
Circadian Rhythm Regulates Reactive Oxygen Species Production and Inhibits Al-Induced Programmed Cell Death in Peanut
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
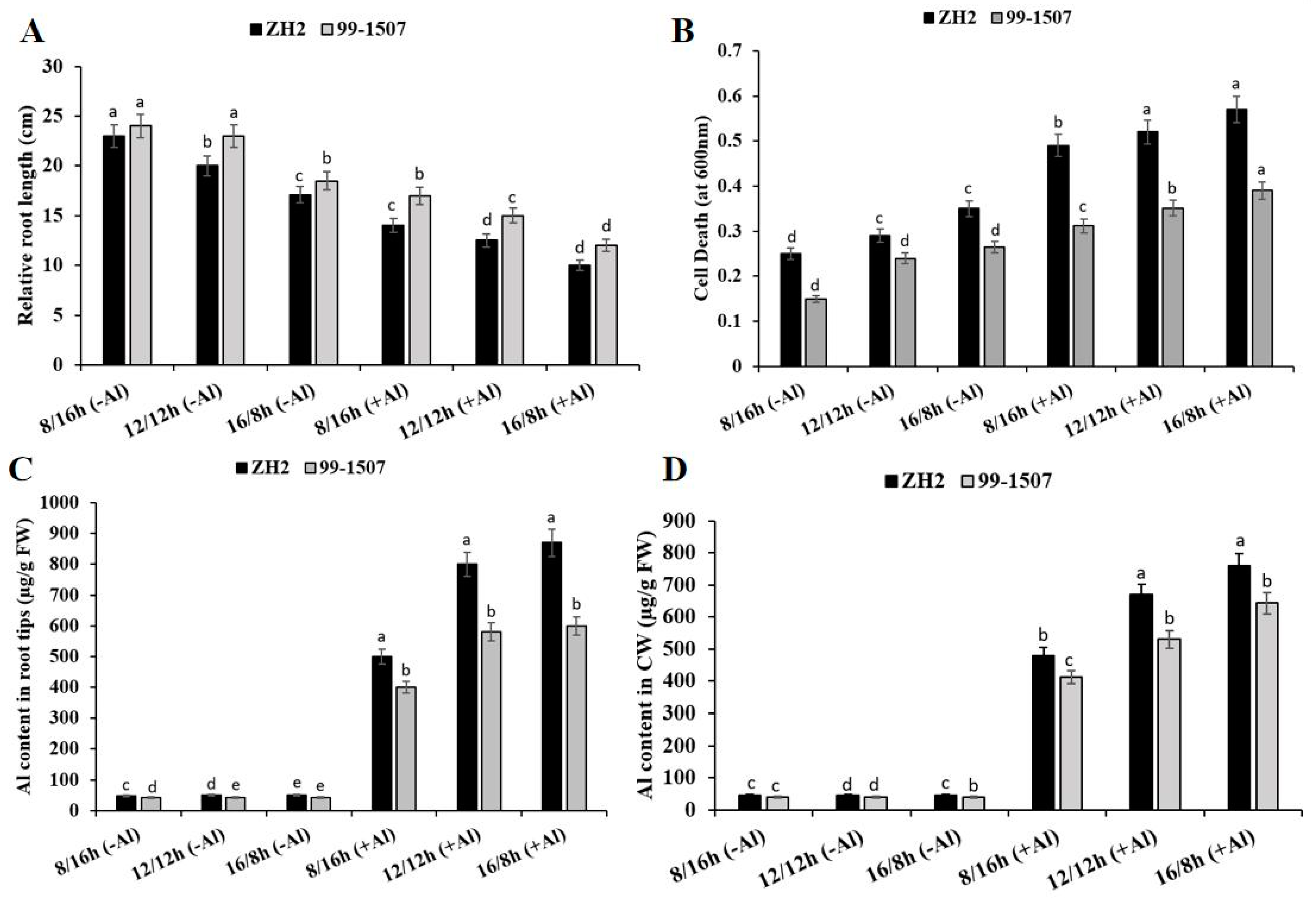
2.1. Effects of Photoperiod and Al Stress on Root Elongation and Cell Death
2.2. Effects of Photoperiod and Al Stress on Al Accumulation in the Root Tips and CW
2.3. Effects of Photoperiod and Al Stress on MDA Content, PME Activity, and LOX Activity
2.4. Effects of Photoperiod and Al on ROS Production and Antioxidant Enzyme Activities
2.5. Effect of Photoperiod on AhLHY and AhSTS Gene Expression under Al Stress
3. Discussion
3.1. Effects of Photoperiods on Root Elongation and Cell Death under Al Stress
3.2. Effect of Photoperiod and Al Stress on MDA, PME, and LOX Activity
3.3. Circadian Rhythm Regulates Al-Induced PCD by Controlling ROS Production and Antioxidant Enzyme Activity
3.4. Effect of Photoperiod and Al Stress on AhLHY and AhSTS Gene Expression
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Relative Root Growth, Evan’s Blue Staining, and Cell Death Assay
4.3. Al Content in Root Tips and Cell Wall
4.4. Lipid Peroxidation and PME Activity Assay
4.5. ROS (H2O2 and O2.−) Content
4.6. Antioxidant Enzyme Activity
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
| Gene Name | Gene ID | Primer |
| AhLHY-forward | AH02G04520.1 | 5′-ATTGACTCTAGTAATCGTCGTA-3′ |
| AhLHY-reverse | 5′-CTTTGTGGCAACACCTCT-3′ | |
| AhSTS-forward | AH04G08660.1 | 5′-CCCAAGCGTCAAGAGGTA-3′ |
| AhSTS-reverse | 5′-TTGCCCACAAGACTATCCA-3′ | |
| Actin-forward | AH03G02610.1 | 5′-ACCTTCTACAACGAGCTTCGTGTG-3′ |
| Actin-reverse | 5′-GAAAGAACAGCCTGAATGGCAAC-3′ |
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Panda, S.K.; Baluška, F.; Matsumoto, H. Aluminum stress signaling in plants. Plant Signal. Behav. 2009, 4, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Shetty, R.; Vidya, C.S.-N.; Prakash, N.B.; Lux, A.; Vaculík, M. Aluminum toxicity in plants and its possible mitigation in acid soils by biochar: A review. Sci. Total Environ. 2021, 765, 142744. [Google Scholar] [CrossRef] [PubMed]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, Y.; Zhang, J.; Shi, W.; Qian, C.; Peng, X. Identification of aluminum-responsive proteins in rice roots by a proteomic approach: Cysteine synthase as a key player in Al response. PROTEOMICS 2007, 7, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gao, H.; Wu, X.; Fang, Q.; Chen, L.; Zhao, F.-J.; Huang, C.-F. Isolation and Characterization of an Aluminum-resistant Mutant in Rice. Rice 2016, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Savić, J.; Stevic, N.; Maksimović, V.; Samardžić, J.; Nikolić, D.; Nikolić, M. Root malate efflux and expression of taalmt1 in serbian winter wheat cultivars differing in Al tolerance. J. Soil Sci. Plant Nutr. 2018, 18, 90–99. [Google Scholar] [CrossRef]
- Szurman-Zubrzycka, M.; Chwiałkowska, K.; Niemira, M.; Kwaśniewski, M.; Nawrot, M.; Gajecka, M.; Larsen, P.B.; Szarejko, I. Aluminum or Low pH—Which Is the Bigger Enemy of Barley? Transcriptome Analysis of Barley Root Meristem Under Al and Low pH Stress. Front. Genet. 2021, 12, 675260. [Google Scholar] [CrossRef]
- Zhou, G.; Delhaize, E.; Zhou, M.; Ryan, P.R. Biotechnological solutions for enhancing the aluminium resistance of crop plants. In Abiotic Stress in Plants—Mechanisms and Adaptations; IntechOpen: London, UK, 2011; pp. 119–142. [Google Scholar]
- Ninamango-Cárdenas, F.E.; Guimaraes, C.T.; Martins, P.R.; Parentoni, S.N.; Carneiro, N.P.; Lopes, M.A.; Moro, J.R.; Paiva, E. Mapping QTLs for aluminum tolerance in maize. Euphytica 2003, 130, 223–232. [Google Scholar] [CrossRef]
- Mattiello, L.; da Silva, F.R.; Menossi, M. Linking microarray data to QTLs highlights new genes related to Al tolerance in maize. Plant Sci. 2012, 191–192, 8–15. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Koyama, H. QTL analysis of Al tolerance in recombinant inbred lines of Arabidopsis thaliana. Plant Cell Physiol. 2002, 43, 1526–1533. [Google Scholar] [CrossRef]
- Farokhzadeh, S.; Fakheri, B.A.; Nezhad, N.M.; Tahmasebi, S.; Mirsoleimani, A. Mapping QTLs of flag leaf morphological and physiological traits related to aluminum tolerance in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2019, 25, 975–990. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.D.; Brar, D.S.; Bui, B.C.; Nguyen, T.V.; Pham, L.N.; Nguyen, H.T. Identification and mapping of the QTL for aluminum tolerance introgressed from the new source, ORYZA RUFIPOGON Griff., into indica rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 106, 583–593. [Google Scholar] [CrossRef]
- Sade, H.; Meriga, B.; Surapu, V.; Gadi, J.; Sunita, M.S.L.; Suravajhala, P.; Kishor, P.B.K. Toxicity and tolerance of aluminum in plants: Tailoring plants to suit to acid soils. BioMetals 2016, 29, 187–210. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, H.; He, L.-F. Genome-wide analysis of the MATE gene family in potato. Mol. Biol. Rep. 2018, 46, 403–414. [Google Scholar] [CrossRef]
- Augusto, A.R.; Yang, B.; de Oliveira, V.; Felipe, L.; Lauro, B.N.; Schunemann, M.; dos Santos, M.F.; Mariath, J.; Silveiro, A.; Sachetto-Martins, G.; et al. New Insights into Aluminum Tolerance in Rice: The ASR5 Protein Binds the STAR1 Promoter and Other Aluminum-Responsive Genes. Mol. Plant 2014, 7, 709–721. [Google Scholar] [CrossRef]
- Zhaorong, H.; Tian, X.; Wang, F.; Zhang, L.; Xin, M.; Hu, Z.; Yao, Y.; Ni, Z.; Sun, Q.; Peng, H. Characterization of wheat MYB genes responsive to high temperatures. BMC Plant Biol. 2017, 17, 208. [Google Scholar] [CrossRef]
- Caniato, F.F.; Guimarães, C.T.; Schaffert, R.E.; Alves, V.M.C.; Kochian, L.V.; Borém, A.; Klein, P.E.; Magalhaes, J.V. Genetic diversity for aluminum tolerance in sorghum. Theor. Appl. Genet. 2007, 114, 863–876. [Google Scholar] [CrossRef]
- Wakao, S.; Niyogi, K.K. Chlamydomonas as a model for reactive oxygen species signaling and thiol redox regulation in the green lineage. Plant Physiol. 2021, 187, 687–698. [Google Scholar] [CrossRef]
- Watt, D.A. Aluminium-responsive genes in sugarcane: Identification and analysis of expression under oxidative stress. J. Exp. Bot. 2003, 54, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Guo, J.; Zhou, F.; Singh, S.; Xu, X.; Xie, Q.; Yang, Z.; Huang, C.-F. F-box protein RAE1 regulates the stability of the aluminum-resistance transcription factor STOP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 116, 319–327. [Google Scholar] [CrossRef]
- Fang, Q.; Zhou, F.; Zhang, Y.; Singh, S.; Huang, C.-F. Degradation of STOP1 mediated by the F-box proteins RAH1 and RAE1 balances aluminum resistance and plant growth in Arabidopsis thaliana. Plant J. 2021, 106, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Ohyama, Y.; Kobayashi, Y.; Ito, H.; Iuchi, S.; Fujita, M.; Zhao, C.-R.; Tanveer, T.; Ganesan, M.; Kobayashi, M.; et al. STOP2 Activates Transcription of Several Genes for Al- and Low pH-Tolerance that Are Regulated by STOP1 in Arabidopsis. Mol. Plant 2014, 7, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Gong, Z.; Chen, X.; Wang, H.; Tan, R.; Mao, Y. Transcriptomic responses to aluminum stress in tea plant leaves. Sci. Rep. 2021, 11, 5800. [Google Scholar] [CrossRef] [PubMed]
- Hardin, P.E.; Panda, S. Circadian timekeeping and output mechanisms in animals. Curr. Opin. Neurobiol. 2013, 23, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.; Shin, J.; Davis, S.J. Abiotic stress and the plant circadian clock. Plant Signal. Behav. 2011, 6, 223–231. [Google Scholar] [CrossRef]
- Shalit-Kaneh, A.; Kumimoto, R.W.; Filkov, V.; Harmer, S.L. Multiple feedback loops of the Arabidopsis circadian clock provide rhythmic robustness across environmental conditions. Proc. Natl. Acad. Sci. USA 2018, 115, 7147–7152. [Google Scholar] [CrossRef]
- Green, R.M.; Tingay, S.; Wang, Z.-Y.; Tobin, E.M. Circadian Rhythms Confer a Higher Level of Fitness to Arabidopsis Plants. Plant Physiol. 2002, 129, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Y.; He, Y.; Wang, Y.; Wang, L. Pseudo Response Regulators Regulate Photoperiodic Hypocotyl Growth by Repressing PIF4/5 Transcription. Plant Physiol. 2020, 183, 686–699. [Google Scholar] [CrossRef] [PubMed]
- Mase, K.; Tsukagoshi, H. Reactive Oxygen Species Link Gene Regulatory Networks during Arabidopsis Root Development. Front. Plant Sci. 2021, 12, 660274. [Google Scholar] [CrossRef]
- Spoel, S.H.; Van Ooijen, G. Circadian redox signaling in plant immunity and abiotic stress. Antioxid. Redox Signal. 2014, 20, 3024–3039. [Google Scholar] [CrossRef]
- Egrundy, J.; Estoker, C.; Carre, I.A. Circadian regulation of abiotic stress tolerance in plants. Front. Plant Sci. 2015, 6, 648. [Google Scholar] [CrossRef]
- Mishra, A.P.; Salehi, B.; Sharifi-Rad, M.; Pezzani, R.; Kobarfard, F.; Sharifi-Rad, J.; Nigam, M. Programmed Cell Death, from a Cancer Perspective: An Overview. Mol. Diagn. Ther. 2018, 22, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Li, X.; Zhou, Y.-Y.; Wei, L.; Keovongkod, C.; He, H.-Y.; Zhan, J.; Wang, A.-Q.; He, L.-F. Transcriptome analysis reveals significant difference in gene expression and pathways between two peanut cultivars under Al stress. Gene 2021, 781, 145535. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.; Shi, Y.; Ntambiyukuri, A.; Li, X.; Zhan, J.; Wang, A.; Xiao, D.; He, L. Integration of Small RNA and Degradome Sequencing Reveals the Regulatory Network of Al-Induced Programmed Cell Death in Peanut. Int. J. Mol. Sci. 2021, 23, 246. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Roeber, V.M.; Schmülling, T.; Cortleven, A. The Photoperiod: Handling and Causing Stress in Plants. Front. Plant Sci. 2022, 12, 781988. [Google Scholar] [CrossRef]
- Nitschke, S.; Cortleven, A.; Iven, T.; Feussner, I.; Havaux, M.; Riefler, M.; Schmülling, T. Circadian Stress Regimes Affect the Circadian Clock and Cause Jasmonic Acid-Dependent Cell Death in Cytokinin-Deficient Arabidopsis Plants. Plant Cell 2016, 28, 1616–1639. [Google Scholar] [CrossRef]
- Wang, T.A.; Yu, Y.V.; Govindaiah, G.; Ye, X.; Artinian, L.; Coleman, T.P.; Sweedler, J.V.; Cox, C.L.; Gillette, M.U. Circadian Rhythm of Redox State Regulates Excitability in Suprachiasmatic Nucleus Neurons. Science 2012, 337, 839–842. [Google Scholar] [CrossRef]
- Hirayama, J.; Cho, S.; Sassone-Corsi, P. Circadian control by the reduction/oxidation pathway: Catalase represses light-dependent clock gene expression in the zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 15747–15752. [Google Scholar] [CrossRef]
- Shim, J.S.; Imaizumi, T. Circadian Clock and Photoperiodic Response in Arabidopsis: From Seasonal Flowering to Redox Homeostasis. Biochemistry 2014, 54, 157–170. [Google Scholar] [CrossRef]
- Lai, A.G.; Doherty, C.J.; Mueller-Roeber, B.; Kay, S.A.; Schippers, J.H.M.; Dijkwel, P.P. CIRCADIAN CLOCK-ASSOCIATED 1 regulates ROS homeostasis and oxidative stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 17129–17134. [Google Scholar] [CrossRef] [PubMed]
- Abuelsoud, W.; Cortleven, A.; Schmülling, T. Photoperiod stress induces an oxidative burst-like response and is associated with increased apoplastic peroxidase and decreased catalase activities. J. Plant Physiol. 2020, 253, 153252. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.-J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-Z.; Gong, Z.-Q.; Zhang, Y.-L.; Li, P.-J. Growth, Cadmium Accumulation and Physiology of Marigold (Tagetes erecta L.) as Affected by Arbuscular Mycorrhizal Fungi. Pedosphere 2011, 21, 319–327. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Chen, Z.-S.; Hong, C.-Y. Cadmium-induced physiological response and antioxidant enzyme changes in the novel cadmium accumulator, Tagetes patula. J. Hazard. Mater. 2011, 189, 724–731. [Google Scholar] [CrossRef]
- Uraguchi, S.; Watanabe, I.; Yoshitomi, A.; Kiyono, M.; Kuno, K. Characteristics of cadmium accumulation and tolerance in novel Cd-accumulating crops, Avena strigosa and Crotalaria juncea. J. Exp. Bot. 2006, 57, 2955–2965. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef]
- Ahn, H.R.; Kim, Y.-J.; Lim, Y.J.; Duan, S.; Eom, S.H.; Jung, K.-H. Key Genes in the Melatonin Biosynthesis Pathway with Circadian Rhythm Are Associated with Various Abiotic Stresses. Plants 2021, 10, 129. [Google Scholar] [CrossRef]
- Xiang, Y.; Sapir, T.; Rouillard, P.; Ferrand, M.; Jiménez-Gómez, J.M. Interaction between photoperiod and variation in circadian rhythms in tomato. BMC Plant Biol. 2022, 22, 187. [Google Scholar] [CrossRef]
- Guo, R.; Li, W.; Wang, X.; Chen, B.; Huang, Z.; Liu, T.; Chen, X.; XuHan, X.; Lai, Z. Effect of photoperiod on the formation of cherry radish root. Sci. Hortic. 2018, 244, 193–199. [Google Scholar] [CrossRef]
- Osnato, M.; Cota, I.; Nebhnani, P.; Cereijo, U.; Pelaz, S. Photoperiod Control of Plant Growth: Flowering Time Genes beyond Flowering. Front. Plant Sci. 2022, 12, 805635. [Google Scholar] [CrossRef] [PubMed]
- Duressa, D.; Soliman, K.; Chen, D. Identification of Aluminum Responsive Genes in Al-Tolerant Soybean Line PI 416937. Int. J. Plant Genom. 2010, 2010, 164862. [Google Scholar] [CrossRef] [PubMed]
- Riboni, M.; Test, A.R.; Galbiati, M.; Tonelli, C.; Conti, L. Environmental stress and flowering time: The photoperiodic connection. Plant Signal. Behav. 2014, 9, e29036. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.; Cortleven, A.; Novák, O.; Schmülling, T. Root-derived trans -zeatin cytokinin protects Arabidopsis plants against photoperiod stress. Plant Cell Environ. 2020, 43, 2637–2649. [Google Scholar] [CrossRef]
- Fortini, E.A.; Batista, D.S.; De Castro, K.M.; Silva, T.D.; Felipe, S.H.S.; Correia, L.N.F.; Chagas, K.; Farias, L.M.; Leite, J.P.V.; Otoni, W.C. Photoperiod modulates growth and pigments and 20-hydroxyecdysone accumulation in Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen] grown in vitro. Plant Cell Tissue Organ Cult. 2020, 142, 595–611. [Google Scholar] [CrossRef]
- He, H.; Oo, T.L.; Huang, W.; He, L.-F.; Gu, M. Nitric oxide acts as an antioxidant and inhibits programmed cell death induced by aluminum in the root tips of peanut (Arachis hypogaea L.). Sci. Rep. 2019, 9, 9516. [Google Scholar] [CrossRef]
- Laczko, R.; Csiszar, K. Lysyl Oxidase (LOX): Functional Contributions to Signaling Pathways. Biomolecules 2020, 10, 1093. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef]
- Sun, C.; Lu, L.; Yu, Y.; Liu, L.; Hu, Y.; Ye, Y.; Jin, C.; Lin, X. Decreasing methylation of pectin caused by nitric oxide leads to higher aluminium binding in cell walls and greater aluminium sensitivity of wheat roots. J. Exp. Bot. 2015, 67, 979–989. [Google Scholar] [CrossRef]
- Darley, C.P.; Forrester, A.M.; McQueen-Mason, S.J. The molecular basis of plant cell wall extension. Plant Mol. Biol. 2001, 47, 179–195. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2020, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Krasensky-Wrzaczek, J.; Kangasjärvi, J. The role of reactive oxygen species in the integration of temperature and light signals. J. Exp. Bot. 2018, 69, 3347–3358. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.A.; Wakin, T.; Batista-Silva, W.; Silva, J.C.F.; Vicente, M.H.; Silva, J.C.; Clarindo, W.R.; Zsögön, A.; Peres, L.E.; De Veylder, L.; et al. A long and stressful day: Photoperiod shapes aluminium tolerance in plants. J. Hazard. Mater. 2022, 432, 128704. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Ito, S.; Imaizumi, T. Similarities in the circadian clock and photoperiodism in plants. Curr. Opin. Plant Biol. 2010, 13, 594–603. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Schmidt, M.; Grief, J.; Feierabend, J. Mode of transcreiptional activation of the catalase (cat1) mRNA of rye leaves (Secale cereale L.) and its control through blue light and reactive oxygen. Planta 2006, 223, 835–846. [Google Scholar] [CrossRef]
- Hertwig, B.; Streb, P.; Feierabend, J. Light Dependence of Catalase Synthesis and Degradation in Leaves and the Influence of Interfering Stress Conditions. Plant Physiol. 1992, 100, 1547–1553. [Google Scholar] [CrossRef]
- Park, M.-J.; Kwon, Y.-J.; Gil, K.-E.; Park, C.-M. LATE ELONGATED HYPOCOTYL regulates photoperiodic flowering via the circadian clock in Arabidopsis. BMC Plant Biol. 2016, 16, 114. [Google Scholar] [CrossRef]
- Markham, K.K.; Greenham, K. Abiotic stress through time. N. Phytol. 2021, 231, 40–46. [Google Scholar] [CrossRef]
- Bagnall, D.; King, R. Response of peanut (Arachis hypogaea) to temperature, photoperiod and irradiance 1. Effect on flowering. Field Crop. Res. 1991, 26, 263–277. [Google Scholar] [CrossRef]
- Pan, C.-L.; Yao, S.-C.; Xiong, W.-J.; Luo, S.-Z.; Wang, Y.-L.; Wang, A.-Q.; Xiao, D.; Zhan, J.; He, L.-F. Nitric Oxide Inhibits Al-Induced Programmed Cell Death in Root Tips of Peanut (Arachis hypogaea L.) by Affecting Physiological Properties of Antioxidants Systems and Cell Wall. Front. Physiol. 2017, 8, 1037. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, R.; Wan, Q.; Xie, G.; Bi, Y. Glucose-6-Phosphate Dehydrogenase Plays a Pivotal Role in Nitric Oxide-Involved Defense Against Oxidative Stress Under Salt Stress in Red Kidney Bean Roots. Plant Cell Physiol. 2007, 48, 511–522. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntambiyukuri, A.; Li, X.; Xiao, D.; Wang, A.; Zhan, J.; He, L. Circadian Rhythm Regulates Reactive Oxygen Species Production and Inhibits Al-Induced Programmed Cell Death in Peanut. Life 2022, 12, 1271. https://doi.org/10.3390/life12081271
Ntambiyukuri A, Li X, Xiao D, Wang A, Zhan J, He L. Circadian Rhythm Regulates Reactive Oxygen Species Production and Inhibits Al-Induced Programmed Cell Death in Peanut. Life. 2022; 12(8):1271. https://doi.org/10.3390/life12081271
Chicago/Turabian StyleNtambiyukuri, Aaron, Xia Li, Dong Xiao, Aiqin Wang, Jie Zhan, and Longfei He. 2022. "Circadian Rhythm Regulates Reactive Oxygen Species Production and Inhibits Al-Induced Programmed Cell Death in Peanut" Life 12, no. 8: 1271. https://doi.org/10.3390/life12081271
APA StyleNtambiyukuri, A., Li, X., Xiao, D., Wang, A., Zhan, J., & He, L. (2022). Circadian Rhythm Regulates Reactive Oxygen Species Production and Inhibits Al-Induced Programmed Cell Death in Peanut. Life, 12(8), 1271. https://doi.org/10.3390/life12081271