, A.; Li, L.F.; Guo, L.; Su, N. Reponse of Rooting Traits of Machilus longipedicellata Rooted Cutting to IBA, NAA and IAA. North. Hortic. 2017, 16, 102–106. [Google Scholar]
- Wiesman, Z.; Lavee, S. Enhancement of IBA stimulatory effect on rooting of olive cultivar stem cuttings. Sci. Hortic. 1995, 62, 189–198. [Google Scholar] [CrossRef]
- Copes, D.L.; Mandel, N.L. Effects of IBA and NAA treatments on rooting Douglas-fir stemcuttigs. New For. 2000, 20, 249–257. [Google Scholar] [CrossRef]
- Wang, X.M.; Peng, D.Q.; Wu, W.L.; Zhu, H.; Huang, T.; Lv, L.F.; Li, W.L. Effects of culture Medium and plant growth regulators on rooting of softwood cuttings of beach plum. J. Northwest For. Univ. 2014, 29, 114–118. [Google Scholar]
- Xu, S.S.; Liu, X.J.; Xu, D.P.; Hong, Z.; Guo, J.-Y.; Yang, Z.-J. Influence of IAA and NAA on cutting propagation of dalbergia odorifera. For. Res. 2021, 34, 168–176. [Google Scholar]
- Zheng, J.H.; Dong, Q.; Duan, H.C.; Cha, X.F.; Zhou, C.; Zhou, Z.C. Effect of Three Growth Regulators on Rooting of Cuting of Crateva unilocularis. Bull. Bot Res. 2020, 40, 202–208. [Google Scholar]
- Garrido, G.; Guerrero, J.R.; Cano, E.M. Origin and basipetal transport of the IAA responsible for carnation cuttings. Physiol. Plant. 2002, 31, 302–312. [Google Scholar] [CrossRef]
- Haussig, B.E. Influence of indole-3-acetic acid on adventitious root primordia of brittle willow. Planta 1970, 95, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Trefois, R.; Brunner, T. Influence of the endogenous auxinic contene on the response to propagation by cuttings and on the responce to propagation by cuttings and on the dwarfing effect of some types of Prunus. Bot.-Bot. Publ. 1982, 69, 197–204. [Google Scholar]
- Xu, J.Z.; Chen, S.W. The effect of the endogenous hormone’s contents (ABA and IAA) in hardwood cuttings of peach to rooting. Acta Hortic. Sin. 1989, 4, 275–278. [Google Scholar]
- Lv, M.; Fang, Y.M.; Jia, H.; Wang, A.X. Dynamics of endogenous hormones during the process of adventitious root formation in Red Alder. Dev. For. Sci. Technol. 2009, 23, 16–19. [Google Scholar]
- Taramino, G.; Sauer, M.; Stauffer, J.L., Jr.; Multani, D.; Niu, X.; Sakai, H.; Hochholdinger, F. The maize (Zea mays L.) RTCS gene encodes a LOB domain protein that is a key regulator of embryonic seminal and post-embryonic shoot-borne root initiation. Plant J. 2007, 50, 649–659. [Google Scholar] [CrossRef]
- Wang, Q. Propagation Techniques and Rooting Mechanism of Chukrasia Tabularis Softwood Cuttings; Northeast Forestry University: Harbin, China, 2019. [Google Scholar]
- Zhao, X.M.; Huo, C.F.; Shen, H.L. Studied on changes hormone in the periods of making roots in the softwood of Robinia pseudoacacia. J. Northwest For. Univ. 2013, 28, 18–23. [Google Scholar]
- Han, H.; Zhang, S.G.; Sun, X.M. A review on the molecular mechanism of plants rooting modulated by auxin. Afr. J. Biotechnol. 2009, 8, 348–353. [Google Scholar]
- Nanda, K.K.; Anand, V.K.; Chibbar, R.N. The promotive Effect of gibberellic on the production of adventitious roots on stem cuttings of Ipomoea fistulosa. Planta 1972, 105, 360. [Google Scholar] [CrossRef]
- Sun, X.D.; Zhang, P.; Shen, S.L. Effects of Rooting Agents and Cutting Treatment on Rooting of Shoot Cuttings of Acer mono. J. Northest. For. Univ. 2018, 46, 5–8. [Google Scholar]
- Chi, Y.; Hou, Y.L.; Yu, Y.J.; Yu, D.Y.; Feng, B.M.; Yao, Z.A.; Jiang, G.B. Dynamic Changes of Endogenous Hormones Contents in Mutant of Populus tomentosa and its Relative Species in the Cutting and Rooting Procedure. J. Anhui Agric. Sci. 2007, 35, 8438–8439. [Google Scholar]
- Lin, S.J.; Jiang, J.; Feng, X.; Li, T.H.; Chang, W.X. The Study of Adventitious Root Generation in Leaves of Tissue Culture of (P.simonii × P.nigra) × P.15 ACL in the Relation to Exogenous Hormones and Endogenous Hormones. For. Sci. Technol. 2016, 21, 68–79. [Google Scholar]
- Huang, Z.L.; He, H.K.; Cao, Y.Y.; Peng, Y.H.; Wang, X.J.; Liu, X. Changes of endogenous hormones in Castanopsis hystrix cuttings. J. Cent. South Univ. For. Technol. 2015, 2, 22–25. [Google Scholar]
- Ao, H.; Wang, K.; Feng, Y.L. Endogenous hormones levels in cuttings of Larix olgensis and the irrelations to rooting. Bull. Bot. Res. 2002, 22, 190–195. [Google Scholar]
- Ao, H.; Wang, K.; Feng, Y.L. Qualitative Analysis of Endogenesis Hormone and Polyphenol During Rooting of Cuttings in Norway Spruce (Picea abies). For. Sci. 2015, 51, 155–162. [Google Scholar]
- Han, J.H.; Shao, W.H.; Liu, J.F.; Diao, S.F. Content changes of endogenous hormone and polyphenols during hardwood cuttage progress in Sapindus ukorossi. Non-Wood For. Res. 2019, 37, 37–43+51. [Google Scholar]
- Qiao, Z.Q.; Wang, X.M.; Zeng, H.J.; Li, Y.X.; Cai, N.; Wang, X.Y. Changes of endogenous hormones during Lagerstroemia indica ‘Xiang Yun’cutting. Hunan For. Sci Technol. 2015, 42, 49–53. [Google Scholar]
- Zhou, Z.Z.; Liu, S.C.; Zhang, J.H.; Liang, K.N.; Ma, H.M.; Huang, G.H. Effects of IBA treatments on adventitious rooting and endogenous hormones contents of shoot cuttings of Callicarpa nudiflora. Chin. J. Trop. Crops 2016, 37, 1075–1080. [Google Scholar]
- Guo, S.J.; Lin, H.Q.; Li, F.L. Progress of Study on Rooting Anatomy and Physiology of Forest Tree Cuttings. J. Beijing For. Univ. 1997, 4, 64–69. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Number | A Growth Regulators | B Concentration (mg·L−1,) | C Treatment Time (h) |
|---|---|---|---|
| T1 | IAA | 500 | 1 |
| T2 | IAA | 1000 | 1 |
| T3 | IAA | 1500 | 1 |
| T4 | IBA | 500 | 1 |
| T5 | IBA | 1000 | 1 |
| T6 | IBA | 1500 | 1 |
| T7 | ABT-1 | 500 | 1 |
| T8 | ABT-1 | 1000 | 1 |
| T9 | ABT-1 | 1500 | 1 |
| CK | Water |
| Growth Regulator | Rooting Rate/% | Callus Production Rate/% |
|---|---|---|
| IAA | 32.89 ± 1.68 cC | 8.22 ± 3.15 bAB |
| IBA | 62.44 ± 2.14 aA | 18.44 ± 3.67 aA |
| ABT-1 | 49.33 ± 1.76 bB | 17.56 ± 5.35 aA |
| CK | 8.00 ± 2.00 dD | 2.00 ± 2.00 bB |
| Growth Regulator | Number of Roots/n | Average Maximum Root Length/cm | Average Maximum Root Diameter/mm |
|---|---|---|---|
| IAA | 8.33 ± 2.91 bA | 16.34 ± 7.38 abA | 1.47 ± 0.28 aA |
| IBA | 16.56 ± 3.42 aA | 18.31 ± 0.88 aA | 1.61 ± 0.30 aA |
| ABT-1 | 10.11 ± 3.79 bA | 20.51 ± 0.62 aA | 1.27 ± 0.19 aAB |
| CK | 7.00 ± 3.00 bA | 10.40 ± 1.13 bA | 0.70 ± 0.10 bB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, J.; Ni, R.; Wang, Y.; Sun, J.; Ma, M.; Bi, H. Effects of Different Growth Regulators on the Rooting of Catalpa bignonioides Softwood Cuttings. Life 2022, 12, 1231. https://doi.org/10.3390/life12081231
Quan J, Ni R, Wang Y, Sun J, Ma M, Bi H. Effects of Different Growth Regulators on the Rooting of Catalpa bignonioides Softwood Cuttings. Life. 2022; 12(8):1231. https://doi.org/10.3390/life12081231
Chicago/Turabian StyleQuan, Jin’e, Ruoyi Ni, Yange Wang, Jiajia Sun, Mingyue Ma, and Huitao Bi. 2022. "Effects of Different Growth Regulators on the Rooting of Catalpa bignonioides Softwood Cuttings" Life 12, no. 8: 1231. https://doi.org/10.3390/life12081231
APA StyleQuan, J., Ni, R., Wang, Y., Sun, J., Ma, M., & Bi, H. (2022). Effects of Different Growth Regulators on the Rooting of Catalpa bignonioides Softwood Cuttings. Life, 12(8), 1231. https://doi.org/10.3390/life12081231