
Exploring the Impact of Controlled Ovarian Stimulation and Non-Invasive Oocyte Assessment in ART Treatments


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Search Methods
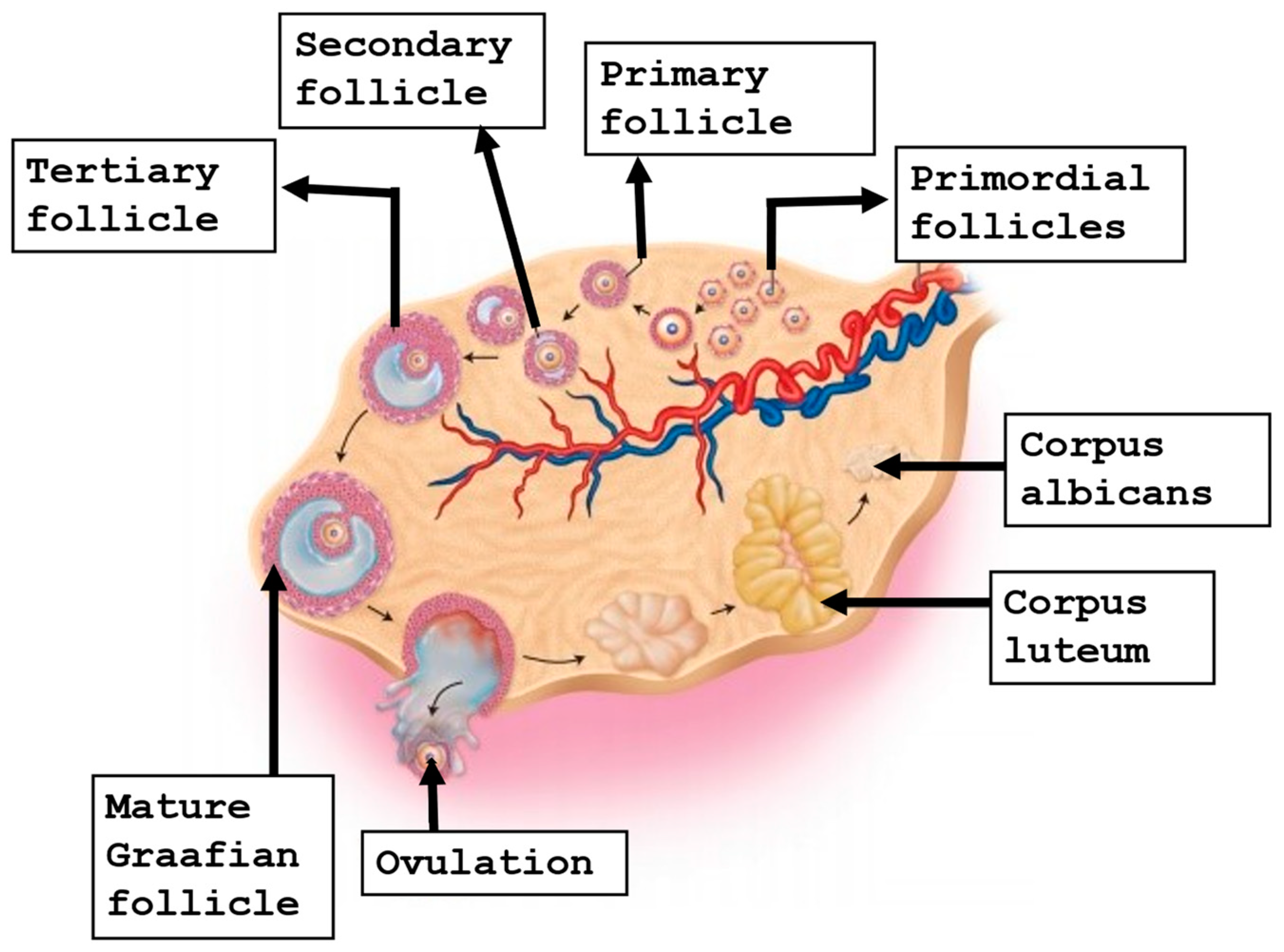
3. Folliculogenesis
4. Oocyte Competence in ART Cycles
5. The Relevance of OS in ART
5.1. Mild or Moderate Ovarian Stimulation
5.2. Full Stimulation and Risk of OHSS
5.3. Long-Acting Recombinant FSH
5.4. OS with Adjuvant Treatment
5.5. Double Stimulation (DuoStim): Is It Risky?
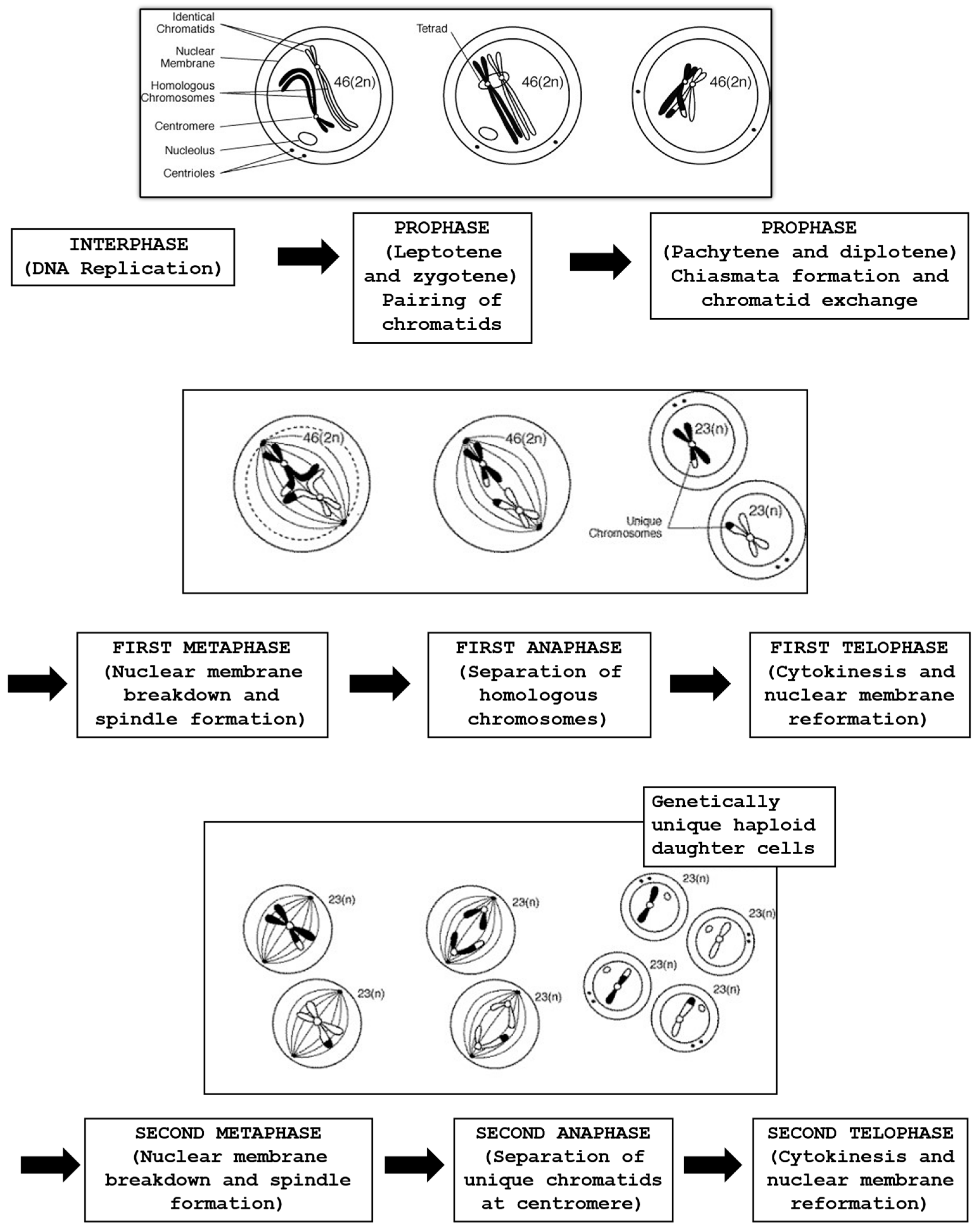
6. Oocyte Nuclear Maturation
6.1. From Birth to Puberty: Oocyte Chromatin Segregation and Resumption of Meiosis
6.2. Cytoplasmic Maturation
6.3. Polar Body Appearance
7. The Influence of Noninvasive Evaluation of Oocyte Quality
8. Further Concerns and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Geyter, C.; Wyns, C.; Calhaz-Jorge, C.; de Mouzon, J.; Ferraretti, A.P.; Kupka, M.; Andersen, A.N.; Nygren, K.G.; Goossens, V. 20 years of the European IVF-monitoring Consortium registry: What have we learned? A comparison with registries from two other regions. Hum. Reprod. 2020, 35, 2832–2849. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Mtango, N.R.; Potireddy, S.; Latham, K.E. Oocyte quality and maternal control of development. Int. Rev. Cell Mol. Biol. 2008, 268, 223–290. [Google Scholar] [CrossRef] [PubMed]
- Humaidan, P.; Alviggi, C.; Fischer, R.; Esteves, S.C. The novel POSEIDON stratification of ‘Low prognosis patients in Assisted Reproductive Technology’ and its proposed marker of successful outcome. F1000Research 2016, 5, 2911. [Google Scholar] [CrossRef]
- Mahutte, N.G.; Arici, A. Failed fertilization: Is it predictable? Curr. Opin. Obs. Gynecol. 2003, 15, 211–218. [Google Scholar] [CrossRef]
- Swain, J.E.; Pool, T.B. ART failure: Oocyte contributions to unsuccessful fertilization. Hum. Reprod. Update 2008, 14, 431–446. [Google Scholar] [CrossRef]
- Marteil, G.; Richard-Parpaillon, L.; Kubiak, J.Z. Role of oocyte quality in meiotic maturation and embryonic development. Reprod. Biol. 2009, 9, 203–224. [Google Scholar] [CrossRef]
- Nagy, Z.P.; Jones-Colon, S.; Roos, P.; Botros, L.; Greco, E.; Dasig, J.; Behr, B. Metabolomic assessment of oocyte viability. Reprod. Biomed. Online 2009, 18, 219–225. [Google Scholar] [CrossRef]
- Telfer, E.E.; Grosbois, J.; Odey, Y.L.; Rosario, R.; Anderson, R.A. Making a good egg: Human oocyte health, aging, and in vitro development. Physiol. Rev. 2023, 103, 2623–2677. [Google Scholar] [CrossRef]
- Combelles, C.M.; Racowsky, C. Assessment and Optimization of Oocyte Quality during Assisted Reproductive Technology Treatment. Semin. Reprod. Med. 2005, 23, 277–284. [Google Scholar] [CrossRef]
- Gondos, B.; Bhiraleus, P.; Hobel, C.J. Ultrastructural observations on germ cells in human fetal ovaries. Am. J. Obstet. Gynecol. 1971, 110, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.B.; Myers, M.; Anderson, R.A. The dynamics of the primordial follicle reserve. Reproduction 2013, 146, R205–R215. [Google Scholar] [CrossRef]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A.; Chainy, G.B. Morphometric studies of small follicles in ovaries of women at different ages. Reproduction 1987, 81, 433–442. [Google Scholar] [CrossRef] [PubMed]
- McGee, E.A.; Hsueh, A.J. Initial and Cyclic Recruitment of Ovarian Follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef]
- Albertini, D.F.; Combelles, C.M.; Benecchi, E.; Carabatsos, M.J. Cellular basis for paracrine regulation of ovarian follicle de-velopment. Reproduction 2001, 121, 647–653. [Google Scholar] [CrossRef]
- Soyal, S.M.; Amleh, A.; Dean, J. FIGalpha, a germ cell-specific transcription factor required for ovarian follicle formation. Development 2000, 127, 4645–4654. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Irving-Rodgers, H.F.; van Wezel, I.L.; Krupa, M.; Lavranos, T.C. Dynamics of the membrana granulosa during expansion of the ovarian follicular antrum. Mol. Cell. Endocrinol. 2001, 171, 41–48. [Google Scholar] [CrossRef]
- Diaz, F.J.; Wigglesworth, K.; Eppig, J.J. Oocytes determine cumulus cell lineage in mouse ovarian follicles. J. Cell Sci. 2007, 120, 1330–1340. [Google Scholar] [CrossRef]
- Eppig, J.J. Intercommunication between mammalian oocytes and companion somatic cells. BioEssays 1991, 13, 569–574. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Lalioti, M.D.; Aydiner, F.; Sasson, I.; Ilbay, O.; Sakkas, D.; Lowther, K.M.; Mehlmann, L.M.; Seli, E. Embryonic poly(A)-binding protein (EPAB) is required for oocyte maturation and female fertility in mice. Biochem. J. 2012, 446, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Ezoe, K.; Miki, T.; Akaike, H.; Shimazaki, K.; Takahashi, T.; Tanimura, Y.; Amagai, A.; Sawado, A.; Mogi, M.; Kaneko, S.; et al. Maternal age affects pronuclear and chromatin dynamics, morula compaction and cell polarity, and blastulation of human embryos. Hum. Reprod. 2023, 38, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Minasi, M.G.; Anagnostopoulou, C.; Boitrelle, F.; Vogiatzi, P.; Sallam, H.; Saleh, R.; Colpi, G.; Agarwal, A. Oocytes evaluation and in-vitro fertilization/intra cytoplasmic sperm injection outcomes. Panminerva Med. 2023, 65, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Rienzi, L.; Vajta, G.; Ubaldi, F. Predictive value of oocyte morphology in human IVF: A systematic review of the literature. Hum. Reprod. Update 2010, 17, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Tiitinen, A. Single embryo transfer: Why and how to identify the embryo with the best developmental potential. Best Pr. Res. Clin. Endocrinol. Metab. 2019, 33, 77–88. [Google Scholar] [CrossRef]
- Ciepiela, P.; Baczkowski, T.; Drozd, A.; Kazienko, A.; Stachowska, E.; Kurzawa, R. Arachidonic and linoleic acid derivatives impact oocyte ICSI fertilization--a prospective analysis of follicular fluid and a matched oocyte in a ‘one follicle--one retrieved oocyte--one resulting embryo’ investigational setting. PLoS ONE 2015, 10, e0119087. [Google Scholar] [CrossRef]
- Griesinger, G.; Diedrich, K.; Devroey, P.; Kolibianakis, E.M. GnRH agonist for triggering final oocyte maturation in the GnRH antagonist ovarian hyperstimulation protocol: A systematic review and meta-analysis. Hum. Reprod. Update 2006, 12, 159–168. [Google Scholar] [CrossRef]
- Griesinger, G.; Vverweij, P.J.; Gates, D.; Devroey, P.; Gordon, K.; Stegmann, B.J.; Tarlatzis, B.C. Prediction of ovarian hyper-stimulation syndrome in patients treated with corifollitropin alfa or rFSH in a GnrH antagonist protocol. PLoS ONE 2016, 11, e0149615. [Google Scholar] [CrossRef]
- Liebermann, J.; Tucker, M.J. Comparison of vitrification and conventional cryopreservation of day 5 and day 6 blastocysts during clinical application. Fertil. Steril. 2006, 86, 20–26. [Google Scholar] [CrossRef]
- Dosouto, C.; Haahr, T.; Humaidan, P. Gonadotropin-releasing hormone agonist (GnRHa) trigger—State of the art. Reprod. Biol. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Ferraretti, A.P.; Gianaroli, L.; Magli, C.; Fortini, D.; Selman, H.A.; Feliciani, E. Elective cryopreservation of all pronucleate embryos in women at risk of ovarian hyperstimulation syndrome: Efficiency and safety. Hum. Reprod. 1999, 14, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Roque, M.; Lattes, K.; Serra, S.; Solà, I.; Geber, S.; Carreras, R.; Checa, M.A. Fresh embryo transfer versus frozen embryo transfer in in vitro fertilization cycles: A systematic review and meta-analysis. Fertil. Steril. 2013, 99, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Roque, M. Freeze-all policy: Is it time for that? J. Assist. Reprod. Genet. 2015, 32, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Roque, M.; Haahr, T.; Geber, S.; Esteves, S.C.; Humaidan, P. Fresh versus elective frozen embryo transfer in IVF/ICSI cycles: A systematic review and meta-analysis of reproductive outcomes. Hum. Reprod. Update 2019, 25, 2–14. [Google Scholar] [CrossRef]
- Gullo, G.; Basile, G.; Cucinella, G.; Greco, M.E.; Perino, A.; Chiantera, V.; Marinelli, S. Affiliations expand Fresh vs. frozen embryo transfer in assisted reproductive techniques: A single center retrospective cohort study and ethical-legal im-plications. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 6809–6823. [Google Scholar]
- Gullo, G.; Perino, A.; Cucinella, G. Open vs. closed vitrification system: Which one is safer? Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1065–1067. [Google Scholar] [CrossRef]
- Sciorio, R.; Thong, K.; Pickering, S.J. Single blastocyst transfer (SET) and pregnancy outcome of day 5 and day 6 human blastocysts vitrified using a closed device. Cryobiology 2018, 84, 40–45. [Google Scholar] [CrossRef]
- Sciorio, R.; Thong, K.J.; Pickering, S.J. Increased pregnancy outcome after day 5 versus day 6 transfers of human vitri-fied-warmed blastocysts. Zygote 2019, 27, 279–284. [Google Scholar] [CrossRef]
- Burgio, S.; Polizzi, C.; Buzzaccarini, G.; Laganà, A.S.; Gullo, G.; Perricone, G.; Perino, A.; Cucinella, G.; Alesi, M. Psychological variables in medically assisted reproduction: A systematic review. Menopausal Rev. 2022, 21, 47–63. [Google Scholar] [CrossRef]
- Santos, M.A.; Kuijk, E.W.; Macklon, N.S. The impact of ovarian stimulation for IVF on the developing embryo. Reproduction 2010, 139, 23–34. [Google Scholar] [CrossRef]
- Casarini, L.; Paradiso, E.; Lazzaretti, C.; D’alessandro, S.; Roy, N.; Mascolo, E.; Zaręba, K.; García-Gasca, A.; Simoni, M. Regulation of antral follicular growth by an interplay between gonadotropins and their receptors. J. Assist. Reprod. Genet. 2022, 39, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, J.V.; Kristensen, S.G.; Nielsen, M.E.; Humaidan, P.; Canto, M.D.; Fadini, R.; Schmidt, K.T.; Ernst, E.; Andersen, C.Y. LH-Receptor Gene Expression in Human Granulosa and Cumulus Cells from Antral and Preovulatory Follicles. J. Clin. Endocrinol. Metab. 2012, 97, E1524–E1531. [Google Scholar] [CrossRef] [PubMed]
- Bosch, E.; Alviggi, C.; Lispi, M.; Conforti, A.; Hanyaloglu, A.C.; Chuderland, D.; Simoni, M.; Raine-Fenning, N.; Crépieux, P.; Kol, S.; et al. Reduced FSH and LH action: Implica-tions for medically assisted reproduction. Hum. Reprod. 2021, 36, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Pakarainen, T.; Zhang, F.-P.; Nurmi, L.; Poutanen, M.; Huhtaniemi, I. Knockout of luteinizing hormone receptor abolishes the effects of follicle-stimulating hormone on preovulatory maturation and ovulation of mouse graafian follicles. Mol. Endocrinol. 2005, 19, 2591–25602. [Google Scholar] [CrossRef]
- Galway, A.B.; Lapolt, P.S.; Tsafriri, A.; Dargan, C.M.; Boime, I.; Hsueh, A.J. Recombinant follicle-stimulating hormone induces ovulation and tissue plasminogen activator expression in hypophysectomized rats. Endocrinology 1990, 127, 3023–3028. [Google Scholar] [CrossRef] [PubMed]
- Casarini, L.; Simoni, M. Recent advances in understanding gonadotropin signaling. Fac. Rev. 2021, 10, 41. [Google Scholar] [CrossRef]
- Filicori, M.; Cognigni, G.E.; Samara, A.; Melappioni, S.; Perri, T.; Cantelli, B.; Parmegiani, L.; Pelusi, G.; DeAloysio, D. The use of LH activity to drive folliculo-genesis: Exploring uncharted territories in ovulation induction. Hum. Reprod. Update 2002, 8, 543–557. [Google Scholar] [CrossRef]
- Khalili, M.A.; Shahedi, S.A.N.; Macchiarelli, G. Contribution of human oocyte architecture to success of in vitro maturation technology. Iran. J. Reprod. Med. 2013, 11, 1–10. [Google Scholar]
- Al Jord, A.; Letort, G.; Chanet, S.; Tsai, F.-C.; Antoniewski, C.; Eichmuller, A.; Da Silva, C.; Huynh, J.-R.; Gov, N.S.; Voituriez, R.; et al. Cytoplasmic forces functionally reorganize nuclear condensates in oocytes. Nat. Commun. 2022, 13, 5070. [Google Scholar] [CrossRef]
- Vaiarelli, A.; Cimadomo, D.; Scarafia, C.; Innocenti, F.; Amendola, M.G.; Fabozzi, G.; Casarini, L.; Conforti, A.; Alviggi, C.; Gennarelli, G.; et al. Metaphase-II oocyte competence is unlinked to the gonadotrophins used for ovarian stimulation: A matched case–control study in women of advanced maternal age. J. Assist. Reprod. Genet. 2023, 40, 169–177. [Google Scholar] [CrossRef]
- Santi, D.; Casarini, L.; Alviggi, C.; Simoni, M. Response: Commentary: Efficacy of Follicle-Stimulating Hormone (FSH) Alone, FSH + Luteinizing Hormone, Human Menopausal Gonadotropin or FSH + Human Chorionic Gonadotropin on Assisted Re-productive Technology Outcomes in the “Personalized” Medicine Era: A Meta-analysis. Front. Endocrinol. 2018, 9, 113. [Google Scholar]
- Conforti, A.; Esteves, S.C.; Humaidan, P.; Longobardi, S.; D’ Hooghe, T.; Orvieto, R.; Vaiarelli, A.; Cimadomo, D.; Rienzi, L.; Ubaldi, F.M.; et al. Recombinant human luteinizing hormone co-treatment in ovarian stimulation for assisted reproductive technology in women of advanced reproductive age: A systematic review and meta-analysis of randomized controlled trials. Reprod. Biol. Endocrinol. 2021, 19, 91. [Google Scholar] [CrossRef] [PubMed]
- Humaidan, P.; Chin, W.; Rogoff, D.; D’Hooghe, T.; Longobardi, S.; Hubbard, J.; Schertz, J.; ESPART Study Investigators. Efficacy and safety of follitropin al-fa/lutropin alfa in ART: A randomized controlled trial in poor ovarian responders. Hum. Reprod. 2017, 32, 544–555. [Google Scholar] [CrossRef]
- Alviggi, C.; Conforti, A. Mild/moderate versus full stimulation. Fertil. Steril. 2022, 117, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Banker, M.; Garcia-Velasco, J.A. Revisiting ovarian hyper stimulation syndrome: Towards OHSS free clinic. J. Hum. Reprod. Sci. 2015, 8, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Devroey, P.; Adriaensen, P. OHSS Free Clinic. Facts Views Vis. Obgyn. 2011, 3, 43–45. [Google Scholar] [PubMed]
- The Corifollitropin Alfa Dose-finding Study Group A randomized dose-response trial of a single injection of corifollitropin alfa to sustain multifollicular growth during controlled ovarian stimulation. Hum. Reprod. 2008, 23, 2484–2492. [CrossRef]
- Pouwer, A.W.; Farquhar, C.; Kremer, J.A. Long-acting FSH versus daily FSH for women undergoing assisted reproduction. Cochrane Database Syst. Rev. 2015, 7, CD009577. [Google Scholar] [CrossRef]
- Devroey, P.; Boostanfar, R.; Koper, N.; Mannaerts, B.; Ijzerman-Boon, P.; Fauser, B.; on behalf of the ENGAGE Investigators. A double-blind, non-inferiority RCT comparing corifollitropin alfa and recombinant FSH during the first seven days of ovarian stimulation using a GnRH antagonist protocol. Hum. Reprod. 2009, 24, 3063–3072. [Google Scholar] [CrossRef]
- Corifollitropin Alfa Ensure Study Group. Corifollitropin alfa for ovarian stimulation in IVF: A randomized trial in low-er-body-weight women. Reprod. Biomed. Online 2010, 21, 66–76. [Google Scholar] [CrossRef]
- Norman, R.J.; Zegers-Hochschild, F.; Salle, B.S.; Elbers, J.; Heijnen, E.; Marintcheva-Petrova, M.; Mannaerts, B.; Trust Investigators. Repeated ovarian stimulation with corifollitropin alfa in patients in a GnRH anatagonist protocol: No concern for immunogenicity. Hum. Reprod. 2011, 26, 2200–2208. [Google Scholar] [CrossRef] [PubMed]
- Boostanfar, R.; Mannaerts, B.; Pang, S.; Fernandez-Sanchez, M.; Witjes, H.; Devroey, P. A comparison of live birth rates and cumulative ongoing pregnancy rates between Europe and North America after ovarian stimulation with corifollitropin alfa or recombinant follicle-stimulating hormone. Fertil. Steril. 2012, 97, 1351–1358. [Google Scholar] [CrossRef]
- Griesinger, G.; Boostanfar, R.; Gordon, K.; Gates, D.; Sisk, C.M.; Stegmann, B.J. Corifollitropin alfa versus recombinant follicle-stimulating hormone: An individual patient data me-ta-analysis. Reprod. Biomed. Online 2016, 33, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Fusi, F.M.; Zanga, L.; Arnoldi, M.; Melis, S.; Cappato, M.; Candeloro, I.; Di Pasqua, A. Corifollitropin alfa for poor responders patients, a prospective randomized study. Reprod. Biol. Endocrinol. 2020, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-Y.; Li, Y.-C.; Tian, X.-Y.; Zhou, Y.; Guo, K.-P.; Fan, H.-Y.; Liang, X.-W.; Ou, X.-H.; Sha, Q.-Q. A long-acting recombinant FSH supports high-quality mouse follicle development and oocyte maturation in vitro by coordinating somatic and germ cell transcriptomes. Mol. Hum. Reprod. 2023, 29, gaad013. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Shu, J.; Guo, J.; Chang, H.-M.; Leung, P.C.K.; Sheng, J.-Z.; Huang, H. Adjuvant treatment strategies in ovarian stimulation for poor responders undergoing IVF: A systematic review and network meta-analysis. Hum. Reprod. Update 2020, 26, 247–263. [Google Scholar] [CrossRef]
- The ESHRE Guideline Group. ESHRE Guideline: Ovarian stimulation for IVF/ICSI. Hum. Reprod. Open 2020, 2, hoaa009. [Google Scholar]
- Hohmann, F.P.; Macklon, N.S.; Fauser, B.C.J.M. A Randomized Comparison of Two Ovarian Stimulation Protocols with Gonadotropin-Releasing Hormone (GnRH) Antagonist Cotreatment for in Vitro Fertilization Commencing Recombinant Follicle-Stimulating Hormone on Cycle Day 2 or 5 with the Standard Long GnRH Agonist Protocol. J. Clin. Endocrinol. Metab. 2003, 88, 166–173. [Google Scholar] [CrossRef]
- Baart, E.B.; Martini, E.; Eijkemans, M.J.; Van Opstal, D.; Beckers, N.G.; Verhoeff, A.; Macklon, N.S.; Fauser, B.C. Milder ovarian stimulation for in-vitro fertilization reduces aneuploidy in the human preimplantation embryo: A randomized controlled trial. Hum. Reprod. 2007, 22, 980–988. [Google Scholar] [CrossRef]
- Luo, Y.Y.; Zeng, X.; Zhu, L.; Li, C.; Xie, J.; Dong, Q.; Sun, Q.-Y.; Huang, G.-H.; Li, J.-Y. Growth hormone reduces aneuploidy and improves oocytes quality by JAK2-MAPK3/1 pathway in aged mice. J. Transl. Med. 2023, 21, 426. [Google Scholar] [CrossRef]
- Zhou, P.; Baumgarten, S.C.; Wu, Y.; Bennett, J.; Winston, N.; Hirshfeld-Cytron, J.; Stocco, C. IGF-I Signaling is Essential for FSH Stimulation of AKT and Steroidogenic Genes in Granulosa Cells. Mol. Endocrinol. 2013, 27, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Bachelot, A.; Monget, P.; Imbert-Bolloré, P.; Coshigano, K.; Kopchick, J.J.; Kelly, P.A.; Binart, N. Growth Hormone is Required for Ovarian Follicular Growth. Endocrinology 2002, 143, 4104–4112. [Google Scholar] [CrossRef] [PubMed]
- Tocci, A. Why double ovarian stimulation in an in vitro fertilization cycle is potentially unsafe. Hum. Reprod. 2021, 37, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Casarini, L.; Vaiarelli, A.; Cimadomo, D.; Santi, D.; Simoni, M.; Garcìa-Velasco, J.A.; Alviggi, C.; La Marca, A.; Rienzi, L.; Ubaldi, F.M. Neither rationale nor scientific evidence exist to support that double stimulation is potentially unsafe. Hum. Reprod. 2022, 37, 1949–1950. [Google Scholar] [CrossRef] [PubMed]
- Rienzi, L.; Ubaldi, F.; Martinez, F.; Iacobelli, M.; Minasi, M.; Ferrero, S.; Tesarik, J.; Greco, E. Relationship between meiotic spindle location with regard to the polar body position and oocyte developmental potential after ICSI. Hum. Reprod. 2003, 18, 1289–1293. [Google Scholar] [CrossRef]
- Montag, M.; Schimming, T.; Koster, M.; Zhou, C.; Dorn, C.; Rosing, B.; van der Ven, H.; Ven der Ven, K. Oocyte zona bire-fringence intensity is associated with embryonic implantation potential in ICSI cycles. Reprod. Biomed. Online 2008, 16, 239–244. [Google Scholar] [CrossRef]
- Shen, Y.; Stalf, T.; Mehnert, C.; De Santis, L.; Cino, I.; Tinneberg, H.-R.; Eichenlaub-Ritter, U. Light retardance by human oocyte spindle is positively related to pronuclear score after ICSI. Reprod. Biomed. Online 2006, 12, 737–751. [Google Scholar] [CrossRef]
- Rienzi, L.; Ubaldi, F.; Iacobelli, M.; Minasi, M.G.; Romano, S.; Greco, E. Meiotic spindle visualization in living human oocytes. Reprod. Biomed. Online 2005, 10, 192–198. [Google Scholar] [CrossRef]
- Chamayou, S.; Ragolia, C.; Alecci, C.; Storaci, G.; Maglia, E.; Russo, E.; Guglielmino, A. Meiotic spindle presence and oocyte morphology do not predict clinical ICSI outcomes: A study of 967 transferred embryos. Reprod. Biomed. Online 2006, 13, 661–667. [Google Scholar] [CrossRef]
- Taylor, T.H.; Chang, C.-C.; Elliott, T.; Colturato, L.F.; Kort, H.I.; Nagy, Z.P. Effect of denuding on polar body position in in-vitro matured oocytes. Reprod. Biomed. Online 2008, 17, 515–519. [Google Scholar] [CrossRef]
- Rienzi, L.; Ubaldi, F. Oocyte Retrieval and Selection. In Textbook of Assisted Reproductive Technologies: Laboratory and Clinical Perspectives; Gardner, D., Ed.; Informa Healthcare: London, UK, 2009; pp. 5–101. [Google Scholar]
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef] [PubMed]
- Gorbsky, G.J. The spindle checkpoint and chromosome segregation in meiosis. FEBS J. 2015, 282, 2471–2487. [Google Scholar] [CrossRef] [PubMed]
- Canto, M.D.; Guglielmo, M.C.; Renzini, M.M.; Fadini, R.; Moutier, C.; Merola, M.; De Ponti, E.; Coticchio, G. Dysmorphic patterns are associated with cytoskeletal alterations in human oocytes. Hum. Reprod. 2017, 32, 1–8. [Google Scholar] [CrossRef]
- Montag, M.; van der Ven, H. Symposium: Innovative techniques in human embryo viability assessment. Oocyte assessment and embryo viability prediction: Birefringence imaging. Reprod. Biomed. Online 2008, 17, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-H.; Meng, L.; Hackett, R.J.; Oldenbourg, R.; Keefe, D.L. Rigorous thermal control during intracytoplasmic sperm injection stabilizes the meiotic spindle and improves fertilization and pregnancy rates. Fertil. Steril. 2002, 77, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; FitzHarris, G. Recent Insights into Spindle Function in Mammalian Oocytes and Early Embryos. Biol. Reprod. 2013, 89, 71. [Google Scholar] [CrossRef]
- Capalbo, A.; Bono, S.; Spizzichino, L.; Biricik, A.; Baldi, M.; Colamaria, S.; Ubaldi, F.M.; Rienzi, L.; Fiorentino, F. Sequential comprehensive chromosome analysis on polar bodies, blastomeres and trophoblast: Insights into female meiotic errors and chromosomal segregation in the preimplantation window of embryo development. Hum. Reprod. 2012, 28, 509–518. [Google Scholar] [CrossRef]
- Gonzalez, X.V.; Odia, R.; Naja, R.; Serhal, P.; Saab, W.; Seshadri, S.; Ben-Nagi, J. Euploid blastocysts implant irrespective of their morphology after NGS-(PGT-A) testing in advanced maternal age patients. J. Assist. Reprod. Genet. 2019, 36, 1623–1629. [Google Scholar] [CrossRef]
- Gullo, G.; Scaglione, M.; Buzzaccarini, G.; Laganà, A.S.; Basile, G.; Chiantera, V.; Cucinella, G.; Zaami, S. Cell-Free Fetal DNA and Non-Invasive Prenatal Diagnosis of Chromosomopathies and Pediatric Monogenic Diseases: A Critical Appraisal and Medicolegal Remarks. J. Pers. Med. 2022, 13, 1. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Henry, G. Oocyte dysmorphism and aneuploidy in meiotically mature human oocytes after ovarian stimula-tion. Hum. Reprod. 1992, 7, 379–390. [Google Scholar] [CrossRef]
- Shoubridge, E.A.; Wai, T. Mitochondrial DNA and the Mammalian Oocyte. Curr. Top. Dev. Biol. 2007, 77, 87–111. [Google Scholar] [CrossRef] [PubMed]
- Wai, T.; Ao, A.; Zhang, X.; Cyr, D.; Dufort, D.; Shoubridge, E.A. The Role of Mitochondrial DNA Copy Number in Mammalian Fertility. Biol. Reprod. 2010, 83, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub-Ritter, U.; Schmiady, H.; Kentenich, H.; Soewarto, D. Recurrent failure in polar body formation and premature chromosome condensation in oocytes from a human patient: Indicators of asynchrony in nuclear and cytoplasmic maturation. Hum. Reprod. 1995, 10, 2343–2349. [Google Scholar] [CrossRef]
- Ebner, T.; Moser, M.; Yaman, C.; Feichtinger, O.; Hartl, J.; Tews, G. Elective transfer of embryos selected on the basis of first polar body morphology is associated with increased rates of implantation and pregnancy. Fertil. Steril. 1999, 72, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Moser, M.; Sommergruber, M.; Yaman, C.; Pfleger, U.; Tews, G. First polar body morphology and blastocyst formation rate in ICSI patients. Hum. Reprod. 2002, 17, 2415–2418. [Google Scholar] [CrossRef] [PubMed]
- Fancsovits, P.; Tóthné, Z.G.; Murber, A.; Takács, F.Z.; Papp, Z.; Urbancsek, J. Correlation between first polar body morphology and further embryo development. Acta Biol. Hung. 2006, 57, 331–338. [Google Scholar] [CrossRef]
- De Santis, L.; Cino, I.; Rabellotti, E.; Calzi, F.; Persico, P.; Borini, A.; Coticchio, G. Polar body morphology and spindle imaging as predictors of oocyte quality. Reprod. Biomed. Online 2005, 11, 36–42. [Google Scholar] [CrossRef]
- Verlhac, M.-H.; Lefebvre, C.; Guillaud, P.; Rassinier, P.; Maro, B. Asymmetric division in mouse oocytes: With or without Mos. Curr. Biol. 2000, 10, 1303–1306. [Google Scholar] [CrossRef]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef]
- Giudice, L.C. Environmental toxicants: Hidden players on the reproductive stage. Fertil. Steril. 2016, 106, 791–794. [Google Scholar] [CrossRef]
- Handyside, A.H.; Kontogianni, E.H.; Hardy, K.; Winston, R.M.L. Pregnancies from biopsied human preimplantation embryos sexed by Y-specific DNA amplification. Nature 1990, 344, 768–770. [Google Scholar] [CrossRef] [PubMed]
- Verlinsky, Y.; Cieslak, J.; Ivakhnenko, V.; Evsikov, S.; Wolf, G.; White, M.; Lifchez, A.; Kaplan, B.; Moise, J.; Valle, J.; et al. Preimplantation Diagnosis of Common Aneuploidies by the First- and Second-Polar Body FISH Analysis. J. Assist. Reprod. Genet. 1998, 15, 285–289. [Google Scholar] [CrossRef]
- Verpoest, W.; Staessen, C.; Bossuyt, P.M.; Goossens, V.; Altarescu, G.; Bonduelle, M.; Devesa, M.; Eldar-Geva, T.; Gianaroli, L.; Griesinger, G.; et al. Preimplantation genetic testing for aneuploidy by microarray analysis of polar bodies in advanced maternal age: A randomized clinical trial. Hum. Reprod. 2018, 33, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, F.; Bono, S.; Biricik, A.; Nuccitelli, A.; Cotroneo, E.; Cottone, G.; Kokocinski, F.; Michel, C.E.; Minasi, M.G.; Greco, E. Application of next-generation sequencing technology for comprehensive aneuploidy screening of blastocysts in clinical pre-implantation genetic screening cycles. Hum. Reprod. 2014, 29, 2802–2813. [Google Scholar] [CrossRef]
- Kushnir, V.A.; Darmon, S.K.; Albertini, D.F.; Barad, D.H.; Gleicher, N. Effectiveness of in vitro fertilization with preimplanta-tion genetic screening: A reanalysis of United States assisted reproductive technology data 2011–2012. Fertil. Steril. 2016, 106, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Sciorio, R.; Dattilo, M. PGT-A preimplantation genetic testing for aneuploidies and embryo selection in routine ART cycles: Time to step back? Clin. Genet. 2020, 98, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Pauly, M. Single vs multiple embryo transfer: Comparative costs and a call for change. JAMA Pediatr. 2014, 168, 997–998. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.; Gusmano, M.K.; Patrizio, P. Preterm births, multiples, and fertility treatment: Recommendations for changes to policy and clinical practices. Fertil. Steril. 2014, 102, 36–39. [Google Scholar] [CrossRef]
- Ten, J.; Mendiola, J.; Vioque, J.; de Juan, J.; Bernabeu, R. Donor oocyte dysmorphisms and their influence on fertilization and embryo quality. Reprod. Biomed. Online 2007, 14, 40–48. [Google Scholar] [CrossRef]
- Esfandiari, N.; Burjaq, H.; Gotlieb, I.; Casper, R.F. Brown oocytes: Implications for assisted reproductive technology. Fertil. Steril. 2006, 86, 1522–1525. [Google Scholar] [CrossRef]
- Rienzi, L.; Ubaldi, F.M.; Iacobelli, M.; Minasi, M.G.; Romano, S.; Ferrero, S.; Sapienza, F.; Baroni, E.; Litwicka, K.; Greco, E. Significance of metaphase II human oocyte morphology on ICSI outcome. Fertil. Steril. 2008, 90, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Balaban, B.; Ata, B.; Ata, A.; Isiklar, A.; Yakin, K.; Urman, B. Severe cytoplasmic abnormalities of the oocyte decrease cry-osurvival and subsequent embryonic development of cryopreserved embryos. Hum. Reprod. 2008, 23, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Loutradis, D.; Drakakis, P.; Kallianidis, K.; Milingos, S.; Dendrinos, S.; Michalas, S. Oocyte morphology correlates with embryo quality and pregnancy rate after intracytoplasmic sperm injection. Fertil. Steril. 1999, 72, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Wilding, M.; Di Matteo, L.; D’Andretti, S.; Montanaro, N.; Capobianco, C.; Dale, B. An oocyte score for use in assisted re-production. J. Assist. Reprod. Genet. 2007, 24, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Coticchio, G.; Ezoe, K.; Lagalla, C.; Zacà, C.; Borini, A.; Kato, K. The destinies of human embryos reaching blastocyst stage between Day 4 and Day 7 diverge as early as fertilization. Hum. Reprod. 2023, 38, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Sciorio, R.; Meseguer, M. Focus on time-lapse analysis: Blastocyst collapse and morphometric assessment as new features of embryo viability. Reprod. Biomed. Online 2021, 43, 821–832. [Google Scholar] [CrossRef]
- Sciorio, R.; Campos, G.; Palini, S.; Baldini, D.; Janssens, R. Real-time image and time-lapse technology to select the single blastocyst to transfer in assisted reproductive cycles. Zygote 2023, 31, 207–216. [Google Scholar] [CrossRef]
- Bickendorf, K.; Qi, F.; Peirce, K.; Natalwala, J.; Chapple, V.; Liu, Y. Spontaneous collapse as a prognostic marker for human blastocysts: A systematic review and meta-analysis. Hum. Reprod. 2023, 1–10. [Google Scholar] [CrossRef]
- Sciorio, R.; Saura, R.H.; Thong, K.J.; Algam, M.E.; Pickering, S.J.; Meseguer, M. Blastocyst collapse as an embryo marker of low implantation potential: A time-lapse multicentre study. Zygote 2020, 28, 139–147. [Google Scholar] [CrossRef]
- Sciorio, R.; Thong, D.; Thong, K.J.; Pickering, S.J. Clinical pregnancy is significantly associated with the blastocyst width and area: A time-lapse study. J. Assist. Reprod. Genet. 2021, 38, 847–855. [Google Scholar] [CrossRef]
- Montag, M.; Koster, M.; van der Ven, K.; van der Ven, H. Gamete competence assessment by polarizing optics in assisted reproduction. Hum. Reprod. Update 2011, 17, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Swann, M.M.; Mitchison, J.M. Refinements in Polarized Light Microscopy. J. Exp. Biol. 1950, 27, 226–237. [Google Scholar] [CrossRef]
- Keefe, D.; Tran, P.; Pellegrini, C.; Oldenbourg, R. Polarized light microscopy and digital image processing identify a multilaminar structure of the hamster zona pellucida. Hum. Reprod. 1997, 12, 1250–1252. [Google Scholar] [CrossRef]
- Silva, C.P.; Kommineni, K.; Oldenbourg, R.; Keefe, D.L. The first polar body does not predict accurately the location of the metaphase II meiotic spindle in mammalian oocytes. Fertil. Steril. 1999, 71, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Squirrell, J.M.; Wokosin, D.L.; White, J.G.; Bavister, B.D. Long-term two-photon fluorescence imaging of mammalian embryos without compromising viability. Nat. Biotechnol. 1999, 17, 763–767. [Google Scholar] [CrossRef]
- Wan, Y.; McDole, K.; Keller, P.J. Light-Sheet Microscopy and Its Potential for Understanding Developmental Processes. Annu. Rev. Cell Dev. Biol. 2019, 35, 655–681. [Google Scholar] [CrossRef]
- Albert-Smet, I.; Marcos-Vidal, A.; Vaquero, J.J.; Desco, M.; Muñoz-Barrutia, A.; Ripoll, J. Applications of Light-Sheet Microscopy in Microdevices. Front. Neuroanat. 2019, 13, 1. [Google Scholar] [CrossRef]
- Wood, B.R.; Chernenko, T.; Matthäus, C.; Diem, M.; Chong, C.; Bernhard, U.; Jene, C.; Brandli, A.A.; McNaughton, D.; Tobin, M.J.; et al. Shedding New Light on the Molecular Architecture of Oocytes Using a Combination of Synchrotron Fourier Transform-Infrared and Raman Spectroscopic Mapping. Anal. Chem. 2008, 80, 9065–9072. [Google Scholar] [CrossRef]
- Rusciano, G.; Pesce, G.; Salemme, M.; Selvaggi, L.; Vaccaro, C.; Sasso, A.; Carotenuto, R. Raman spectroscopy of Xenopus laevis oocytes. Methods 2010, 51, 27–36. [Google Scholar] [CrossRef]
- Bogliolo, L.; Murrone, O.; Di Emidio, G.; Piccinini, M.; Ariu, F.; Ledda, S.; Tatone, C. Raman spectroscopy-based approach to detect aging-related oxidative damage in the mouse oocyte. J. Assist. Reprod. Genet. 2013, 30, 877–882. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciorio, R.; Cariati, F.; Fleming, S.; Alviggi, C. Exploring the Impact of Controlled Ovarian Stimulation and Non-Invasive Oocyte Assessment in ART Treatments. Life 2023, 13, 1989. https://doi.org/10.3390/life13101989
Sciorio R, Cariati F, Fleming S, Alviggi C. Exploring the Impact of Controlled Ovarian Stimulation and Non-Invasive Oocyte Assessment in ART Treatments. Life. 2023; 13(10):1989. https://doi.org/10.3390/life13101989
Chicago/Turabian StyleSciorio, Romualdo, Federica Cariati, Steven Fleming, and Carlo Alviggi. 2023. "Exploring the Impact of Controlled Ovarian Stimulation and Non-Invasive Oocyte Assessment in ART Treatments" Life 13, no. 10: 1989. https://doi.org/10.3390/life13101989
APA StyleSciorio, R., Cariati, F., Fleming, S., & Alviggi, C. (2023). Exploring the Impact of Controlled Ovarian Stimulation and Non-Invasive Oocyte Assessment in ART Treatments. Life, 13(10), 1989. https://doi.org/10.3390/life13101989