


Natural Ghee Enhances the Biochemical and Immunohistochemical Reproductive Performance of Female Rabbits

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Dietary Fat Sources
2.3. Experimental Design
2.4. Blood Sampling
2.5. Measuring Serum Biochemical Parameters
2.6. Ovarian Tissue Sampling
2.7. Statistical Analysis
3. Results
3.1. Fertility Hormonal Effects
3.2. Effects on Lipid Profile, Hepatic (ALT), and Renal (Creatinine)
3.3. Histopathological Examination
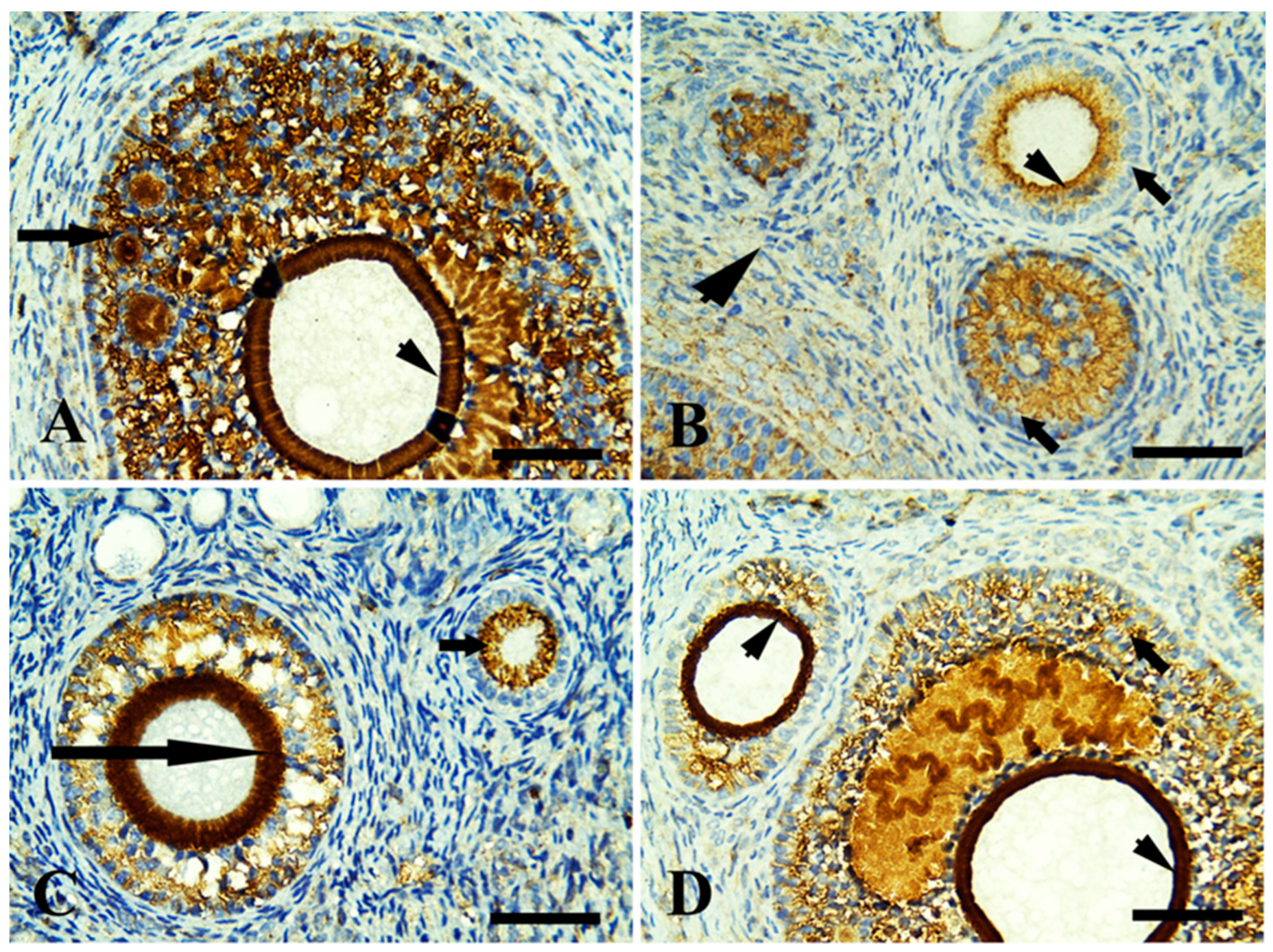
3.4. Immunohistochemical Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newton, I.S. Long-Chain Fatty Acids in Health and Nutrition; ACS Publications: Washington, DC, USA, 2001. [Google Scholar]
- Aluyor, E.; Ozigagu, C.; Oboh, O.; Aluyor, P. Chromatographic analysis of vegetable oils: A review. Sci. Res. Essays 2009, 4, 191–197. [Google Scholar]
- Covas, M.-I. Olive oil and the cardiovascular system. Pharmacol. Res. 2007, 55, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Tufarelli, V.; Laudadio, V.; Casalino, E. An extra-virgin olive oil rich in polyphenolic compounds has antioxidant effects in meat-type broiler chickens. Environ. Sci. Pollut. Res. 2016, 23, 6197–6204. [Google Scholar] [CrossRef]
- Iqbal, M.P. Trans fatty acids–A risk factor for cardiovascular disease. Pak. J. Med. Sci. 2014, 30, 194. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, Y.-Z.; Hu, C.-H. Radical-induced cis–trans isomerization of fatty acids: A theoretical study. J. Phys. Chem. A 2014, 118, 4554–4564. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, I.A.; Wanders, A.J.; Katan, M.B. Effect of animal and industrial trans fatty acids on HDL and LDL cholesterol levels in humans–a quantitative review. PLoS ONE 2010, 5, e9434. [Google Scholar] [CrossRef]
- Odegaard, A.O.; Pereira, M.A. Trans fatty acids, insulin resistance, and type 2 diabetes. Nutr. Rev. 2006, 64, 364–372. [Google Scholar] [CrossRef]
- Willerson, J.T. Systemic and local inflammation in patients with unstable atherosclerotic plaques. Prog. Cardiovasc. Dis. 2002, 44, 469–478. [Google Scholar] [CrossRef]
- Sserunjogi, M.L.; Abrahamsen, R.K.; Narvhus, J. A review paper: Current knowledge of ghee and related products. Int. Dairy J. 1998, 8, 677–688. [Google Scholar] [CrossRef]
- Sindhuja, S.; Prakruthi, M.; Manasa, R.; Shivananjappa, M. Health benefits of ghee (clarified butter)-A review from ayurvedic perspective. IP J. Nutr. Metab. Health Sci. 2020, 3, 64–72. [Google Scholar]
- Chaturvedi, P.; Moseki, P.; Mazunga, K. Effects of Used Sunflower Oil and Ghee (Clarified Butter) on Lipid Profile and Antioxidants in SD Rats. Eur. J. Biomed. Pharm. Sci. EJBPS 2016, 3, 59–64. [Google Scholar]
- Hassanzadeh-Taheri, M.; Jahani, F.; Hassanzadeh-Taheri, M.; Doostabadi, M.; Doostabadi, H.; Hosseini, M. The impacts of yoghurt butter oil on rat testicular morphology and sexual hormones in a 150-day study. Comp. Clin. Pathol. 2018, 27, 959–965. [Google Scholar] [CrossRef]
- Egba, S.; Udom, I.; Okonkwo, C. Comparative effect of oral administration of some dietary lipids on fertility hormones of female wistar albino rats. Glob. J. Biotech. Biochem. 2014, 9, 24–29. [Google Scholar]
- Encinias, H.; Lardy, G.; Encinias, A.; Bauer, M. High linoleic acid safflower seed supplementation for gestating ewes: Effects on ewe performance, lamb survival, and brown fat stores. J. Anim. Sci. 2004, 82, 3654–3661. [Google Scholar] [CrossRef]
- de Catalfo, G.E.H.; de Alaniz, M.J.; Marra, C.A. Influence of commercial dietary oils on lipid composition and testosterone production in interstitial cells isolated from rat testis. Lipids 2009, 44, 345. [Google Scholar] [CrossRef]
- Mumford, S.L.; Chavarro, J.E.; Zhang, C.; Perkins, N.J.; Sjaarda, L.A.; Pollack, A.Z.; Schliep, K.C.; Michels, K.A.; Zarek, S.M.; Plowden, T.C. Dietary fat intake and reproductive hormone concentrations and ovulation in regularly menstruating women. Am. J. Clin. Nutr. 2016, 103, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, A.; Espensen, P.L.; Hansen, B.F.; Ibsen, P. The influence of dietary olive oil and margarine on aortic cholesterol accumulation in cholesterol-fed rabbits maintained at similar plasma cholesterol level. Atherosclerosis 1992, 96, 159–170. [Google Scholar] [CrossRef]
- Fassati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Stone, P.; Ellis, S.; Colwell, J.A. Cholesterol determination in high-density lipoproteins separated by three different methods. Clin. Chem. 1977, 23, 882–884. [Google Scholar] [CrossRef]
- Tremblay, R.E.; Nagin, D.S.; Séguin, J.R.; Zoccolillo, M.; Zelazo, P.D.; Boivin, M.; Perusse, D.; Japel, C. Physical aggression during early childhood: Trajectories and predictors. Pediatrics 2004, 114, e43–e50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Voller, A.; Bartlett, A.; Bidwell, D.E. Enzyme immunoassays with special reference to ELISA techniques. J. Clin. Pathol. 1978, 31, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, H.M.; El-Sayed, Y.S.; Naser, S.M.; Eltahawy, A.S.; Onoda, A.; Umezawa, M. Efficacy of α-lipoic acid against cadmium toxicity on metal ion and oxidative imbalance, and expression of metallothionein and antioxidant genes in rabbit brain. Environ. Sci. Pollut. Res. 2017, 24, 24593–24601. [Google Scholar] [CrossRef] [PubMed]
- Draper, H.; Hadley, M. [43] Malondialdehyde determination as index of lipid Peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Nishikimi, M.; Appaji, N.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Philadelphia, PA, USA, 2008. [Google Scholar]
- Sharma, H.; Zhang, X.; Dwivedi, C. The effect of ghee (clarified butter) on serum lipid levels and microsomal lipid peroxidation. Ayu 2010, 31, 134. [Google Scholar] [CrossRef] [Green Version]
- Gross, D.A.; Silver, D.L. Cytosolic lipid droplets: From mechanisms of fat storage to disease. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 304–326. [Google Scholar] [CrossRef]
- Krahmer, N.; Farese, R.V., Jr.; Walther, T.C. Balancing the fat: Lipid droplets and human disease. EMBO Mol. Med. 2013, 5, 973–983. [Google Scholar] [CrossRef]
- Hamza-Reguig, S.; Daidj, N.B.B.; Louala, S.; Boualga, A.; Lamri-Senhadji, M. Effect of replacing sardine oil with margarine on dyslipidemia, dysglycemia and redox status of adipose tissue in high-fat diet-induced obesity in Wistar rats. Nutr. Food Sci. 2017, 47, 2–17. [Google Scholar] [CrossRef]
- Abd-Rabo, F.H.; Elshaghabee, F.M.; Sakr, S.S.; El-Arabi, N.I.; El-Maaty, S.A. Different dietary fats impact on biochemical and histological parameters and gene expression of lipogenesis-related genes in rats. Food Biosci. 2020, 34, 100540. [Google Scholar] [CrossRef]
- Longhi, R.; Almeida, R.F.; Machado, L.; Duarte, M.M.M.F.; Souza, D.G.; Machado, P.; de Assis, A.M.; Quincozes-Santos, A.; Souza, D.O. Effect of a trans fatty acid-enriched diet on biochemical and inflammatory parameters in Wistar rats. Eur. J. Nutr. 2017, 56, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Pischon, T.; Hankinson, S.E.; Rifai, N.; Joshipura, K.; Willett, W.C.; Rimm, E.B. Dietary intake of trans fatty acids and systemic inflammation in women. Am. J. Clin. Nutr. 2004, 79, 606–612. [Google Scholar] [CrossRef]
- Karabulut, I.; Turan, S. Some properties of margarines and shortenings marketed in Turkey. J. Food Compos. Anal. 2006, 19, 55–58. [Google Scholar] [CrossRef]
- Ohlsson, L. Dairy products and plasma cholesterol levels. Food Nutr. Res. 2010, 54, 5124. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.M.; Iqbal, S.; Rashid, M.A. Comparison of lipid lowering effect of extra virgin olive oil and atorvastatin in dyslipidaemia in type 2 diabetes mellitus. JAMC 2017, 29, 83–86. [Google Scholar]
- Massimo, C.; Lucio, T.; Jesus, M.; Giovanni, L.; Caramia, G.M. Extra virgin olive oil and oleic acid. Nutr. Clin. Y Diet. Hosp. 2009, 29, 12–24. [Google Scholar]
- Nour, S.; Eldin, N.; Abd Alla, A.; Abd Elhady, A. Health Impact of Traditional Egyptian Ghee. Alex. Sci. Exch. J. 2018, 39, 620–628. [Google Scholar]
- Abo-Ghanema, I.I.; Aboelnour, A.; Abd Elrehim, A.M.; Ghoneim, H.A. Effect of Different Dietary Fat Sources on Lipid Profile and Testosterone Level in Male Albino Rats. Damanhour J. Vet. Sci. 2022, 8, 24–30. [Google Scholar]
- Vega, G.L.; Toto, R.D.; Grundy, S.M. Metabolism of low density lipoproteins in nephrotic dyslipidemia: Comparison of hypercholesterolemia alone and combined hyperlipidemia. Kidney Int. 1995, 47, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Karandikar, Y.S.; Bansude, A.S.; Angadi, E.A. Comparison between the effect of cow ghee and butter on memory and lipid profile of wistar rats. J. Clin. Diagn. Res. JCDR 2016, 10, FF11. [Google Scholar] [CrossRef] [PubMed]
- Al-Othman, A.A. Growth and lipid metabolism responses in rats fed different dietary fat sources. Int. J. Food Sci. Nutr. 2000, 51, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Kumthekar, M.M.; Katyare, S.S. Altered kinetic attributes of Na (+)+ K (+)-ATPase activity in kidney, brain and erythrocyte membranes in alloxan-diabetic rats. Indian J. Exp. Biol. 1992, 30, 26–32. [Google Scholar] [PubMed]
- Dhibi, M.; Brahmi, F.; Mnari, A.; Houas, Z.; Chargui, I.; Bchir, L.; Gazzah, N.; Alsaif, M.A.; Hammami, M. The intake of high fat diet with different trans fatty acid levels differentially induces oxidative stress and non alcoholic fatty liver disease (NAFLD) in rats. Nutr. Metab. 2011, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siener, R.; Alteheld, B.; Terjung, B.; Junghans, B.; Bitterlich, N.; Stehle, P.; Metzner, C. Change in the fatty acid pattern of erythrocyte membrane phospholipids after oral supplementation of specific fatty acids in patients with gastrointestinal diseases. Eur. J. Clin. Nutr. 2010, 64, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Matuszewska, J.; Ziarniak, K.; Dudek, M.; Kołodziejski, P.; Pruszyńska-Oszmałek, E.; Śliwowska, J.H. Effects of short-term exposure to high-fat diet on histology of male and female gonads in rats. Acta Histochem. 2020, 122, 151558. [Google Scholar] [CrossRef]
- Mekircha, F.; Chebab, S.; Gabbianelli, R.; Leghouchi, E. The possible ameliorative effect of Olea europaea L. oil against deltamethrin-induced oxidative stress and alterations of serum concentrations of thyroid and reproductive hormones in adult female rats. Ecotoxicol. Environ. Saf. 2018, 161, 374–382. [Google Scholar] [CrossRef]
- Ochoa-Herrera, J.J.; Huertas, J.R.; Quiles, J.L.; Mataix, J. Dietary oils high in oleic acid, but with different non-glyceride contents, have different effects on lipid profiles and peroxidation in rabbit hepatic mitochondria. J. Nutr. Biochem. 2001, 12, 357–364. [Google Scholar] [CrossRef]
- Chimi, H.; Morel, I.; Lescoat, G.; Pasdeloup, N.; Cillard, P.; Cillard, J. Inhibition of iron toxicity in rat hepatocyte culture by natural phenolic compounds. Toxicol. Vitr. 1995, 9, 695–702. [Google Scholar] [CrossRef]
- Jensen, T.K.; Heitmann, B.L.; Jensen, M.B.; Halldorsson, T.I.; Andersson, A.-M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgård, C. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.V.; Sambaiah, K.; Lokesh, B.R. Hypocholesterolemic effect of anhydrous milk fat ghee is mediated by increasing the secretion of biliary lipids. J. Nutr. Biochem. 2000, 11, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Usta, B.; Yılmaz-Ersan, L. Antioxidant enzymes of milk and their biological effects. Ziraat Fakültesi Derg. Uludağ Üniversitesi 2013, 27, 123–130. [Google Scholar]
- Spiteller, G. The relation of lipid peroxidation processes with atherogenesis: A new theory on atherogenesis. Mol. Nutr. Food Res. 2005, 49, 999–1013. [Google Scholar] [CrossRef]
- Bowman, W.C.; Rand, M.J. Textbook of Pharmacology; Blackwell Scientific Publications: Oxford, UK, 1980. [Google Scholar]
- Hodgen, G. Neuroendocrinology of the normal menstrual cycle. J. Reprod. Med. 1989, 34, 68–75. [Google Scholar]
- Oi-Kano, Y.; Kawada, T.; Watanabe, T.; Koyama, F.; Watanabe, K.; Senbongi, R.; Iwai, K. Oleuropein supplementation increases urinary noradrenaline and testicular testosterone levels and decreases plasma corticosterone level in rats fed high-protein diet. J. Nutr. Biochem. 2013, 24, 887–893. [Google Scholar] [CrossRef]
- McCANN, S.M.; Mastronardi, C.; De Laurentiis, A.; Rettori, V. The nitric oxide theory of aging revisited. Ann. N. Y. Acad. Sci. 2005, 1057, 64–84. [Google Scholar] [CrossRef]
- Hertoghe, T. The “Multiple Hormone Deficienc” Theory of Aging: Is Human Senescence Caused Mainly by Multiple Hormone Deficiencies? Ann. N. Y. Acad. Sci. 2005, 1057, 448–465. [Google Scholar] [CrossRef]
- Chebab, S.; Mekircha, F.; Leghouchi, E. Potential protective effect of Pistacia lentiscus oil against chlorpyrifos-induced hormonal changes and oxidative damage in ovaries and thyroid of female rats. Biomed. Pharmacother. 2017, 96, 1310–1316. [Google Scholar] [CrossRef]
- Okuyama, H.; Ohara, N.; Tatematsu, K.; Fuma, S.; Nonogaki, T.; Yamada, K.; Ichikawa, Y.; Miyazawa, D.; Yasui, Y.; Honma, S. Testosterone-lowering activity of canola and hydrogenated soybean oil in the stroke-prone spontaneously hypertensive rat. J. Toxicol. Sci. 2010, 35, 743–747. [Google Scholar] [CrossRef]
- Li, M.; Cao, J.; Zhao, Y.; Wu, P.; Li, X.; Khodaei, F.; Han, Y.; Wang, J. Fluoride impairs ovary development by affecting oogenesis and inducing oxidative stress and apoptosis in female zebrafish (Danio rerio). Chemosphere 2020, 256, 127105. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, P.C.; Pawar, N.N.; Chandratre, G.A.; Telang, A.; Sharma, A. Fipronil induced oxidative stress in kidney and brain of mice: Protective effect of vitamin E and vitamin C. Pestic. Biochem. Physiol. 2015, 118, 10–18. [Google Scholar] [CrossRef]
- de Medeiros, H.C.D.; Constantin, J.; Ishii-Iwamoto, E.L.; Mingatto, F.E. Effect of fipronil on energy metabolism in the perfused rat liver. Toxicol. Lett. 2015, 236, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Ramos, E.; Ares, I.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.; Anadón, A.; Martínez, M. Fipronil sulfone induced higher cytotoxicity than fipronil in SH-SY5Y cells: Protection by antioxidants. Toxicol. Lett. 2016, 252, 42–49. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, Z.; Zhang, Y.; Shao, X.; Xu, X.; Cheng, J.; Li, Z. Fipronil induces apoptosis through caspase-dependent mitochondrial pathways in Drosophila S2 cells. Pestic. Biochem. Physiol. 2015, 119, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Taha, R.; Ghazy, E.W.; El-Sayed, Y.S. Synergistic ameliorative effects of sesame oil and alpha-lipoic acid against subacute diazinon toxicity in rats: Hematological, biochemical, and antioxidant studies. Can. J. Physiol. Pharmacol. 2015, 94, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M.; Coombes, J.S.; Bennett, N.; Johnson, D.W.; Gobe, G.C. Oxidative stress, anti-oxidant therapies and chronic kidney disease. Nephrology 2012, 17, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Miner, J.L.; Cederberg, C.A.; Nielsen, M.K.; Chen, X.; Baile, C.A. Conjugated linoleic acid (CLA), body fat, and apoptosis. Obes. Res. 2001, 9, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Alsenosy, A.-w.A.; El-Far, A.H.; Sadek, K.M.; Ibrahim, S.A.; Atta, M.S.; Sayed-Ahmed, A.; Al Jaouni, S.K.; Mousa, S.A. Graviola (Annona muricata) attenuates behavioural alterations and testicular oxidative stress induced by streptozotocin in diabetic rats. PLoS One 2019, 14, e0222410. [Google Scholar] [CrossRef]
- Sadek, K.M.; Lebda, M.A.; Abouzed, T.K. The possible neuroprotective effects of melatonin in aluminum chloride-induced neurotoxicity via antioxidant pathway and Nrf2 signaling apart from metal chelation. Environmental Science and Pollution Research 2019, 26, 9174–9183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.-J.; Hu, L.-L.; Ren, Y.-P.; Lu, X.; Luo, X.-Q.; Li, Y.-H.; Xu, Y.-N. Podophyllotoxin affects porcine oocyte maturation by inducing oxidative stress-mediated early apoptosis. Toxicon 2020, 176, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Moghissi, K. Gonadotropin releasing hormones. Clinical applications in gynecology. J. Reprod. Med. 1990, 35, 1097–1107. [Google Scholar] [PubMed]
- Castañeda-Gutiérrez, E.; Benefield, B.; De Veth, M.; Santos, N.; Gilbert, R.; Butler, W.; Bauman, D. Evaluation of the mechanism of action of conjugated linoleic acid isomers on reproduction in dairy cows. J. Dairy Sci. 2007, 90, 4253–4264. [Google Scholar] [CrossRef]
- Perks, C.; Peters, A.; Wathes, D. Follicular and luteal expression of insulin-like growth factors I and II and the type 1 IGF receptor in the bovine ovary. Reproduction 1999, 116, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.; Spicer, L.; Hamilton, T.; Keefer, B. Effects of insulin-like growth factor I and insulin on proliferation and on basal and luteinizing hormone-induced steroidogenesis of bovine thecal cells: Involvement of glucose and receptors for insulin-like growth factor I and luteinizing hormone. J. Anim. Sci. 1995, 73, 3719–3731. [Google Scholar] [CrossRef]
- Spicer, L.J.; Chamberlain, C.S.; Maciel, S.M. Influence of gonadotropins on insulin-and insulin-like growth factor-I (IGF-I)-induced steroid production by bovine granulosa cells. Domest. Anim. Endocrinol. 2002, 22, 237–254. [Google Scholar] [CrossRef]
- Spicer, L.; Alvarez, P.; Prado, T.; Morgan, G.; Hamilton, T. Effects of intraovarian infusion of insulin-like growth factor-I on ovarian follicular function in cattle☆. Domest. Anim. Endocrinol. 2000, 18, 265–278. [Google Scholar] [CrossRef]
- Upadhyay, N.; Kumar, A.; Goyal, A.; Lal, D. Complete liquification time test coupled with solvent fractionation technique to detect adulteration of foreign fats in ghee. Int. J. Dairy Technol. 2017, 70, 110–118. [Google Scholar] [CrossRef]
- Antony, B.; Sharma, S.; Mehta, B.M.; Ratnam, K.; Aparnathi, K.D. Study of Fourier transform near infrared (FT-NIR) spectra of ghee (anhydrous milk fat). Int. J. Dairy Technol. 2018, 71, 484–490. [Google Scholar] [CrossRef]
- Sharma, H.M. Free radicals and natural antioxidants in health and disease. J. Appl. Nutr. 2002, 52, 26–44. [Google Scholar]
- Mumtaz, S.; Ali, S.; Khan, R.; Shakir, H.A.; Tahir, H.M.; Mumtaz, S.; Andleeb, S. Therapeutic role of garlic and vitamins C and E against toxicity induced by lead on various organs. Environ. Sci. Pollut. Res. 2020, 27, 8953–8964. [Google Scholar] [CrossRef] [PubMed]
- Clelland, E.; Peng, C. Endocrine/paracrine control of zebrafish ovarian development. Mol. Cell. Endocrinol. 2009, 312, 42–52. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | g | Ingredient | g |
|---|---|---|---|
| Yellow corn | 7.5 | Di-calcium phosphate | 0.5 |
| Wheat bran | 24 | DI-methionine | 0.2 |
| Barley | 20 | Anti-aflatoxin + anti-coccidian | 0.5 |
| Clover hay | 22 | Vitamin and minerals premix | 0.30 |
| Soybean meal (44% CP) | 23.5 | NaCl | 0.35 |
| Limestone | 1.15 | total | 100 |
| Group | Body Weight (kg) | Oxidant and Antioxidant Markers | ||
|---|---|---|---|---|
| MDA (nmol/mg Protein) | Catalase (U/mg Protein) | SOD (U/mg Protein) | ||
| Control (NC) | 3.9 ± 1.4 b | 101.3 ± 0.87 b | 31.7 ± 2.1 b | 14.7 ± 2 b |
| Margarine | 4.57 ±1.2 a | 145.8 ± 3.2 a | 18.3 ± 2.1 c | 8.9 ± 1.6 c |
| Ghee | 3.53 ± 1.9 b | 109 ± 1.8 b | 45.1 ± 1.8 a | 18.2 ± 1.2 a |
| Olive Oil | 3.75 ± 1.7 b | 92.9 ± 3.4 c | 46.5 ± 3.2 a | 21.3 ± 1.55 a |
| Group | Fertility Hormones | |||||
|---|---|---|---|---|---|---|
| FSH | Estradiol (E2) | LH | PROGES | PRL. | Testosterone | |
| Control | 6.6 ± 0.4 b | 77.1 ± 3.7 b | 4.8 ± 0.4 a | 1.84 ± 0.15 a | 16 ± 1.6 b | 1.38 ± 0.03 a |
| Margarine | 5.1 ± 0.2 c | 68.5 ± 1.5 c | 4.6 ± 0.12 a | 1.78 ± 0.09 a | 31 ± 2.5 a | 1.32 ± 0.08 a |
| Ghee | 9.3 ± 0.6 a | 88.4 ± 1.6 a | 5.09 ± 0.88 a | 2.1 ± 0.22 a | 14.5 ± 0.7 b | 1.41 ± 0.04 a |
| Olive Oil | 8.7 ± 1.4 a | 82 ± 4.5 a | 4.9 ± 0.33 a | 1.9 ± 0.05 a | 13.7 ± 1.2 b | 1.49 ± 0.01 a |
| Group | TAG | CHOL. | HDL | LDL | VLDL | ALT | Creatinine. |
|---|---|---|---|---|---|---|---|
| Control (NC) | 201.3 ± 2.9 a | 143.3 ± 9.3 b | 37 ± 2.1 b | 66 ± 3.5 b | 40.26 ± 0.6 a | 43.7 ± 4.2 b | 0.89 ± 0.05 b |
| Margarine | 204.8 ± 6.1 a | 173.2 ± 3.8 a | 26.7 ± 1.7 c | 105.5 ± 4.7 a | 41 ± 1.2 a | 55.8 ± 1.3 a | 1.3 ± 0.01 a |
| Ghee | 171.5 ± 5.8 b | 146.4 ± 3.1 b | 35.5 ± 2.2 b | 76.6 ± 4.1 b | 34.3 ± 1.2 b | 46.6 ± 1.8 b | 1.02 ± 0.04 b |
| Olive Oil | 148.6 ± 5.7 b | 119.3 ± 2.7 c | 48.5 ± 1.5 a | 41.1 ± 3.1 c | 29.7 ± 1.1 b | 44 ± 2.2 b | 0.92 ± 0.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Gharrawy, H.T.; Sadek, K.M.; Mahmoud, S.F.; Abd Elrehim, A.M.; Shukry, M.; Ghamry, H.I.; Ibrahim, S.F.; Fericean, L.; Abdo, M.; Zeweil, M.M. Natural Ghee Enhances the Biochemical and Immunohistochemical Reproductive Performance of Female Rabbits. Life 2023, 13, 80. https://doi.org/10.3390/life13010080
El-Gharrawy HT, Sadek KM, Mahmoud SF, Abd Elrehim AM, Shukry M, Ghamry HI, Ibrahim SF, Fericean L, Abdo M, Zeweil MM. Natural Ghee Enhances the Biochemical and Immunohistochemical Reproductive Performance of Female Rabbits. Life. 2023; 13(1):80. https://doi.org/10.3390/life13010080
Chicago/Turabian StyleEl-Gharrawy, Hassan T., Kadry M. Sadek, Sahar F. Mahmoud, Attaa. M. Abd Elrehim, Mustafa Shukry, Heba I. Ghamry, Samah F. Ibrahim, Liana Fericean, Mohamed Abdo, and Mohamed M. Zeweil. 2023. "Natural Ghee Enhances the Biochemical and Immunohistochemical Reproductive Performance of Female Rabbits" Life 13, no. 1: 80. https://doi.org/10.3390/life13010080
APA StyleEl-Gharrawy, H. T., Sadek, K. M., Mahmoud, S. F., Abd Elrehim, A. M., Shukry, M., Ghamry, H. I., Ibrahim, S. F., Fericean, L., Abdo, M., & Zeweil, M. M. (2023). Natural Ghee Enhances the Biochemical and Immunohistochemical Reproductive Performance of Female Rabbits. Life, 13(1), 80. https://doi.org/10.3390/life13010080