
Structural Insights into Mouse H-FABP

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Protein Expression and Purification
2.3. Crystallization
2.4. Structure Determination and Comparison
2.5. NMR Experiments and Data Processing
3. Results and Discussion
3.1. Crystal Structure of Mouse H-FABP
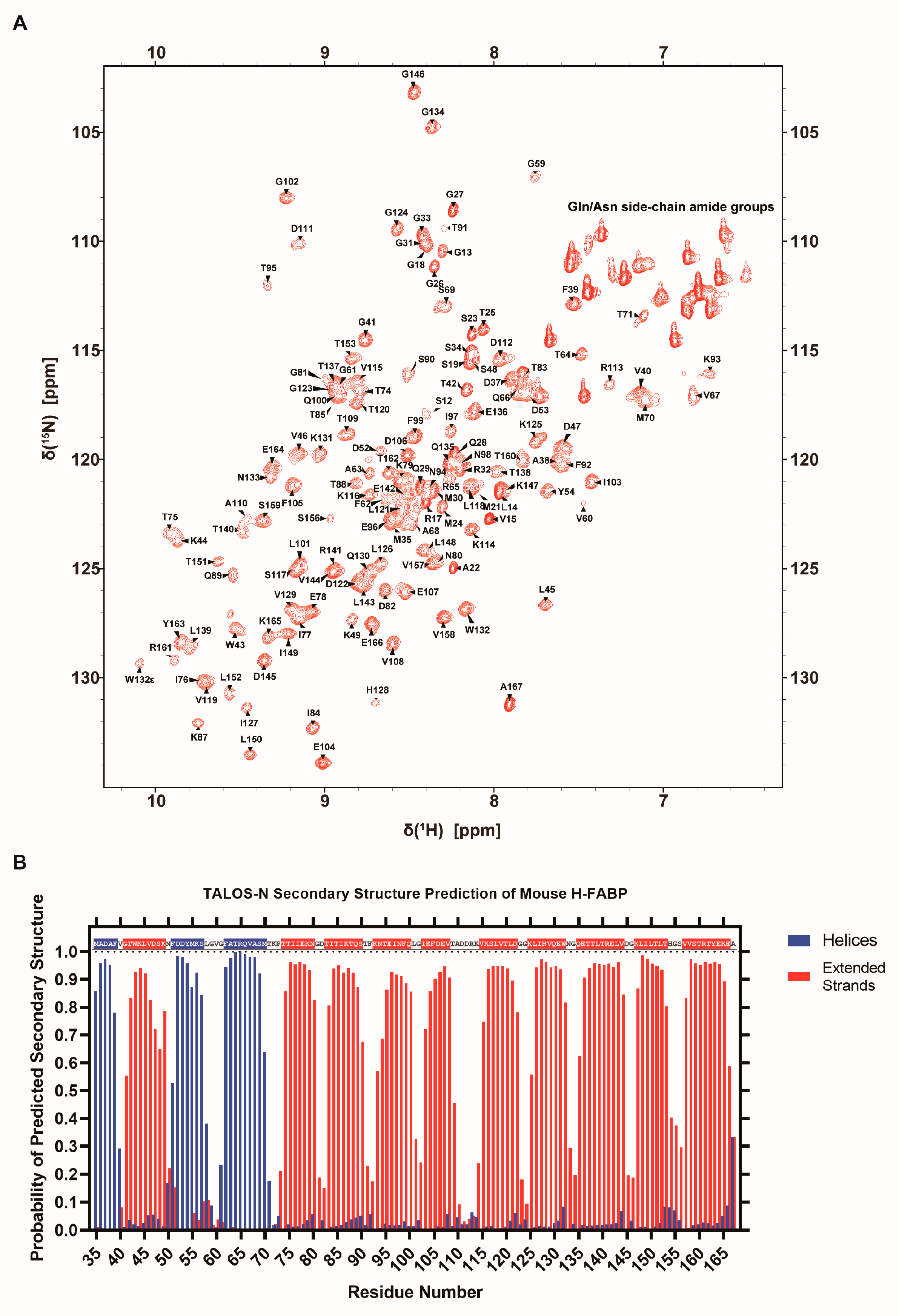
3.2. NMR Assignment and Secondary Structure Prediction of Mouse H-FABP
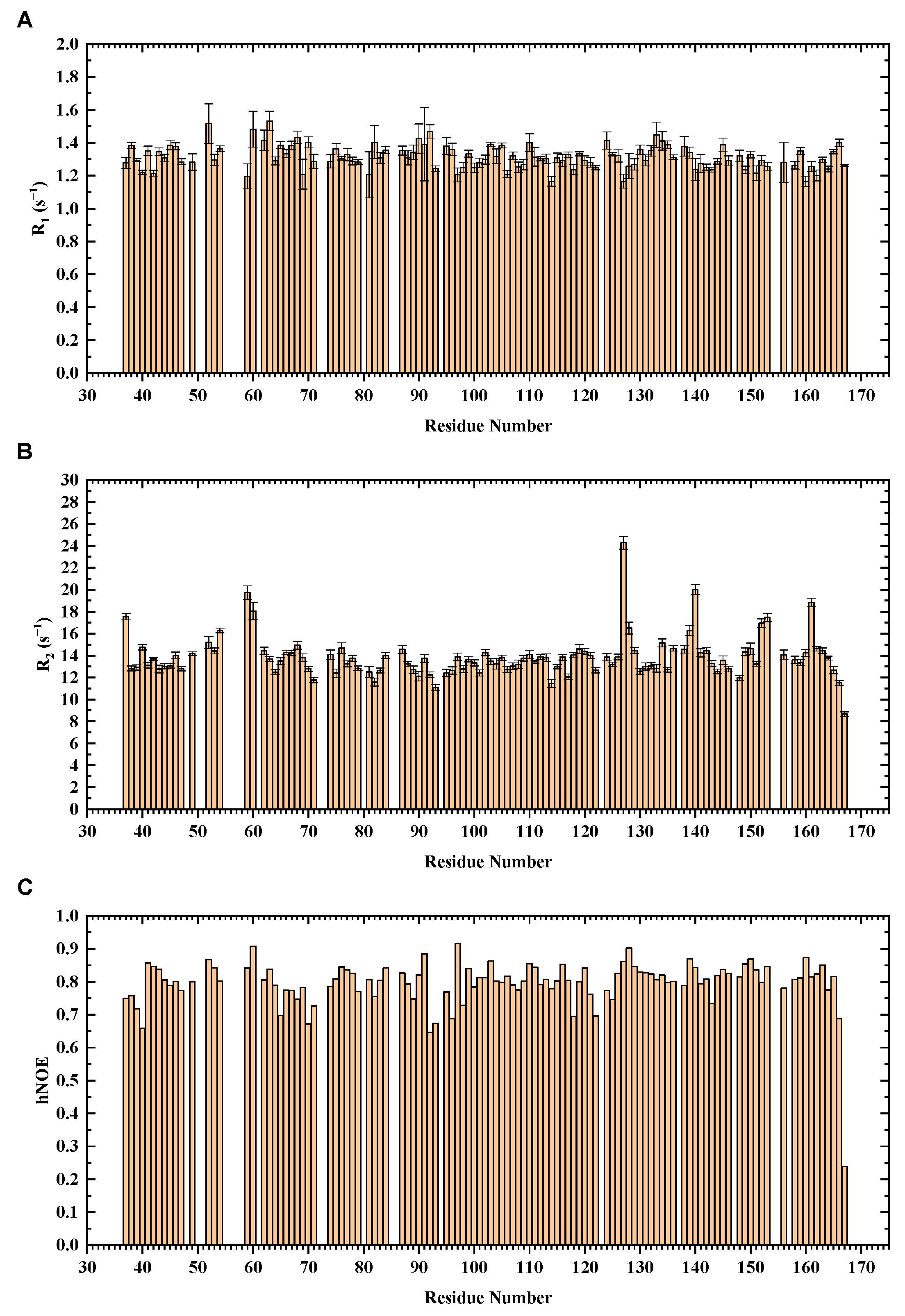
3.3. NMR Dynamics of Mouse H-FABP
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Veerkamp, J.H.; Maatman, R.G.H.J. Cytoplasmic fatty acid-binding proteins: Their structure and genes. Prog. Lipid Res. 1995, 34, 17–52. [Google Scholar] [CrossRef]
- Veerkamp, J.H.; Peeters, R.A.; Maatman, R.G.H.J. Structural and functional features of different types of cytoplasmic fatty ac-id-binding proteins. Biochim. Biophys. Acta Lipids Lipid Metab. 1991, 1081, 1–24. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; van Nieuwenhoven, F.A.; Luiken, J.J.F.P.; Schaap, F.G.; van der Vusse, G.J. Role of membrane-associated and cytoplasmic fatty acid-binding proteins in cellular fatty acid metabolism. Prostaglandins Leukot. Essent. Fat. Acids 1997, 57, 373–378. [Google Scholar] [CrossRef]
- Veerkamp, J.H.; Paulussen, R.J.A.; Peeters, R.A.; Maatman, R.G.H.J.; van Moerkerk, H.T.B.; van Kuppevelt, T.H.M.S.M. Detection, tissue distribution and (sub)cellular localization of fatty acid-binding protein types. Mol. Cell. Biochem. 1990, 98, 11–18. [Google Scholar] [CrossRef]
- Zschiesche, W.; Kleine, A.H.; Spitzer, E.; Veerkamp, J.H.; Glatz, J.F.C. Histochemical localization of heart-type fatty-acid binding protein in human and murine tissues. Histochem. Cell Biol. 1995, 103, 147–156. [Google Scholar] [CrossRef]
- Watanabe, K.; Wakabayashi, H.; Veerkamp, J.H.; Ono, T.; Suzuki, T. Immunohistochemical distribution of heart-type fatty acid-binding protein immunoreactivity in normal human tissues and in acute myocardial infarct. J. Pathol. 1993, 170, 59–65. [Google Scholar] [CrossRef]
- Storch, J.; Corsico, B. The Emerging Functions and Mechanisms of Mammalian Fatty Acid–Binding Proteins. Annu. Rev. Nutr. 2008, 28, 73–95. [Google Scholar] [CrossRef]
- Toke, O. Structural and Dynamic Determinants of Molecular Recognition in Bile Acid-Binding Proteins. Int. J. Mol. Sci. 2022, 23, 505. [Google Scholar] [CrossRef]
- Xu, H.; Diolintzi, A.; Storch, J. Fatty acid-binding proteins: Functional understanding and diagnostic implications. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 407–412. [Google Scholar] [CrossRef]
- Storch, J.; McDermott, L. Structural and functional analysis of fatty acid-binding proteins. J. Lipid Res. 2009, 50, S126–S131. [Google Scholar] [CrossRef] [Green Version]
- Storch, J.; Thumser, A.E.A. The fatty acid transport function of fatty acid-binding proteins. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1486, 28–44. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; van der Vusse, G.J. Cellular fatty acid-binding proteins: Their function and physiological significance. Prog. Lipid Res. 1996, 35, 243–282. [Google Scholar] [CrossRef]
- Niu, Y.-G.; Hauton, D.; Evans, R.D. Utilization of triacylglycerol-rich lipoproteins by the working rat heart: Routes of uptake and metabolic fates. J. Physiol. 2004, 558, 225–237. [Google Scholar] [CrossRef]
- Richieri, G.V.; Ogata, R.T.; Zimmerman, A.W.; Veerkamp, J.H.; Kleinfeld, A.M. Fatty Acid Binding Proteins from Different Tissues Show Distinct Patterns of Fatty Acid Interactions. Biochemistry 2000, 39, 7197–7204. [Google Scholar] [CrossRef]
- Smathers, R.L.; Petersen, D.R. The human fatty acid-binding protein family: Evolutionary divergences and functions. Hum. Genom. 2011, 5, 170–191. [Google Scholar] [CrossRef] [PubMed]
- Hanhoff, T.; Lücke, C.; Spener, F. Insights into binding of fatty acids by fatty acid binding proteins. Mol. Cell Biochem. 2002, 239, 45–54. [Google Scholar] [CrossRef]
- O’Sullivan, S.E.; Kaczocha, M. FABP5 as a novel molecular target in prostate cancer. Drug Discov. Today 2020, 25, 2056–2061. [Google Scholar] [CrossRef]
- Berger, W.T.; Ralph, B.P.; Kaczocha, M.; Sun, J.; Balius, T.E.; Rizzo, R.C.; Haj-Dahmane, S.; Ojima, I.; Deutsch, D.G. Targeting fatty acid binding protein (FABP) anandamide transporters—A novel strategy for development of anti-inflammatory and anti-nociceptive drugs. PLoS ONE 2012, 7, e50968. [Google Scholar] [CrossRef]
- Goel, H.; Melot, J.; Krinock, M.D.; Kumar, A.; Nadar, S.K.; Lip, G.Y.H. Heart-type fatty acid-binding protein: An overlooked cardiac biomarker. Ann. Med. 2020, 52, 444–461. [Google Scholar]
- Burton, P.B.J.; Hogben, C.E.; Joannou, C.L.; Clark, A.G.B.; Hsuan, J.J.; Totty, N.F.; Sorensen, C.; Evans, R.W.; Tynan, M.J. Heart Fatty Acid Binding Protein Is a Novel Regulator of Cardiac Myocyte Hypertrophy. Biochem. Biophys. Res. Commun. 1994, 205, 1822–1828. [Google Scholar] [CrossRef]
- Vaidya, V.S.; Ferguson, M.A.; Bonventre, J.V. Biomarkers of acute kidney injury. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 463–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, H.T.; Larsson, C.; Narod, S.; Pollak, M. Tumor suppressor activity of the gene encoding mammary-derived growth inhibitor. Cancer Res. 1995, 55, 2225–2231. [Google Scholar] [PubMed]
- Pritt, M.L.; Hall, D.G.; Recknor, J.; Credille, K.M.; Brown, D.D.; Yumibe, N.P.; Schultze, A.E.; Watson, D.E. Fabp3 as a Biomarker of Skeletal Muscle Toxicity in the Rat: Comparison with Conventional Biomarkers. Toxicol. Sci. 2008, 103, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Lee, S.H.; Jung, Y.; Lee, Y.; Yoon, J.H.; Choi, J.Y.; Hwang, C.Y.; Son, Y.H.; Park, S.S.; Hwang, G.S.; et al. FABP3-mediated membrane lipid saturation alters fluidity and induces ER stress in skeletal muscle with aging. Nat. Commun. 2020, 11, 5661. [Google Scholar] [CrossRef]
- Chiasserini, D.; Biscetti, L.; Eusebi, P.; Salvadori, N.; Frattini, G.; Simoni, S.; de Roeck, N.; Tambasco, N.; Stoops, E.; Vanderstichele, H.; et al. Differential role of CSF fatty acid binding protein 3, α-synuclein, and Alzheimer’s disease core biomarkers in Lewy body disorders and Alzheimer’s dementia. Alzheimer’s Res. Ther. 2017, 9, 52. [Google Scholar] [CrossRef]
- Lücke, C.; Rademacher, M.; Zimmerman, A.W.; van Moerkerk, H.T.B.; Veerkamp, J.H.; Rüter-Jans, H. Spin-system heterogeneities indicate a selected-fit mechanism in fatty acid binding to heart-type fatty acid-binding protein (H-FABP). Biochem. J. 2001, 354, 259–266. [Google Scholar] [CrossRef]
- Young, A.C.M.; Scapin, G.; Kromminga, A.; Patel, S.B.; Veerkamp, J.H.; Sacchettini, J.C. Structural studies on human muscle fatty acid binding protein at 1.4 å resolution: Binding interactions with three C18 fatty acids. Structure 1994, 2, 523–534. [Google Scholar] [CrossRef]
- Lassen, D.; Lücke, C.; Kveder, M.; Mesgarzadeh, A.; Schmidt, J.M.; Specht, B.; Lezius, A.; Spener, F.; Rüterjans, H. Three-dimensional structure of bovine heart fatty-acid-binding protein with bound palmitic acid, determined by multidimen-sional NMR spectroscopy. Eur. J. Biochem. 1995, 230, 266–280. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallographica. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System; Delano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Rahimi, M.; Lee, Y.; Chiu, A. POKY: A software suite for multidimensional NMR and 3D structure calculation of biomolecules. Bioinformatics 2021, 37, 3041–3042. [Google Scholar] [CrossRef] [PubMed]
- Kawale, A.A.; Burmann, B.M. Characterization of backbone dynamics using solution NMR spectroscopy to discern the functional plasticity of structurally analogous proteins. STAR Protoc. 2021, 2, 100919. [Google Scholar] [CrossRef]
- Rossi, P.; Swapna, G.V.T.; Huang, Y.J.; Aramini, J.M.; Anklin, C.; Conover, K.; Hamilton, K.; Xiao, R.; Acton, T.B.; Ertekin, A.; et al. A microscale protein NMR sample screening pipeline. J. Biomol. NMR 2010, 46, 11–22. [Google Scholar] [CrossRef]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection Statistics | |
| Wavelength (Å) | 1.54178 |
| Space group | P212121 |
| Unit-Cell parameters (Å, °) | |
| a/b/c | a = 37.3050, b = 54.4190, c = 65.3870 |
| α/β/γ | α = 90.00, β = 90.00, γ = 90.00 |
| Resolution range (out shell) (Å) | 41.83–1.7 (1.761–1.7) |
| No. of reflections | 17,298 (1585) |
| No. of unique reflections | 15,182 (1456) |
| Wilson B-factor | 16.59 |
| I/δ (I) | 24.5 (5.0) |
| Redundancy | 5.27 |
| Completeness (out shell) (%) | 99.82 (98.44) |
| R-work | 0.1691 (0.2558) |
| R-free | 0.1972 (0.2606) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhang, H.; Lv, P.; Li, Y.; Teng, M.; Liu, Y.; Wu, D. Structural Insights into Mouse H-FABP. Life 2022, 12, 1445. https://doi.org/10.3390/life12091445
Wang L, Zhang H, Lv P, Li Y, Teng M, Liu Y, Wu D. Structural Insights into Mouse H-FABP. Life. 2022; 12(9):1445. https://doi.org/10.3390/life12091445
Chicago/Turabian StyleWang, Lili, Haoran Zhang, Panjing Lv, Yan Li, Maikun Teng, Yahui Liu, and Donghai Wu. 2022. "Structural Insights into Mouse H-FABP" Life 12, no. 9: 1445. https://doi.org/10.3390/life12091445
APA StyleWang, L., Zhang, H., Lv, P., Li, Y., Teng, M., Liu, Y., & Wu, D. (2022). Structural Insights into Mouse H-FABP. Life, 12(9), 1445. https://doi.org/10.3390/life12091445