
Analysis of Floral Organ Development and Sex Determination in Schisandra chinensis by Scanning Electron Microscopy and RNA-Sequencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Scanning Electron Microscopy Sample Processing
2.3. RNA Extraction and Next Generation Sequencing
2.4. Gene Function Annotation and Differential Gene Expression Analysis
2.5. Validation of RNA-seq by RT-qPCR
3. Results
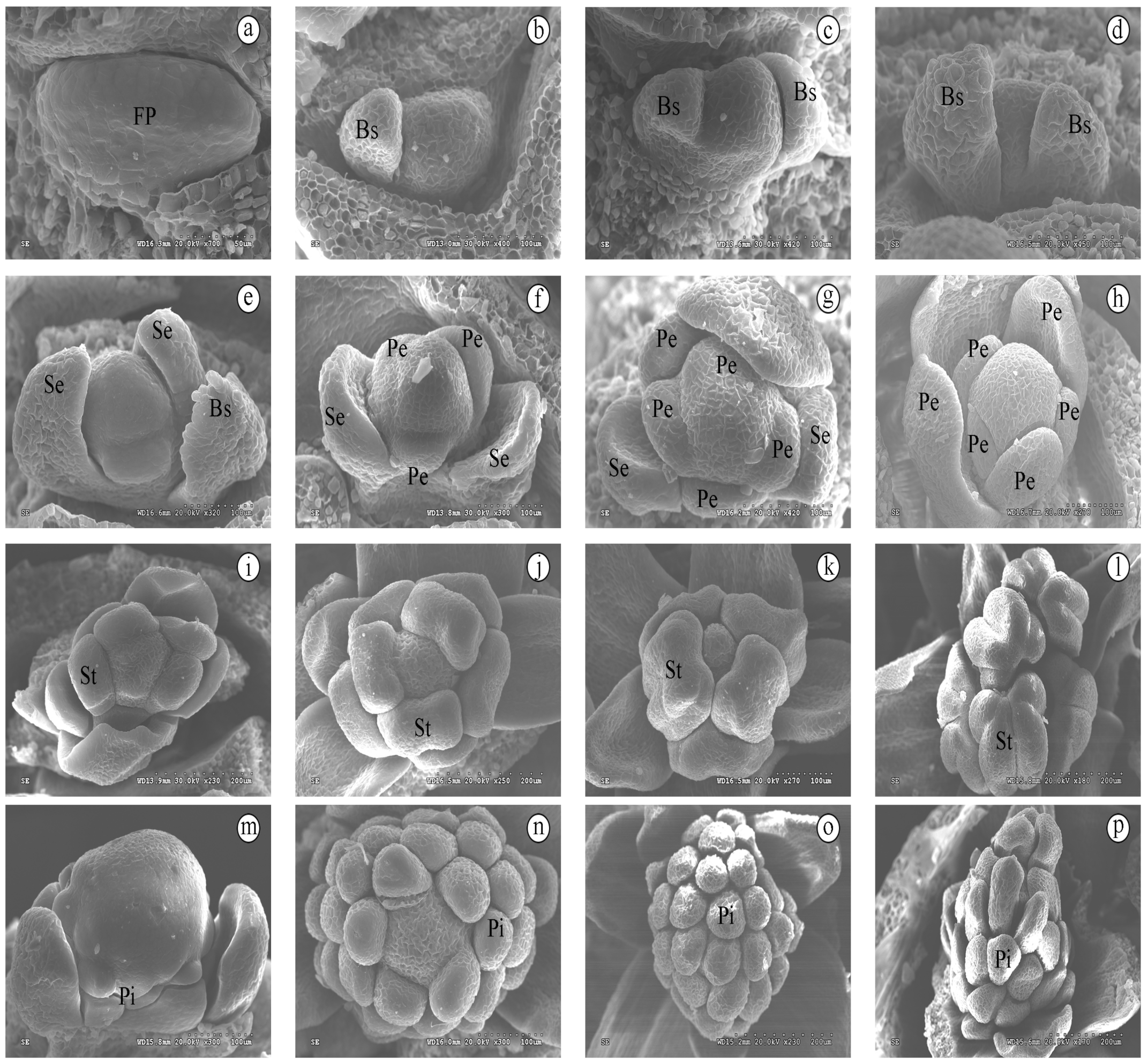
3.1. Development of Floral Organ
3.2. Continuing Development of Floral Organ
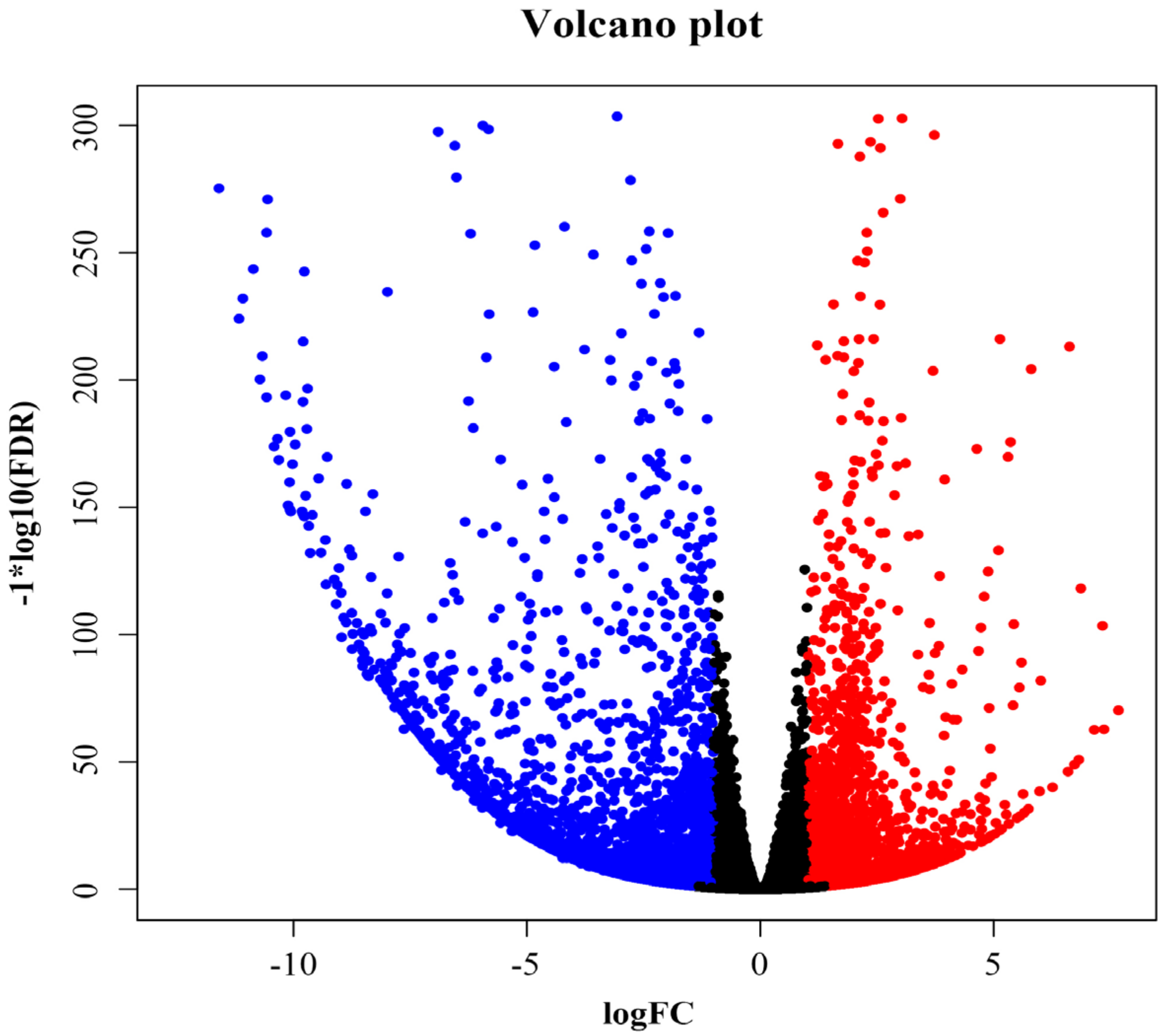
3.3. Overview of RNA-Seq Sequencing
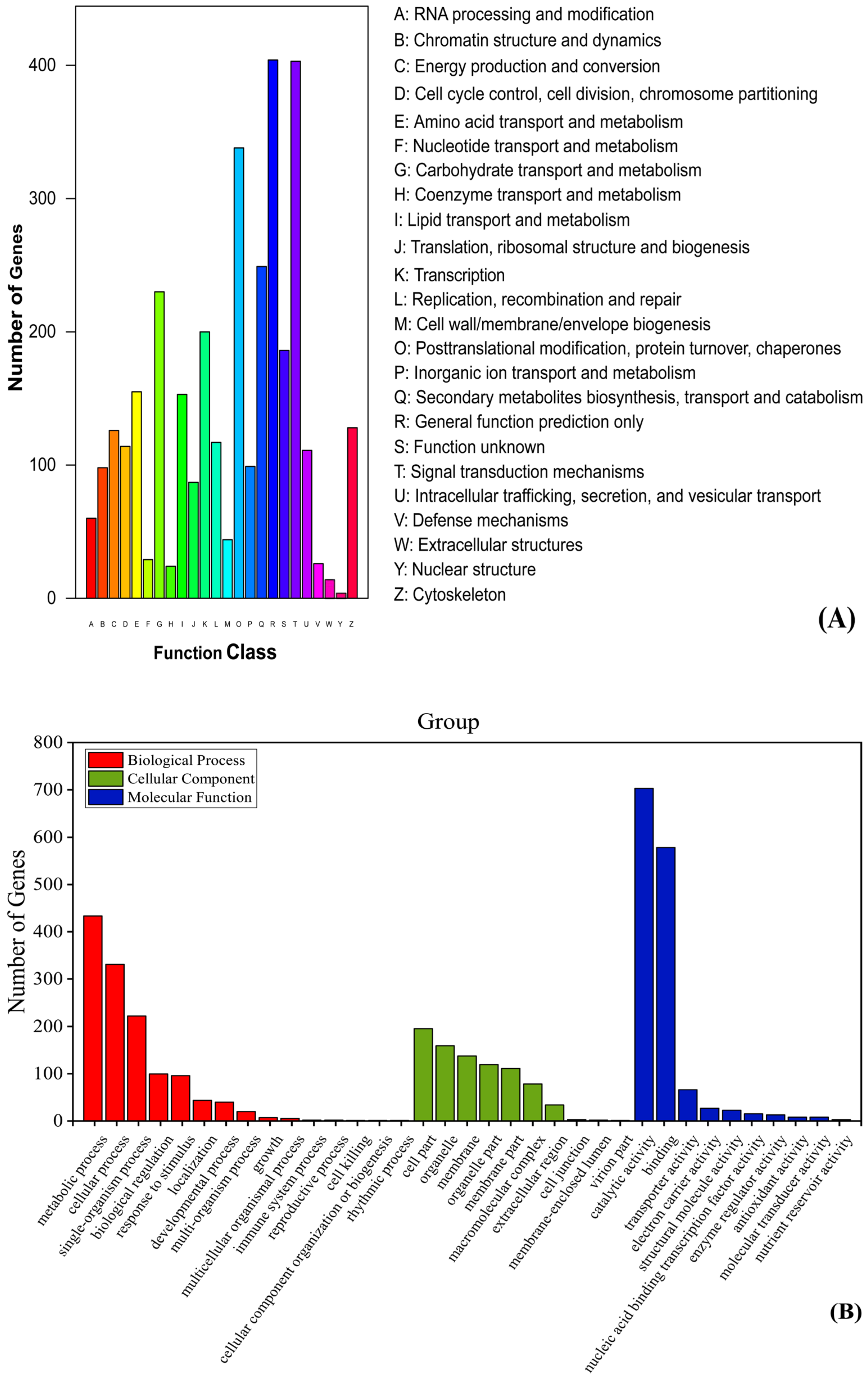
3.4. Differential Genes Function Analysis
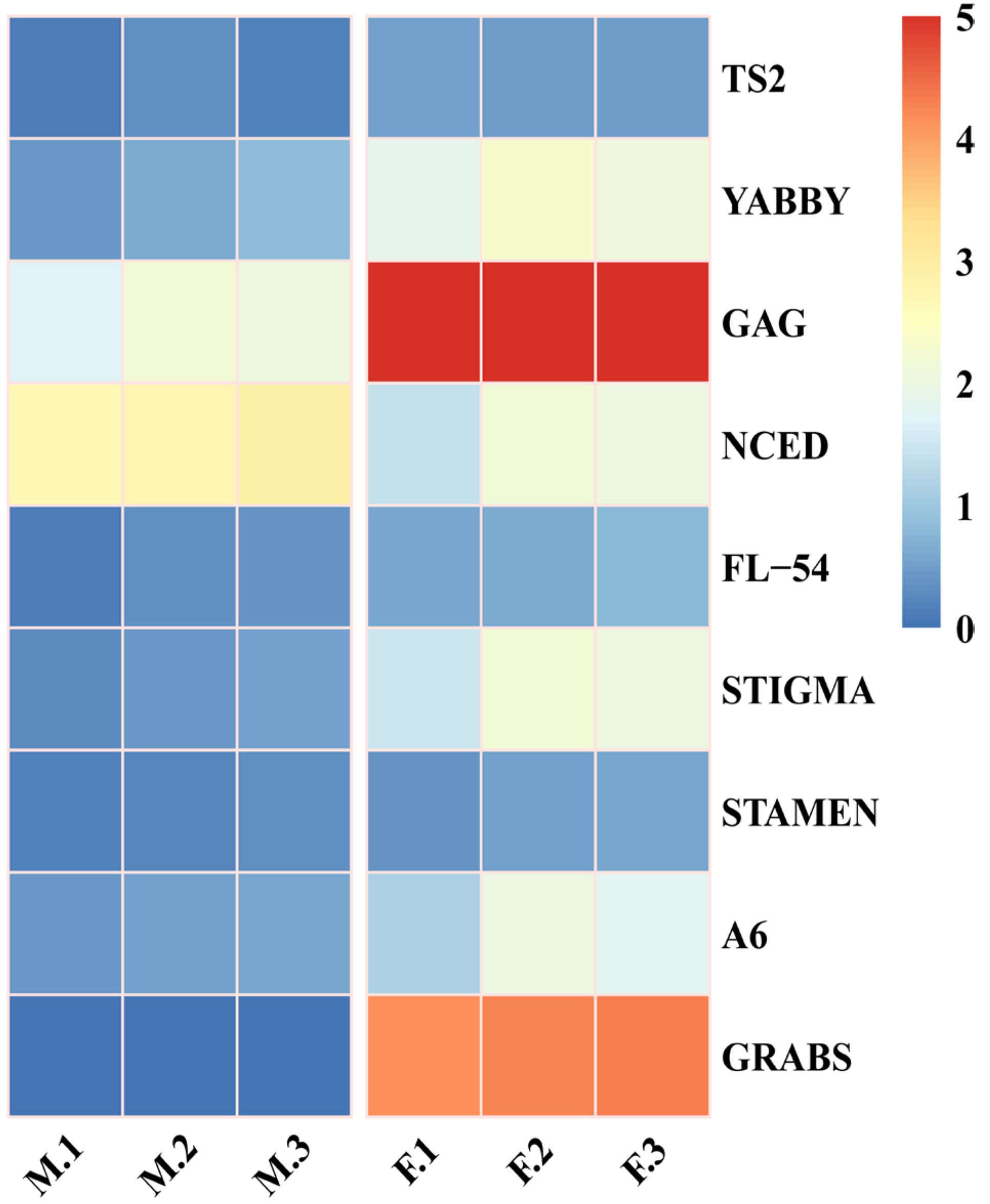
3.5. Identification of Genes Associated with Flower Development from DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hancke, J.L.; Burgos, R.A.; Ahumada, F. S. chinensis (Turcz.) Baill. Fitoterapia 1999, 70, 451–471. [Google Scholar] [CrossRef]
- Yang, S.Y.; Yuan, C.H. S. chinensis: A comprehensive review on its phytochemicals and biological activities. Arab. J. Chem. 2021, 14, 103310. [Google Scholar] [CrossRef]
- Mayer, S.S.; Charlesworth, D. Cryptic dioecy in flowering plants. Trends Ecol. Evol. 1991, 6, 320–325. [Google Scholar] [CrossRef]
- Akagi, T.; Kajita, K.; Kibe, T.; Morimura, H.; Tsujimoto, T.; Nishiyama, S.; Kawai, T.; Yamane, H.; Tao, R. Development of molecular markers associated with sexuality in Diospyros lotus L. and their application in D. kaki Thunb. J. Jpn. Soc. Hortic. Sci. 2013, 83, 214–221. [Google Scholar] [CrossRef]
- Mitchell, C.H.; Diggle, P.K. The evolution of unisexual flowers: Morphological and functional convergence results from diverse developmental transitions. Am. J. Bot. 2005, 92, 1068–1076. [Google Scholar] [CrossRef]
- Sanderson, B.J.; Feng, G.Q.; Hu, N.; Carlson, C.H.; Smart, L.B.; Keefover-Ring, K.; Yin, T.M.; Ma, T.; Liu, J.Q.; DiFazio, S.P.; et al. Sex determination through X-Y heterogamety in Salix nigra. bioRxiv 2020, 3, 000919. [Google Scholar] [CrossRef]
- Anupharb, S.; Lucsame, G.; Tittinat, P.; Prapassorn, D.E.; Jantrararuk, T.; Siam, P. Transcriptome analysis of Jatropha curcas L. flower buds responded to the paclobutrazol treatment. Plant Physiol. Biochem. 2018, 127, 276–286. [Google Scholar]
- Liu, X.L.; Li, J.H.; Zhu, J.Y.; Yang, Y.F. Floral differentiation and growth rhythm of rhizome buds of the spring ephemeroid plant Adonis amurensis Regel et Radde. Phyton-Int. J. Exp. Bot. 2016, 85, 297–304. [Google Scholar]
- Zhang, L.J.; Guo, C.; Lu, X.J.; Sun, X.M.; Liu, C.P.; Zhou, Q.; Deng, J.F. Flower Development of Heterodichogamous Juglans mandshurica (Juglandaceae). Front. Plant Sci. 2021, 12, 541163. [Google Scholar] [CrossRef]
- Di, S.V.; Kramer, E.M.; Baum, D.A. Floral MADS box genes and homeotic gender dimorphism in Thalictrum dioicum (Ranunculaceae)—A new model for the study of dioecy. Plant J. 2005, 41, 755–766. [Google Scholar]
- Maja, J.; Noah, S.D.; Golenberg, E.M. Functional analysis of B and C class floral organ genes in spinach demonstrates their role in sexual dimorphism. BMC Plant Biol. 2010, 10, 46. [Google Scholar]
- Hu, X.Q.; Liao, Z.Y.; Zhang, B.; Yue, J.J.; Wang, Z.; Jie, X.; Liu, J. Transcriptome sequencing and screening of genes related to sex determination of Trichosanthes kirilowii Maxim. PLoS ONE 2020, 15, e0239230. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Akagi, T.; Kawakatsu, T.; Tao, R. Gene networks orchestrated by MeGI: A single-factor mechanism underlying sex determination in persimmon. Plant J. Cell Mol. Biol. 2019, 98, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Zhang, G.J.; Zhang, X.J.; Yuan, J.H.; Deng, C.L.; Gao, W.J. Comparative transcriptome analysis reveals differentially expressed genes associated with sex expression in garden asparagus (Asparagus officinalis). BMC Plant Biol. 2017, 17, 143. [Google Scholar] [CrossRef]
- Mao, Y.J.; Liu, W.B.; Chen, X.; Xu, Y.; Lu, W.L.; Hou, J.Y.; Ni, J.; Wang, Y.T.; Wu, L.F. Flower Development and Sex Determination between Male and Female Flowers in Vernicia fordii. Front. Plant Sci. 2017, 8, 1291. [Google Scholar] [CrossRef]
- Li, N.; Wang, Y.Y.; Wang, J.W.; Zhang, W.Q.; Meng, Z.W.; Wang, Y.S.; Zhang, Y.L.; Li, S.F.; Gao, W.J.; Deng, C.L. Identification of Sex Differentiation-Related microRNAs in Female and Male Flower. Int. J. Mol. Sci. 2022, 23, 4090. [Google Scholar] [CrossRef]
- Murase, K.; Shigenobu, S.; Fujii, S.; Ueda, K.; Murata, T.; Sakamoto, A.; Wada, Y.; Yamaguchi, K.; Osakabe, Y.; Osakabe, K.; et al. MYB transcription factor gene involved in sex determination in Asparagus officinalis. Genes Cells Devoted Mol. Cell. Mech. 2017, 22, 115–123. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Wang, H.Y.; Xu, Z.S.; Zhang, H.L.; Li, G.L.; Wang, X.W.; Qian, W. Transcriptome profiling of differentially expressed genes of male and female inflorescences in spinach (Spinacia oleracea L.). Genome 2021, 64, 777–788. [Google Scholar] [CrossRef]
- Li, X.; Han, R.; Cai, K.W.; Guo, R.X.; Pei, X.N.; Zhao, X.Y. Characterization of Phytohormones and Transcriptomic Profiling of the Female and Male Inflorescence Development in Manchurian Walnut (Juglans mandshurica Maxim.). Int. J. Mol. Sci. 2022, 23, 5433. [Google Scholar] [CrossRef]
- Monika, H.; Kuldeep, S.; Manoj, P.; Veena, A. Review on different mechanisms of sex determination and sex-linked molecular markers in dioecious crops: A current update. Euphytica 2015, 201, 161–194. [Google Scholar]
- Boualem, A.; Troadec, C.; Camps, C.; Lemhemdi, A.; Morin, H.; Sari, M.-A.; Fraenkel-Zagouri, R.; Kovalski, I.; Dogimont, C.; Perl-Treves, R.; et al. A cucurbit androecy gene reveals how unisexual flowers develop and dioecy emerges. Science 2015, 350, 688–691. [Google Scholar]
- Zhang, J.S.; Adnane, B.; Abdelhafid, B.; Ray, M. Genomics of sex determination. Curr. Opin. Plant Biol. 2014, 18, 110–116. [Google Scholar] [CrossRef]
- Luo, Y.; Pan, B.Z.; Li, L.; Yang, C.X.; Xu, Z.F. Developmental basis for flower sex determination and effects of cytokinin on sex determination in Plukenetia volubilis (Euphorbiaceae). Plant Reprod. 2020, 33, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Hui, F.; Yang, Y.C.; Yang, S.H. Deep sequencing and transcriptome analysis to identify genes related to biosynthesis of aristolochic acid in Asarum heterotropoides. Sci. Rep. 2018, 8, 17850. [Google Scholar] [CrossRef] [PubMed]
- Dewey, C.N.; Li, B. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Pfaf, M.W. A new mathematical model for relative quantification real-time PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar]
- Braun, D.M.; Wang, L.; Ruan, Y.L. Understanding and manipulating sucrose phloem loading, unloading, metabolism, and signalling to enhance crop yield and food security. J. Exp. Bot. 2014, 65, 1713–1735. [Google Scholar] [CrossRef]
- Chen, L.Y.; Chu, C.Y.; Huang, M.C. Inflorescence and flower development in Chinese ixora. Am. Soc. Hort. Sci. 2003, 128, 23–28. [Google Scholar] [CrossRef]
- Foster, T.; Johnston, R.; Seleznyova, A. A morphological and quantitative characterization of early floral development in apple (Malus x domestica Borkh). Ann. Bot. 2003, 92, 199–206. [Google Scholar] [CrossRef]
- Gross, T.; Broholm, S.; Becker, A. CRABS CLAW acts as a bifunctional transcription factor in flower development. Front. Plant Sci. 2018, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Smyth, D.R. CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 1999, 126, 2387–2396. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mena, C.; Folter, S.; Rosta, M.M. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development 2004, 132, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Michael, L.; Andrea, B.; Gerd, J.; Thomas, L. Termination of Stem Cell Maintenance in Arabidopsis Floral Meristems by Interactions between WUSCHEL and AGAMOUS. Cell 2001, 105, 805–814. [Google Scholar]
- Guo, L.; Cao, X.W.; Liu, Y.H.; Li, J.; Li, Y.P.; Li, D.M.; Zhang, K.; Gao, C.X.; Dong, A.W.; Liu, X.W. A chromatin loop represses WUSCHEL expression in Arabidopsis. Plant J. 2018, 94, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Sablowski, R. Flowering and determinacy in Arabidopsis. J. Exp. Bot. 2007, 58, 899–907. [Google Scholar] [CrossRef]
- Lohmann, J.U.; Ray, L.H.; Martin, H.; Maximilian, A.B.; François, P.; Rüdiger, S.; Detlef, W. A Molecular Link between Stem Cell Regulation and Floral Patterning in Arabidopsis. Cell 2001, 105, 793–803. [Google Scholar] [CrossRef]
- Cao, X.W.; He, Z.S.; Guo, L.; Liu, X.G. Epigenetic Mechanisms Are Critical for the Regulation of WUSCHEL Expression in Floral Meristems. Plant Physiol. 2015, 168, 1189–1196. [Google Scholar] [CrossRef]
- Cristel, C.C.; Jennifer, C.F. The SAND domain protein ULTRAPETALA1 acts as a trithorax group factor to regulate cell fate in plants. Cold Spring Harb. Lab. Press. 2009, 23, 2723–2728. [Google Scholar]
- Carles, C.C.; Choffnes, I.D.; Reville, K. ULTRAPETALA1 encodes a SAND domain putative transcriptional regulator that controls shoot and floral meristem activity in Arabidopsis. Development 2005, 132, 897–911. [Google Scholar] [CrossRef]
- Engelhorn, J.; Moreau, F.; Fletcher, J.C.; Carles, C.C. ULTRAPETALA1 and LEAFY pathways function independently in specifying identity and determinacy at the Arabidopsis floral meristem. Ann. Bot. 2014, 114, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, R.A.; Bollman, K.; Taylor, R. KANADI regulates organ polarity in Arabidopsis. Nat. Int. Wkly. J. Sci. 2001, 411, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Pires, H.R.; Shemyakina, E.A.; Fletcher, J.C. The ULTRAPETALA1 trxG factor contributes to patterning the Arabidopsis adaxial-abaxial leaf polarity axis. Plant Signal. Behav. 2015, 10, 1559–2324. [Google Scholar] [CrossRef]
- Ito, T.; Ng, K.H.; Lim, T.S.; Yu, H.; Meyerowitz, E.M. The homeotic protein agamous controls late stamen development by regulating a jasmonate biosynthetic gene in Arabidopsis. Plant Cell. 2007, 19, 3516–3529. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; García-Ponce, B.; Fonseca-Salazar, G.; Alvarez-Buylla, E.R.; Yu, H. AGAMOUS-LIKE 17, a novel flowering promoter, acts in a FT-independent photoperiod pathway. Plant J. 2008, 55, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.A.; Yanofsky, M.F. The Arabidopsis AGL9 MADS box gene is expressed in young flower primordia. Sex. Plant Reprod. 1998, 11, 22–28. [Google Scholar] [CrossRef]
- Xu, F.; Dou, J.S.; Wang, L.L.; Yan, J.P.; Fu, M.Y.; Zhang, X. Molecular Cloning and Expression Analysis of a AGAMOUS-like 66 Gene (GbAGL66) in Ginkgo biloba. Biotechnology 2017, 16, 100–107. [Google Scholar]
- Mandaokar, A.; Thines, B.; Shin, B.; Lange, B.M.; Choi, G.; Koo, Y.J.; Yoo, Y.J.; Choi, Y.D.; Choi, G.; Browse, J. Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. 2006, 46, 984–1008. [Google Scholar] [CrossRef]
- Shen, X.P.; Hu, Z.W.; Xiang, X.; Xu, L.A.; Cao, J.S. Overexpression of a stamen-specific R2R3-MYB gene BcMF28 causes aberrant stamen development in transgenic Arabidopsis. Biochem. Biophys. Res. Commun. 2019, 518, 726–731. [Google Scholar] [CrossRef]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA -regulated genes that redundantly facilitate anther development. Plant Cell. 2005, 17, 705–721. [Google Scholar] [CrossRef]
- Phan, H.A.; Iacuone, S.; Li, S.F.; Parish, R.W. The MYB80 transcription factor is required for pollen development and the regulation of tapetal programmed cell death in Arabidopsis thaliana. Plant Cell. 2011, 23, 2209–2224. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Song, J.; Ferguson, A.C.; Klisch, D.; Simpson, K.; Mo, R.; Taylor, B.; Mitsuda, N.; Wilson, Z.A. Transcription factor MYB26 is key to spatial specificity in anther secondary thickening formation. Plant Physiol. 2017, 175, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Ghelli, R.; Brunetti, P.; Napoli, N.; De, P.A.; Cecchetti, V.; Tsuge, T.; Serino, G.; Matsui, M.; Mele, G.; Rinaldi, G.; et al. A newly identified flower-specific splice variant of AUXIN RESPONSE FACTOR8 regulates stamen elongation and endothecium lignification in Arabidopsis. Plant Cell. 2018, 30, 620–637. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Zhu, J.; Gao, J.F.; Wang, C.; Li, H.; Li, H.; Zhang, H.Q.; Zhang, S.; Wang, D.M.; Wang, Q.X.; et al. Transcription factor AtMYB103 is required for another development by regulating tapetum development, callose dissolution and exine formation in Arabidopsis. Plant J. 2007, 52, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Heisler, M.G.; Atkinson, A.; Bylstra, Y.H.; Walsh, R.; Smyth, D.R. SPATULA, a gene that controls development of carpel margin tissues in Arabidopsis, encodes a bHLH protein. Development 2001, 128, 1089–1098. [Google Scholar] [CrossRef]
- Michael, G.; Teodora, P.; David, R.S. Functional domains of SPATU-LA, a bHLH transcription factor involved in carpel and fruit development in Arabidopsis. Plant J. 2008, 55, 40–52. [Google Scholar]
- Gremski, K.; Ditta, G.; Yanofsky, M.F. The HECATE genes regulate female reproductive tract development in Arabidopsis thaliana. Development 2007, 134, 3593–3601. [Google Scholar] [CrossRef]
- Zhu, J.; Lou, Y.; Xu, X.F.; Yang, Z.N. A genetic pathway for tapetum development and function in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 892–900. [Google Scholar] [CrossRef]
- Gu, J.N.; Zhu, J.; Yu, Y.; Teng, X.D.; Lou, Y.; Xu, X.F.; Liu, J.L.; Yang, Z.N. DYT1 directly regulates the expression of TDF1 for tapetum development and pollen wall formation in Arabidopsis. Plant J. 2014, 80, 1005–1013. [Google Scholar] [CrossRef]
- Li, W.; Wang, H.P.; Yu, D.Q. Arabidopsis WRKY transcription factors WRKY12 and WRKY13 oppositely regulate flowering under short-day conditions. Mol. Plant. 2016, 9, 1492–1503. [Google Scholar] [CrossRef]
- Yu, Y.C.; Hu, R.B.; Wang, H.M.; Cao, Y.P.; He, G.; Fu, C.X.; Zhou, G.K. MlWRKY12, a novel Miscanthus transcription factor, participates in pith secondary cell wall formation and promotes flowering. Plant Sci. 2013, 212, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.C.; Liu, Z.H.; Wang, L.; Kim Sang, G.Y.; Seo, P.J.; Qiao, M.; Wang, N.; Li, S.; Cao, X.F.; Park, C.-M.; et al. WRKY 71 accelerates flowering via the direct activation of FLOWERING LOCUS T and LEAFY in Arabidopsis thaliana. Plant J. 2016, 85, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Chen, L.G.; Yu, D.Q. Transcription factor WRKY75 interacts with DELLA proteins to affect flowering. Plant Physiol. 2018, 176, 790–803. [Google Scholar] [CrossRef]
- Ma, Z.B.; Li, W.; Wang, H.P.; Yu, D.Q. WRKY transcription factors WRKY12 and WRKY13 interact with SPL10 to modulate age-mediated flowering. Integr. Plant Biol. 2020, 62, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Qi, S.L.; Touqeer, A.; Li, H.Y.; Zhang, X.L.; Liu, X.F.; Wu, S. SlGT11 controls floral organ patterning and floral determinacy in tomato. BMC Plant Biol. 2020, 20, 562. [Google Scholar] [CrossRef]
- Iván, F.A.; Hélène, L.; Sandra, P.R.; Eric, S.; Mats, H.; Mottinger, J.P.; Moreno, M.A.; Dellaporta, S.L. Tasselseed1 is a Lipoxygenase Affecting Jasmonic Acid Signaling in Sex Determination of Maize. Science 2009, 323, 262–265. [Google Scholar]
- Yadav, S.R.; Khanday, I.; Majhi, B.B.; Veluthambi, K.; Usha, V. Auxin-responsive OsMGH3, a common downstream target of OsMADS1 and OsMADS6, controls rice floret fertility. Plant Cell Physiol. 2011, 52, 2123–2135. [Google Scholar] [CrossRef]
- Huang, H.; Gao, H.; Liu, B.; Qi, T.C.; Tong, J.H.; Xiao, L.T.; Xie, D.X.; Song, S.S. Arabidopsis MYB24 regulates jasmonate-mediated stamen development. Front. Plant Sci. 2017, 8, 1525. [Google Scholar] [CrossRef]
- Niwa, T.; Suzuki, T.; Takebayashi, Y.; Ishiguro, R.; Higashiyama, T.; Sakakibara, H.; Ishiguro, S. Jasmonic acid facilitates flower opening and floral organ development through the upregulated expression of SlMYB21 transcription factor in tomato. Biosci. Biotechnol. Biochem. 2018, 82, 292–303. [Google Scholar] [CrossRef]
- Mandaokar, A.; Browse, J. MYB108 acts together with MYB24 to regulate jasmonate-mediated stamen maturation in Arabidopsis. Plant Physiol. 2009, 149, 851–862. [Google Scholar] [CrossRef]
- Sun, B.M.; Zhu, Z.S.; Chen, C.G.; Chen, G.J.; Cao, B.H.; Chen, C.M.; Lei, J.J. Jasmonate inducible R2R3-MYB transcription factor regulates capsaicinoid biosynthesis and stamen development in Capsicum. J. Agric. Food Chem. 2019, 67, 10891–10903. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xue, J.S.; Yu, Y.H.; Liu, S.Q.; Zhang, J.X.; Yao, X.Z.; Liu, Z.X.; Xu, X.F.; Yang, Z.N. Fine regulation of ARF17 for another development and pollen formation. BMC Plant Biol. 2017, 17, 243. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Wang, B.; Feng, Y.F.; Xue, J.S.; Qian, X.X.; Liu, S.Q.; Zhou, J.; Yu, Y.H.; Yang, N.Y.; Xu, P.; et al. AUXIN RESPONSE FACTOR17 directly regulates MYB108 for anther dehiscence. Plant Physiol. 2019, 181, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Coleman, H.D.; Beamish, L.; Reid, A.; Park, J.-Y.; Mansfield, S.D. Altered sucrose metabolism impacts plant biomass production and flower development. Transgenic Res. 2010, 19, 269–283. [Google Scholar] [CrossRef]
- Jin, S.; Kim, S.Y.; Ahn, J.H. Twin sister of FT (TSF) interacts with FRUCTOKINASE6 and inhibits its kinase activity in Arabidopsis. Front. Plant Sci. 2017, 8, 1807. [Google Scholar] [CrossRef]
- Wahl, V.; Ponnu, J.; Schlereth, A.; Arrivault, S.; Langenecker, T.; Franke, A.; Feil, R.; John, E.L.; Mark, S.; Markus, S. Regulation of Flowering by Trehalose-6-Phosphate Signaling in Arabidopsis thaliana. Science 2013, 339, 704–707. [Google Scholar] [CrossRef]
- Xu, F.Q.; Li, X.R.; Ruan, Y.L. RNAi-mediated suppression of nexokinasegene OsHXK10 in rice leads to non dehiscent anther and reduction of pollen germination. Plant Sci. 2008, 175, 674–684. [Google Scholar] [CrossRef]
- David-Schwartz, R.; Liron, W.; Roee, V.; Hanita, Z.; Leonid, M.; Dvora, S.; David, G. The SlFRK4 promoter is active only during late stages of pollen and anther development. Plant Sci. 2013, 199, 61–70. [Google Scholar] [CrossRef]
- Sivitz, A.B.; Reinders, A.; Ward, J.M. Arabidopsis sucrose transporter AtSUC1 is important for pollen germination and sucrose-induced anthocyanin accumulation. Plant Physiol. 2008, 147, 92–100. [Google Scholar] [CrossRef]
- Goetz, M.; Guivarćh, A.; Hirsche, J.; Bauerfeind, M.A.; González, M.C.; Hyun, T.K.; Eom, S.H.; Chriqui, D.; Engelke, T.; Großkinsky, D.K.; et al. Metabolic control of tobacco pollination by sugars and invertases. Plant Physiol. 2017, 173, 984–997. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhang, L.; Yang, S. Analysis of Floral Organ Development and Sex Determination in Schisandra chinensis by Scanning Electron Microscopy and RNA-Sequencing. Life 2022, 12, 1260. https://doi.org/10.3390/life12081260
Liu X, Zhang L, Yang S. Analysis of Floral Organ Development and Sex Determination in Schisandra chinensis by Scanning Electron Microscopy and RNA-Sequencing. Life. 2022; 12(8):1260. https://doi.org/10.3390/life12081260
Chicago/Turabian StyleLiu, Xiuyan, Lifan Zhang, and Shihai Yang. 2022. "Analysis of Floral Organ Development and Sex Determination in Schisandra chinensis by Scanning Electron Microscopy and RNA-Sequencing" Life 12, no. 8: 1260. https://doi.org/10.3390/life12081260
APA StyleLiu, X., Zhang, L., & Yang, S. (2022). Analysis of Floral Organ Development and Sex Determination in Schisandra chinensis by Scanning Electron Microscopy and RNA-Sequencing. Life, 12(8), 1260. https://doi.org/10.3390/life12081260