


Physiological and Biochemical Responses in Microalgae Dunaliella salina, Cylindrotheca closterium and Phormidium versicolor NCC466 Exposed to High Salinity and Irradiation

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
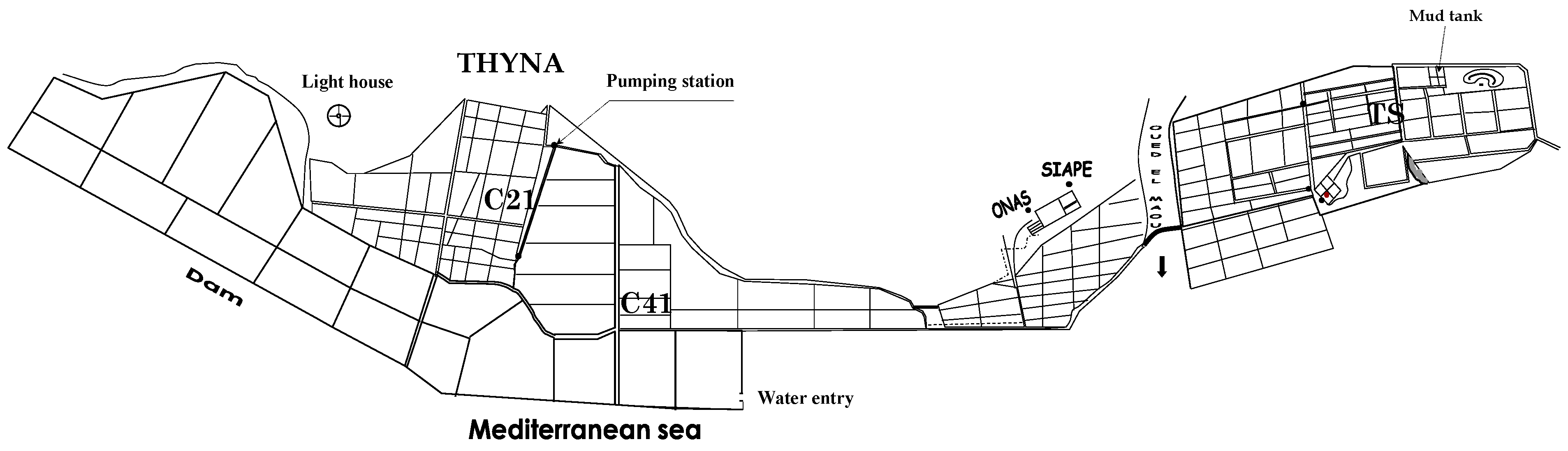
2.1. Isolation and Identification of Microalgae Species
2.2. Culture and Growth Measurement
2.3. Molecular Identification
2.3.1. DNA Extraction
2.3.2. PCR Amplification and Sequencing
2.4. Photosynthetic Activity
2.4.1. Oximetry
2.4.2. Fluorimetry Parameters
2.4.3. Antioxidative Enzyme Activities
2.4.4. Statistical Analysis
3. Results
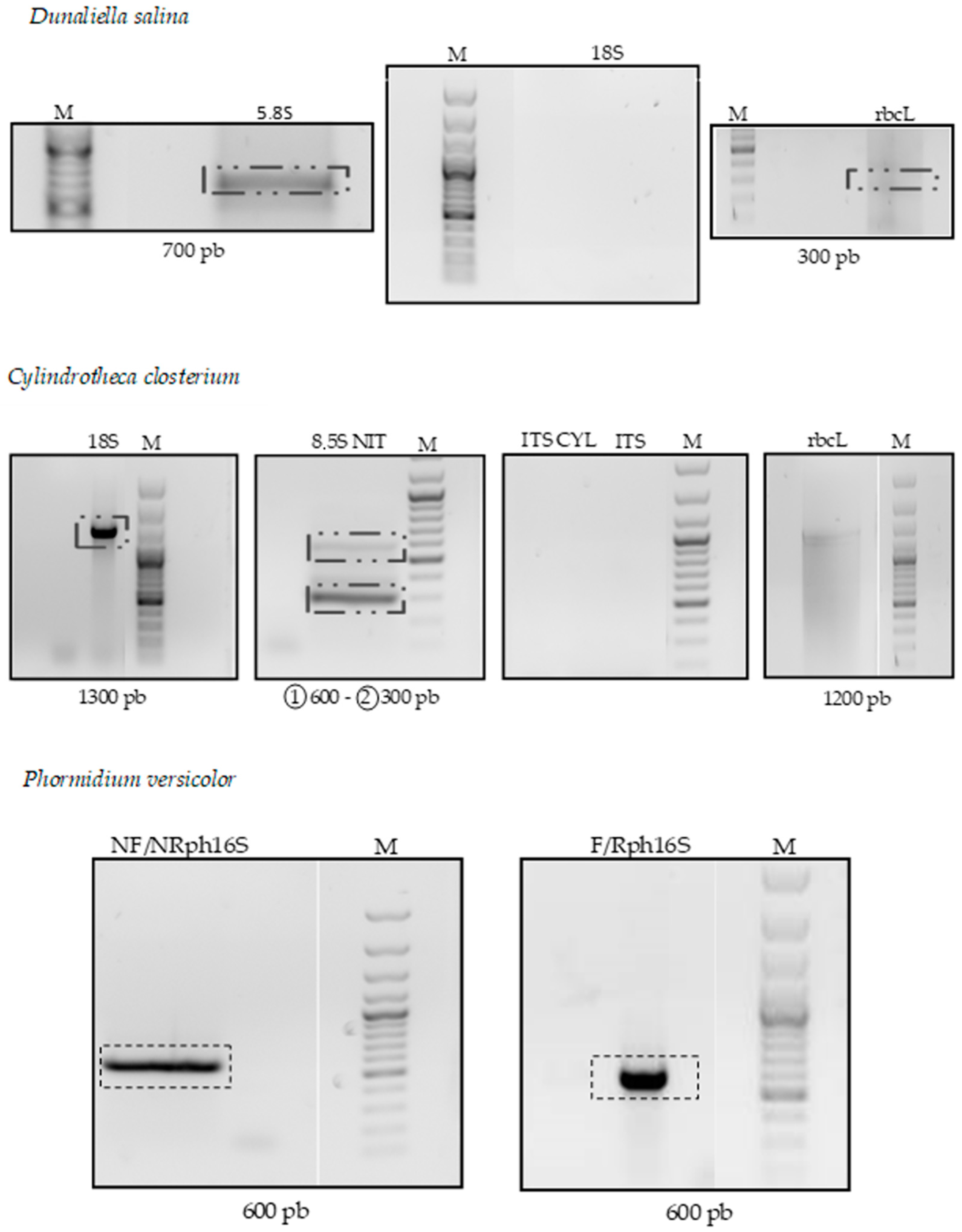
3.1. Molecular Identification
3.1.1. Dunaliella salina
3.1.2. Cylindrotheca closterium
3.1.3. Phormidium versicolor
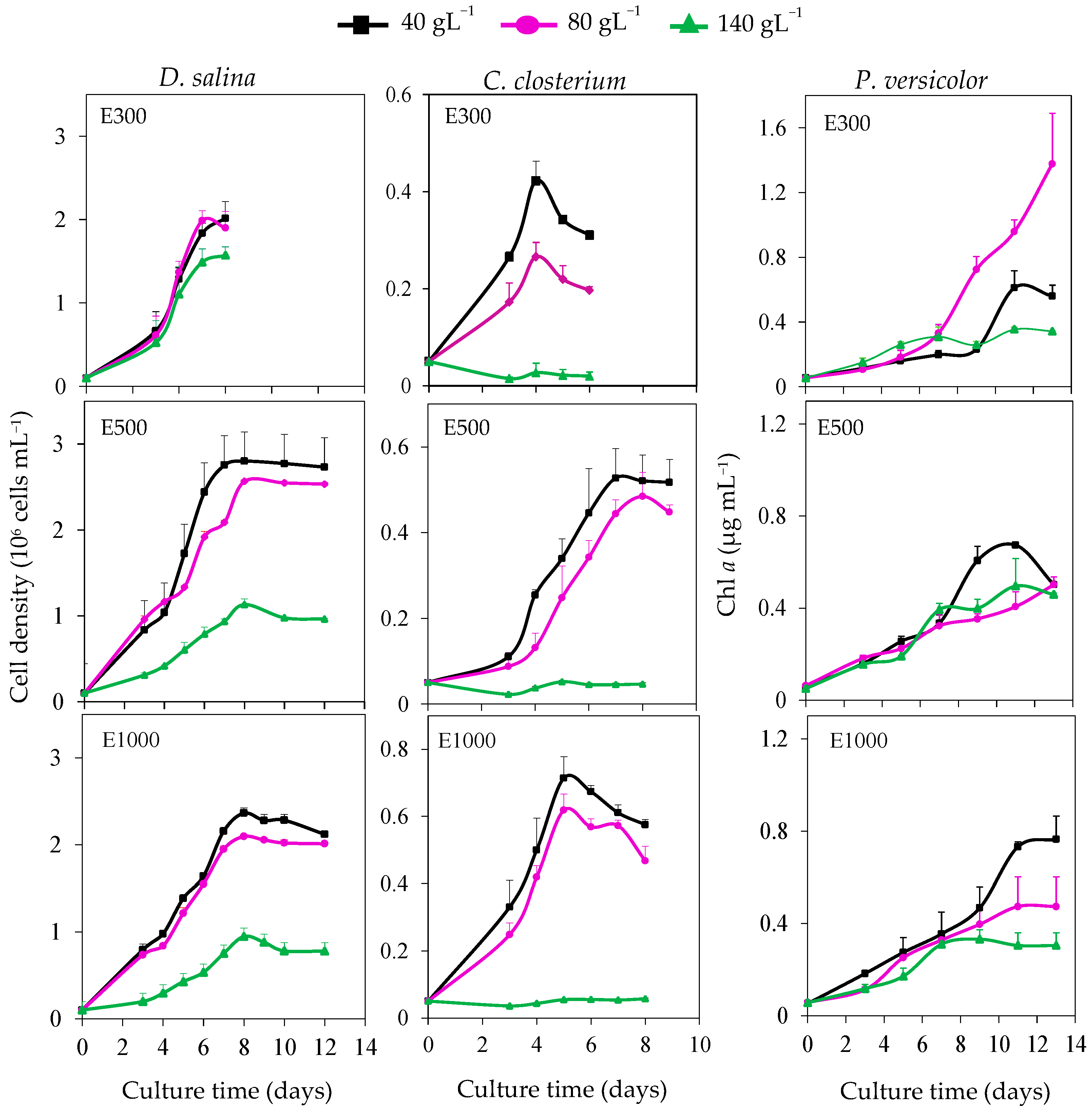
3.2. Kinetic Growth under Salt and Light Stress
3.3. Photosynthetic Activity
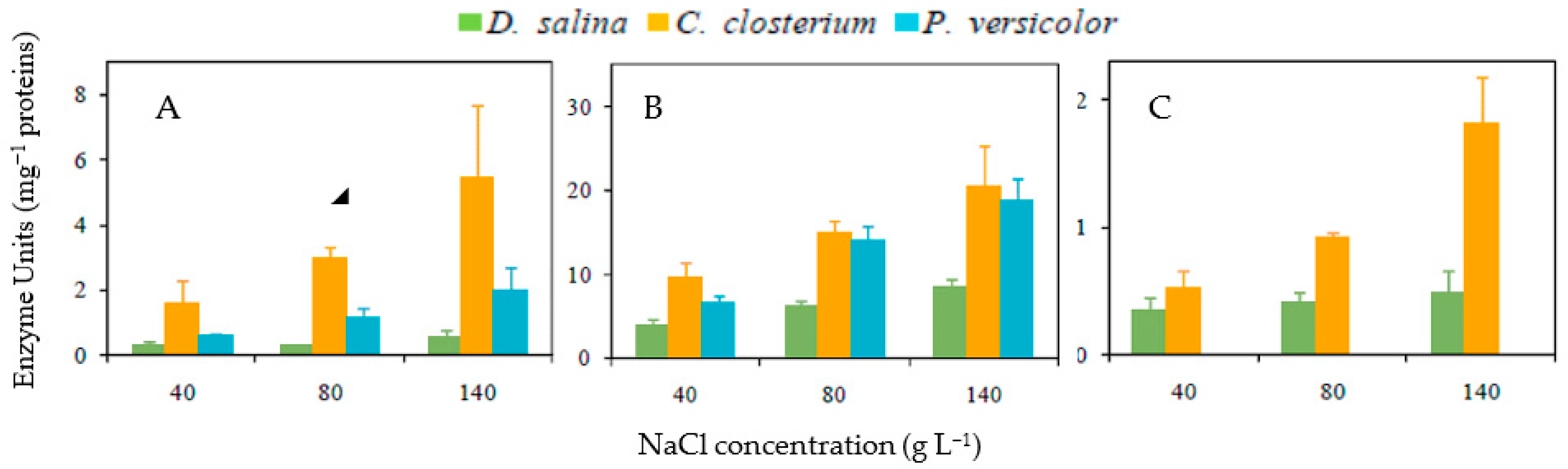
3.4. Antioxidative Enzyme Activities
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of Photosystem II under Environmental Stress. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Masmoudi, S.; Tastard, E.; Guermazi, W.; Caruso, A.; Morant-Manceau, A.; Ayadi, H. Salinity Gradient and Nutrients as Major Structuring Factors of the Phytoplankton Communities in Salt Marshes. Aquat. Ecol. 2015, 49, 1–19. [Google Scholar] [CrossRef]
- Hotos, G.N. A Preliminary Survey on the Planktonic Biota in a Hypersaline Pond of Messolonghi Saltworks (W. Greece). Diversity 2021, 13, 270. [Google Scholar] [CrossRef]
- Hotos, G.N.; Avramidou, D. The Effect of Various Salinities and Light Intensities on the Growth Performance of Five Locally Isolated Microalgae [Amphidinium carterae, Nephroselmis sp., Tetraselmis sp. (Var. Red Pappas), Asteromonas gracilis and Dunaliella sp.] in Laboratory Batch Cultures. J. Mar. Sci. Eng. 2021, 9, 1275. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Nishiyama, Y.; Miyairi, S.; Yamamoto, H.; Inagaki, N.; Kanesaki, Y.; Murata, N. Salt Stress Inhibits the Repair of Photodamaged Photosystem II by Suppressing the Transcription and Translation of PsbA Genes in Synechocystis. Plant Physiol. 2002, 130, 1443–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, A.; Kirst, G.O.; Wiencke, C.; Lehmann, H. Physiological Responses of the Antarctic Green Alga Prasiola Crispa ssp. Antarctica to Salinity Stress. J. Plant Physiol. 1991, 139, 57–62. [Google Scholar] [CrossRef]
- Mishra, A.; Mandoli, A.; Jha, B. Physiological Characterization and Stress-Induced Metabolic Responses of Dunaliella salina Isolated from Salt Pan. J. Ind. Microbiol. Biotechnol. 2008, 35, 1093–1101. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.-G. Osmotic Responses of Dunaliella to the Changes of Salinity. J. Cell. Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Roháček, K.; Bertrand, M.; Moreau, B.; Jacquette, B.; Caplat, C.; Morant-Manceau, A.; Schoefs, B. Relaxation of the Non-Photochemical Chlorophyll Fluorescence Quenching in Diatoms: Kinetics, Components and Mechanisms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130241. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, O.; Khorobrykh, S.; Tyystjärvi, E. Acclimation of Chlamydomonas reinhardtii to Extremely Strong Light. Photosyn. Res. 2021, 147, 91–106. [Google Scholar] [CrossRef]
- Melis, A. Photosystem-II Damage and Repair Cycle in Chloroplasts: What Modulates the Rate of Photodamage in Vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Canion, A.; MacIntyre, H.L.; Phipps, S. Short-Term to Seasonal Variability in Factors Driving Primary Productivity in a Shallow Estuary: Implications for Modeling Production. Estuar. Coast. Shelf Sci. 2013, 131, 224–234. [Google Scholar] [CrossRef]
- Saini, P.; Gani, M.; Kaur, J.J.; Godara, L.C.; Singh, C.; Chauhan, S.S.; Francies, R.M.; Bhardwaj, A.; Bharat Kumar, N.; Ghosh, M.K. Reactive Oxygen Species (ROS): A Way to Stress Survival in Plants. In Abiotic Stress-Mediated Sensing and Signaling in Plants: An Omics Perspective; Zargar, S.M., Zargar, M.Y., Eds.; Springer: Singapore, 2018; pp. 127–153. ISBN 978-981-10-7478-3. [Google Scholar]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, P.; Gitau, M.M.; Maróti, G. Salinity Stress Responses and Adaptation Mechanisms in Eukaryotic Green Microalgae. Cells 2019, 8, 1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tammam, A.A.; Fakhry, E.M.; El-Sheekh, M. Effect of Salt Stress on Antioxidant System and the Metabolism of the Reactive Oxygen Species in Dunaliella salina and Dunaliella tertiolecta. Afr. J. Biotechnol. 2011, 10, 3795–3808. [Google Scholar]
- Pfeil, B.E.; Schoefs, B.; Spetea, C. Function and Evolution of Channels and Transporters in Photosynthetic Membranes. Cell. Mol. Life Sci. 2014, 71, 979–998. [Google Scholar] [CrossRef] [Green Version]
- Neale, P.J.; Melis, A. Salinity-Stress Enhances Photoinhibition of Photosynthesis in Chlamydomonas reinhardtii. J. Plant Physiol. 1989, 134, 619–622. [Google Scholar] [CrossRef]
- Sharma, P.K.; Hall, D.O. Interaction of Salt Stress and Photoinhibition on Photosynthesis in Barley and Sorghum. J. Plant Physiol. 1991, 138, 614–619. [Google Scholar] [CrossRef]
- Lu, C.-M.; Zhang, J.-H. Effects of Salt Stress on PSII Function and Photoinhibition in the Cyanobacterium Spirulina Platensis. J. Plant Physiol. 1999, 155, 740–745. [Google Scholar] [CrossRef]
- Uniacke, J.; Zerges, W. Photosystem II Assembly and Repair Are Differentially Localized in Chlamydomonas. Plant Cell 2007, 19, 3640–3654. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Roy, S.; Alami, M.; Green, B.R.; Campbell, D.A. Photosystem II Photoinactivation, Repair, and Protection in Marine Centric Diatoms. Plant Physiol. 2012, 160, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lan, T.; Zhang, J.; Gao, K.; Beardall, J.; Wu, Y. Nitrogen Limitation Decreases the Repair Capacity and Enhances Photoinhibition of Photosystem II in a Diatom. Photochem. Photobiol. 2021, 97, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Ort, D.R.; Baker, N.R. A Photoprotective Role for O2 as an Alternative Electron Sink in Photosynthesis? Curr. Opin. Plant Biol. 2002, 5, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Lavaud, J.; Rousseau, B.; van Gorkom, H.J.; Etienne, A.-L. Influence of the Diadinoxanthin Pool Size on Photoprotection in the Marine Planktonic Diatom Phaeodactylum tricornutum. Plant Physiol. 2002, 129, 1398–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrine, Z.; Negi, S.; Sayre, R.T. Optimization of Photosynthetic Light Energy Utilization by Microalgae. Algal Res. 2012, 1, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Nymark, M.; Valle, K.C.; Brembu, T.; Hancke, K.; Winge, P.; Andresen, K.; Johnsen, G.; Bones, A.M. An Integrated Analysis of Molecular Acclimation to High Light in the Marine Diatom Phaeodactylum Tricornutum. PLoS ONE 2009, 4, e7743. [Google Scholar] [CrossRef] [Green Version]
- Sournia, A. Atlas du Phytoplancton Marin; Éditions du Centre National de la Recherche Scientifique; Presse du CNRS: Paris, France, 1986. [Google Scholar]
- Abid, O.; Sellami-Kammoun, A.; Ayadi, H.; Drira, Z.; Bouain, A.; Aleya, L. Biochemical Adaptation of Phytoplankton to Salinity and Nutrient Gradients in a Coastal Solar Saltern, Tunisia. Estuar. Coast. Shelf Sci. 2008, 80, 391–400. [Google Scholar] [CrossRef]
- Masmoudi, S. Dynamique du Phytoplancton et Caractérisation Physiologique et Moléculaire de Trois Espèces Autotrophes de la Saline de Sfax(Tunisie), un Milieu Extrémophile. Ph.D. Thesis, Le Mans Université, Maine, France, University of Sfax, Sfax, Tunisia, 2014. [Google Scholar]
- Barsanti, L.; Gualtieri, P. Algal Culturing. In Algae: Anatomy, Biochemistry, and Biotechnology; CRC Press: Boca Raton, FL, USA, 2014; p. 361. ISBN 978-0-429-10718-4. [Google Scholar]
- Speziale, B.J.; Schreiner, S.P.; Giammatteo, P.A.; Schindler, J.E. Comparison of N,N-Dimethylformamide, Dimethyl Sulfoxide, and Acetone for Extraction of Phytoplankton Chlorophyll. Can. J. Fish. Aquat. Sci. 1984, 41, 1519–1522. [Google Scholar] [CrossRef]
- Perni, S.; Andrew, P.W.; Shama, G. Estimating the Maximum Growth Rate from Microbial Growth Curves: Definition Is Everything. Food Microbiol. 2005, 22, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W.; Humphrey, G.F. New Spectrophotometric Equations for Determining Chlorophylls a, b, C1 and C2 in Higher Plants, Algae and Natural Phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Chamovitz, D.; Sandmann, G.; Hirschberg, J. Molecular and Biochemical Characterization of Herbicide-Resistant Mutants of Cyanobacteria Reveals That Phytoene Desaturation Is a Rate-Limiting Step in Carotenoid Biosynthesis. J. Biol. Chem. 1993, 268, 17348–17353. [Google Scholar] [CrossRef]
- Salguero, A.; de la Morena, B.; Vigara, J.; Vega, J.M.; Vilchez, C.; León, R. Carotenoids as Protective Response against Oxidative Damage in Dunaliella bardawil. Biomol. Eng. 2003, 20, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Huang, B. Réponses Physiologiques et Biochimiques à une Limitation Nutritive en Phosphore ou en Azote sur la Réorientation Métabolique des Lipides Polaires chez Différentes Espèces de Microalgues Marines. Ph.D. Thesis, Université du Maine, Le Mans, France, 2018. [Google Scholar]
- Melis, A. Solar Energy Conversion Efficiencies in Photosynthesis: Minimizing the Chlorophyll Antennae to Maximize Efficiency. Plant Sci. 2009, 177, 272–280. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of DNA from Small Amounts of Plant Tissues. BRL Focus 1990, 12, 13–15. [Google Scholar]
- Senapin, S.; Phiwsaiya, K.; Kiatmetha, P.; Withyachumnarnkul, B. Development of Primers and a Procedure for Specific Identification of the Diatom Thalassiosira Weissflogii. Aquac. Int. 2011, 19, 693–704. [Google Scholar] [CrossRef]
- Morant-Manceau, A.; Nguyen, T.L.N.; Pradier, E.; Tremblin, G. Carbonic Anhydrase Activity and Photosynthesis in Marine Diatoms. Eur. J. Phycol. 2007, 42, 263–270. [Google Scholar] [CrossRef]
- Sherwood, J.E.; Stagnitti, F.; Kokkinn, M.J.; Williams, W.D. A Standard Table for Predicting Equilibrium Dissolved Oxygen Concentrations in Salt Lakes Dominated by Sodium Chloride. Int. J. Salt Lake Res. 1992, 1, 1–6. [Google Scholar] [CrossRef]
- Elfwing, T.; Blidberg, E.; Sison, M.; Tedengren, M. A Comparison between Sites of Growth, Physiological Performance and Stress Responses in Transplanted Tridacna Gigas. Aquaculture 2003, 219, 815–828. [Google Scholar] [CrossRef]
- Rech, M.; Mouget, J.-L.; Tremblin, G. Modification of the Hansatech FMS Fluorometer to Facilitate Measurements with Microalgal Cultures. Aquat. Bot. 2003, 77, 71–80. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Meth. Enzymol. 1984, 121–126. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Classics Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Moulton, T.P.; Sommer, T.R.; Burford, M.A.; Borowitzka, L.J. Competition between Dunaliella Species at High Salinity. In Developments in Hydrobiology; Springer: Dordrecht, The Netherlands; Sao Paulo, Brazil, 1987; Volume 41, pp. 107–116. [Google Scholar]
- Pisal, D.S.; Lele, S.S. Carotenoid Production from Microalga, Dunaliella salina. Indian J. Biotechnol. 2005, 4, 476–483. [Google Scholar]
- Xu, X.-Q.; Beardall, J. Effect of Salinity on Fatty Acid Composition of a Green Microalga from an Antarctic Hypersaline Lake. Phytochemistry 1997, 45, 655–658. [Google Scholar] [CrossRef]
- Kan, G.; Shi, C.; Wang, X.; Xie, Q.; Wang, M.; Wang, X.; Miao, J. Acclimatory Responses to High-Salt Stress in Chlamydomonas (Chlorophyta, Chlorophyceae) from Antarctica. Acta Oceanol. Sin. 2012, 31, 116–124. [Google Scholar] [CrossRef]
- Waditee, R.; Hibino, T.; Tanaka, Y.; Nakamura, T.; Incharoensakdi, A.; Takabe, T. Halotolerant Cyanobacterium Aphanothece HalophyticaContains an Na+/H+ Antiporter, Homologous to Eukaryotic Ones, with Novel Ion Specificity Affected by C-Terminal Tail. J. Biol. Chem. 2001, 276, 36931–36938. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, N.; Fulda, S.; Hagemann, M. Glucosylglycerol Accumulation during Salt Acclimation of Two Unicellular Cyanobacteria. J. Gen. Microbiol. 1992, 138, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Allakhverdiev, S.I.; Kinoshita, M.; Inaba, M.; Suzuki, I.; Murata, N. Unsaturated Fatty Acids in Membrane Lipids Protect the Photosynthetic Machinery against Salt-Induced Damage in Synechococcus. Plant Physiol. 2001, 125, 1842–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saros, J.E.; Fritz, S.C. Nutrients as a Link between Ionic Concentration/Composition and Diatom Distributions in Saline Lakes. J. Paleolimnol. 2000, 23, 449–453. [Google Scholar] [CrossRef]
- Xu, Y.; Harvey, P.J. Carotenoid Production by Dunaliella salina under Red Light. Antioxidants 2019, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.I.; González, M.A. The Effect of Temperature and Irradiance on the Growth and Carotenogenic Capacity of Seven Strains of Dunaliella salina (Chlorophyta) Cultivated under Laboratory Conditions. Biol. Res. 2005, 38, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative Damage and Antioxidative System in Algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Hotos, G.N.; Antoniadis, T.I. The Effect of Colored and White Light on Growth and Phycobiliproteins, Chlorophyll and Carotenoids Content of the Marine Cyanobacteria Phormidium Sp. and Cyanothece Sp. in Batch Cultures. Life 2022, 12, 837. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Singh, V.P.; Prasad, S.M. NaCl-Induced Physiological and Biochemical Changes in Two Cyanobacteria Nostoc Muscorum and Phormidium Foveolarum Acclimatized to Different Photosynthetically Active Radiation. J. Photochem. Photobiol. B Biol. 2015, 151, 221–232. [Google Scholar] [CrossRef]
- Janssen, M.; Janssen, M.; de Winter, M.; Tramper, J.; Mur, L.R.; Snel, J.; Wijffels, R.H. Efficiency of Light Utilization of Chlamydomonas reinhardtii under Medium-Duration Light/Dark Cycles. J. Biotechnol. 2000, 78, 123–137. [Google Scholar] [CrossRef]
- Page, L.E.; Liberton, M.; Pakrasi, H.B. Reduction of Photoautotrophic Productivity in the Cyanobacterium Synechocystis Sp. Strain PCC 6803 by Phycobilisome Antenna Truncation. Appl. Environ. Microbiol. 2012, 78, 6349–6351. [Google Scholar] [CrossRef] [Green Version]
- Vonshak, A.; Kancharaksa, N.; Bunnag, B.; Tanticharoen, M. Role of Light and Photosynthesis on the Acclimation Process of the CyanobacteriumSpirulina Platensis to Salinity Stress. J. Appl. Phycol. 1996, 8, 119–124. [Google Scholar] [CrossRef]
- Zeng, M.-T.; Vonshak, A. Adaptation of Spirulina Platensis to Salinity-Stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 120, 113–118. [Google Scholar] [CrossRef]
- Berry, S.; Bolychevtseva, Y.V.; Rögner, M.; Karapetyan, N.V. Photosynthetic and Respiratory Electron Transport in the Alkaliphilic Cyanobacterium Arthrospira (spirulina) platensis. Photosyn. Res. 2003, 78, 67–76. [Google Scholar] [CrossRef]
- Gilmour, D.J.; Hipkins, M.F.; Webber, A.N.; Baker, N.R.; Boney, A.D. The Effect of Ionic Stress on Photosynthesis in Dunaliella tertiolecta: Chlorophyll Fluorescence Kinetics and Spectral Characteristics. Planta 1985, 163, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Balnokin, Y.; Popova, L.; Gimmler, H. Further Evidence for an ATP-Driven Sodium Pump in the Marine Alga Tetraselmis (Platymonas) Viridis. J. Plant Physiol. 1997, 150, 264–270. [Google Scholar] [CrossRef]
- Shono, M.; Hara, Y.; Wada, M.; Fujii, T. A Sodium Pump in the Plasma Membrane of the Marine Alga Heterosigma Akashiwo. Plant Cell Physiol. 1996, 37, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Popova, L.G.; Shumkova, G.A.; Andreev, I.M.; Balnokin, Y.V. Functional Identification of Electrogenic Na+ -Translocating ATPase in the Plasma Membrane of the Halotolerant Microalga Dunaliella maritima. FEBS Lett. 2005, 579, 5002–5006. [Google Scholar] [CrossRef]
- Gordillo, F.J.L.; Jimenez, C.; Chavarrıa, J.; Niell, F.X. Photosynthetic Acclimation to Photon Irradiance and Its Relation to Chlorophyll Fluorescence and Carbon Assimilation in the Halotolerant Green Alga Dunaliella Viridis. Photosyn. Res. 2001, 68, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Osmond, C. What Is Photoinhibition? Some Insights from Sun and Shade Plants. In Photoinhibition of Photosynthesis: From the Molecular Mechanisms to the Field; BIOS Scientific Publishing: Oxford, UK, 1994; pp. 1–24. [Google Scholar]
- Björkman, O.; Demmig, B. Photon Yield of O2 Evolution and Chlorophyll Fluorescence Characteristics at 77 K among Vascular Plants of Diverse Origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Dhiab, B. Photosynthetic Behavior of Microalgae in Response to Environmental Factors. In Applied Photosynthesis; Najafpour, M., Ed.; InTech: London, UK, 2012; pp. 23–46. [Google Scholar]
- Kaňa, R.; Prášil, O.; Komárek, O.; Papageorgiou, G.C. Govindjee Spectral Characteristic of Fluorescence Induction in a Model Cyanobacterium, Synechococcus Sp. (PCC 7942). Biochim. Biophys. Acta Bioenerg. 2009, 1787, 1170–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinetova, M.A.; Mironov, K.S.; Mustardy, L.; Shapiguzov, A.; Bachin, D.; Allakhverdiev, S.I.; Los, D.A. Aquaporin-Deficient Mutant of Synechocystis Is Sensitive to Salt and High-Light Stress. J. Photochem. Photobiol. B Biol. 2015, 152, 377–382. [Google Scholar] [CrossRef]
- Zakhozhii, I.G.; Matalin, D.A.; Popova, L.G.; Balnokin, Y.V. Responses of Photosynthetic Apparatus of the Halotolerant Microalga Dunalliella Maritima to Hyperosmotic Salt Shock. Russ. J. Plant Physiol. 2012, 59, 42–49. [Google Scholar] [CrossRef]
- Bukhov, N.; Carpentier, R. Alternative Photosystem I-Driven Electron Transport Routes: Mechanisms and Functions. Photosyn. Res. 2004, 82, 17–33. [Google Scholar] [CrossRef]
- Liska, A.J.; Shevchenko, A.; Pick, U.; Katz, A. Enhanced Photosynthesis and Redox Energy Production Contribute to Salinity Tolerance in Dunaliella as Revealed by Homology-Based Proteomics. Plant Physiol. 2004, 136, 2806–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. Saltern Evaporation Ponds as Model Systems for the Study of Primary Production Processes under Hypersaline Conditions. Aquat. Microb. Ecol. 2009, 56, 193–204. [Google Scholar] [CrossRef]
- Yao, C.-H.; Ai, J.-N.; Cao, X.-P.; Xue, S. Salinity Manipulation as an Effective Method for Enhanced Starch Production in the Marine Microalga Tetraselmis Subcordiformis. Bioresour. Technol. 2013, 146, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Thaipratum, R.; Melis, A.; Svasti, J.; Yokthongwattana, K. Analysis of Non-Photochemical Energy Dissipating Processes in Wild Type Dunaliella salina (Green Algae) and in Zea1, a Mutant Constitutively Accumulating Zeaxanthin. J. Plant Res. 2009, 122, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, P.; Di Rienzo, J.; Fernandez, L.; Hopp, H.E.; Paniego, N.; Heinz, R.A. Transcriptomic Identification of Candidate Genes Involved in Sunflower Responses to Chilling and Salt Stresses Based on CDNA Microarray Analysis. BMC Plant Biol. 2008, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, B.; Zhang, A.; Liu, Z. Oxidative Stress in Nostoc Flagelliforme Subjected to Desiccation and Effects of Exogenous Oxidants on Its Photosynthetic Recovery. J. Appl. Phycol. 2003, 15, 445–450. [Google Scholar] [CrossRef]
- Nguyen-Deroche, T.L.N.; Caruso, A.; Le, T.T.; Bui, T.V.; Schoefs, B.; Tremblin, G.; Morant-Manceau, A. Zinc Affects Differently Growth, Photosynthesis, Antioxidant Enzyme Activities and Phytochelatin Synthase Expression of Four Marine Diatoms. Sci. World J. 2012, 2012, 982957. [Google Scholar] [CrossRef] [Green Version]
- Sung, M.-S.; Hsu, Y.-T.; Hsu, Y.-T.; Wu, T.-M.; Lee, T.-M. Hypersalinity and Hydrogen Peroxide Upregulation of Gene Expression of Antioxidant Enzymes in Ulva fasciata against Oxidative Stress. Mar. Biotechnol. 2009, 11, 199–209. [Google Scholar] [CrossRef]
- Bamary, Z.; Einali, A. Changes in Carbon Partitioning and Pattern of Antioxidant Enzyme Activity Induced by Arginine Treatment in the Green Microalga Dunaliella salina under Long-Term Salinity. Microb. Ecol. 2022, 84, 198–212. [Google Scholar] [CrossRef]
- Rijstenbil, J.W. UV- and Salinity-Induced Oxidative Effects in the Marine Diatom Cylindrotheca Closterium during Simulated Emersion. Mar. Biol. 2005, 147, 1063–1073. [Google Scholar] [CrossRef]
- Murik, O.; Kaplan, A. Paradoxically, Prior Acquisition of Antioxidant Activity Enhances Oxidative Stress-Induced Cell Death. Environ. Microbiol. 2009, 11, 2301–2309. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer a | Sequence (5’-3’) | Temperature of Hybridation (°C) |
|---|---|---|
| Fph16S | GCAAGCGTTATCCGGAATKAT | 61 |
| Rph16S | CCTGTGTTCGCGCTCCCGAA | 63 |
| NFph16S | TTATCCGGAATKATTGGGCGT | 61 |
| NRph16S | GTTCGCGCTCCCGAAGGCAC | 62 |
| F2NL18S | GCGCACCAAGGTAATGATTAA | 64 |
| R2NL18S | TTAATCATTACCTTGGTGCGC | 60 |
| 18SF | CCGGCGATGGATCATTCAAGT | 58 |
| 18SR | TTCACCGGACCATTCAATCGG | 58 |
| rbcL1 | AAGGAGAAATHAATGTCT | 52 |
| rbcL7 | AARCAACCTTGTGTAAGTCTC | 52 |
| ITS1 | TCCGTAGGTGAACCTGCGG | 52 |
| ITS2 | TCCTCCGCTTATTGATATGC | 52 |
| FDSITS2 | AGGCTAGCTCAAGGACCCGT | 64 |
| RDSITS2 | AGGGCCGAGCCCATGGTCC | 66 |
| FNitITS2 | TACAACTTTCAGCGGTGGAT | 58 |
| RNitITS2 | TACCAGAGATAGGACGAGGA | 56 |
| FcylITS2 | TAACAAGGTTCCGTAGTGAA | 56 |
| RcylITS2 | TAGCACAAAGGCTACTCTCA | 58 |
| Parameters | Dunaliella salina | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Irradiance (μmol Photons m−2 s−1) | 300 | 500 | 1000 | ||||||
| NaCl (g L−1) | 40 | 80 | 140 | 40 | 80 | 140 | 40 | 80 | 140 |
| Maximum growth rate (day −1) | 0.53 ± 0.15 aB | 0.61 ± 0.15 aC | 0.57 ± 0.23 bG | 1.06 ± 0.08 cA | 0.95 ± 0.01 cD | 0.36 ± 0.05 dF | 0.91 ± 0.04 eA | 0.80 ± 0.01 eE | 0.27 ± 0.06 fF |
| Maximum cell density (106 cells mL−1) | 2.02 ± 0.15 aA | 2.00 ± 0.12 bD | 1.61 ± 0.10 cF | 2.89 ± 0.16 dB | 2.57 ± 0.02 eE | 1.13 ± 0.07 fG | 2.37 ± 0.05 gC | 2.09 ± 0.01 hD | 0.95 ± 0.01 iH |
| Chl a (µg mL−1) | 2.55 ± 0.17 aA | 1.93 ± 0.18 bB | 1.62 ± 0.15 bE | 3.46 ± 0.31 cA | 2.92 ± 0.15 dB | 1.67 ± 0.13 eD | 2.41 ± 0.32 fA | 3.10 ± 0.38 fC | 1.56 ± 0.09 gD |
| Chl b (µg mL−1) | 0.60 ± 0.14 aA | 0.50 ± 0.10 aC | 0.48 ± 0.05 aE | 0.54 ± 0.08 bB | 0.56 ± 0.18 bC | 0.32 ± 0.05 bD | 0.38 ± 0.04 cdB | 0.58 ± 0.12 cC | 0.35 ± 0.09 dD |
| Carotenoids (µg mL−1) | 1.56 ± 0.74 aA | 2.57 ± 0.27 aC | 4.65 ± 0.45 bE | 3.35 ± 0.63 cAB | 4.01 ± 1.65 cC | 7.94 ± 1.21 dF | 4.35 ± 1.07 eB | 7.36 ± 1.97 eD | 14.82 ± 1.46 fG |
| Antenna size (Chl a/ Chl b) | 4.4 ± 0.93 aA | 5.31 ± 0.95 aB | 6.29 ± 1.09 aC | 4.90 ± 0.68 bA | 5.57 ± 1.96 bB | 4.95 ± 0.58 bC | 4.10 ± 0.79 cA | 3.53 ± 1.06 cB | 4.65 ± 1.32 cC |
| Cylindrotheca closterium | |||||||||
| Maximum growth rate (day−1) | 0.46 ± 0.07 aA | 0.22 ± 0.05 aB | 0.04 ± 0.02 bE | 0.40 ± 0.05 cA | 0.35 ± 0.02 cC | 0.00 ± 0.00 dD | 0.22 ± 0.12 eA | 0.22 ± 0.02 eBC | 0.0 ± 0.0 fD |
| Maximum cell density (106 cells mL−1) | 0.42 ± 0.04 aA | 0.30 ± 0.02 bC | 0.22 ± 0.02 cG | 0.53 ± 0.07 dA | 0.49 ± 0.05 eD | 0.06 ± 0.01 fF | 0.72 ± 0.06 aB | 0.62± 0.05 bE | 0.06 ± 0.01 cF |
| Chl a (µg mL−1) | 0.97 ± 0.20 aA | 0.86 ± 0.08 aC | 0.56 ± 0.04 bE | 0.95 ± 0.02 cAB | 0.86 ± 0.08 cD | 0 ± 0 dF | 1.13 ± 0.09 eB | 0.89 ± 0.07 eD | 0 ± 0 fF |
| Chl c (µg mL−1) | 0.40 ± 0.05 aA | 0.26 ± 0.06 aC | 0.25 ± 0.06 bE | 0.21 ± 0.02 cB | 0.17 ± 0.05 cD | 0 ± 0 dF | 0.27 ± 0.05 eB | 0.24 ± 0.03 eD | 0 ± 0 fF |
| Fucoxanthin (µg mL−1) | 0.51 ± 0.02 aA | 0.51 ± 0.02 aB | 0.30 ± 0.01 bD | 0.47 ± 0.13 cA | 0.42 ± 0.02 cC | 0 ± 0 dE | 0.45 ± 0.16 eA | 0.51 ± 0.14 eC | 0 ± 0 fE |
| Antenna size (Chl a/ Chl c) | 2.38 ± 0.19 aA | 3.39 ± 0.61 aC | 2.29 ± 0.55 bD | 4.63 ± 0.36 cB | 5.33 ± 1.22 cC | 0 ± 0 a2 dE | 4.20 ± 0.58 eB | 3.78 ± 0.16 eC | 0 ± 0 fE |
| Phormidium versicolor | |||||||||
| Maximum growth rate (day−1) | 0.61 ± 0.07 aA | 0.66 ± 0.04 aC | 0.29 ± 0.01 aD | 0.45 ± 0.04 bA | 0.27 ±0.02 bB | 0.31 ±0.03 bD | 0.35 ± 0.06 cA | 0.32 ± 0.04 cB | 0.27 ± 0.03 cD |
| Chl a (µg mL−1) | 1.55 ± 0.23 aA | 0.77 ± 0.23 aB | 0.71 ± 0.16 bD | 1.38 ± 0.49 cA | 1.31 ± 0.41 cB | 0.17± 0.06 dE | 0.94 ± 0.26 eA | 2.12 ± 0.03 eC | 0.46 ± 0.10 fD |
| Carotenoids (µg mL−1) | 0.13 ± 0.01 aA | 0.19 ± 0.01 bCD | 0.30 ± 0.01 bF | 0.25 ± 0.01 cB | 0.41 ± 0.03 dC | 0.21 ± 0.13 dE | 0.14 ± 0.05 eA | 0.56 ± 0.08 fD | 0.34 ± 0.01 fE |
| Parameters | Dunaliella salina | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Irradiance (μmol Photons m−2 s−1) | 300 | 500 | 1000 | ||||||
| NaCl (g L−1) | 40 | 80 | 140 | 40 | 80 | 140 | 40 | 80 | 140 |
| PN (μmol O2 h−1 mg−1 Chl a) | 728 ± 35 aA | 649 ± 53 aD | 467 ± 32 bE | 1017 ± 36 cB | 711 ± 120 dD | 447 ± 27 eE | 840 ± 27 fC | 668 ± 52 gD | 367 ± 28 hF |
| Fv/Fm | 0.71 ± 0.02 abA | 0.75 ± 0.01 bB | 0.74 ± 0.04 aC | 0.75 ± 0.02 bA | 0.67 ± 0.12 bB | 0.75 ± 0.01 bC | 0.78 ± 0.03 aA | 0.75 ± 0.01 aB | 0.64 ± 0.02 bD |
| ΦPSII | 0.32 ± 0.09 aA | 0.28 ± 0.13 aC | 0.37 ± 0.17 aD | 0.31 ± 0.02 bA | 0.22 ± 0.01 cC | 0.23 ± 0.01 bcE | 0.23 ± 0.02 deB | 0.16 ± 0.01 dC | 0.11 ± 0.01 eE |
| NPQ | 0.23 ± 0.11 aA | 0.23 ± 0.06 bC | 0.49 ± 0.19 cD | 0.74 ± 0.12 dA | 1.01 ± 0.07 cC | 0.71 ± 0.12 dE | 1.3 ± 0.07 fB | 2.48 ± 0.44 eC | 5.97 ± 0.39 eE |
| Cylindrotheca closterium | |||||||||
| PN (μmol O2 h−1 mg−1 Chl a) | 387 ± 69 aA | 343 ± 19 abB | 243 ± 32 b | 427 ± 39 cA | 365 ± 53 cB | nd | 425 ± 34 dA | 287 ± 71 eB | nd |
| Fv/Fm | 0,71 ± 0.02 abA | 0.74 ± 0.09 aB | 0.76 ± 0.11 b | 0,68 ± 0.04 cA | 0,76 ± 0.02 cB | nd | 0.75 ± 0.03 dA | 0.73 ± 0.04 dB | nd |
| ΦPSII | 0.51 ± 0.05 aA | 0.39 ± 0.13 aD | 0.20 ± 0.08 ab | 0.25 ± 0.03 cB | 0.37 ± 0.02 dE | nd | 0.20 ± 0.03 eC | 0.30 ± 0.01 eF | nd |
| NPQ | 0.51 ± 0.10 aA | 0.31± 0.18 aC | 8.05 ± 2.24 b | 4.18 ± 0.19 cAB | 10.89 ± 2.47 cB | nd | 7.76 ± 0.15 dB | 21.31 ± 4.63 dD | nd |
| Phormidium versicolor | |||||||||
| PN (μmol O2 h−1 mg−1 Chla) | 552 ± 66 aA | 432 ± 26 aB | 278 ± 42 bC | 421 ± 52 cA | 370 ± 79 cdB | 274 ± 28 dC | 474 ± 46 eA | 372 ± 33 fB | 279 ± 37 fC |
| Fv/Fm | 0.39 ± 0.04 aA | 0.46 ± 0.08 aCD | 0.50 ± 0.07 aE | 0.35 ± 0.05 bA | 0.33 ± 0.04 bC | 0.45 ± 0.09 bE | 0.55 ± 0.07 cB | 0.55 ± 0.10 cD | 0.50 ± 0.19 cE |
| ΦPSII | 0.28 ± 0.04 aB | 0.26 ± 0.03 aC | 0.52 ± 0.03 bF | 0.15 ± 0.05 cA | 0.16 ± 0.01 cD | 0.21 ± 0.05 cG | 0.14 ± 0.05 eA | 0.38 ± 0.04 dE | 0.47 ± 0.09 dF |
| NPQ | 0.12 ± 0.03 aA | 0.19 ± 0.13 aC | 0.21± 0.14 aE | 0.14 ± 0.07 bAB | 0.15 ± 0.02 bC | 0.67 ± 0.24 cE | 0.65 ± 0.35 dB | 1.5 ± 0.87 dB | 0.33 ± 0.21 dE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guermazi, W.; Masmoudi, S.; Trabelsi, N.A.; Gammoudi, S.; Ayadi, H.; Morant-Manceau, A.; Hotos, G.N. Physiological and Biochemical Responses in Microalgae Dunaliella salina, Cylindrotheca closterium and Phormidium versicolor NCC466 Exposed to High Salinity and Irradiation. Life 2023, 13, 313. https://doi.org/10.3390/life13020313
Guermazi W, Masmoudi S, Trabelsi NA, Gammoudi S, Ayadi H, Morant-Manceau A, Hotos GN. Physiological and Biochemical Responses in Microalgae Dunaliella salina, Cylindrotheca closterium and Phormidium versicolor NCC466 Exposed to High Salinity and Irradiation. Life. 2023; 13(2):313. https://doi.org/10.3390/life13020313
Chicago/Turabian StyleGuermazi, Wassim, Salma Masmoudi, Neila Annabi Trabelsi, Sana Gammoudi, Habib Ayadi, Annick Morant-Manceau, and George N. Hotos. 2023. "Physiological and Biochemical Responses in Microalgae Dunaliella salina, Cylindrotheca closterium and Phormidium versicolor NCC466 Exposed to High Salinity and Irradiation" Life 13, no. 2: 313. https://doi.org/10.3390/life13020313
APA StyleGuermazi, W., Masmoudi, S., Trabelsi, N. A., Gammoudi, S., Ayadi, H., Morant-Manceau, A., & Hotos, G. N. (2023). Physiological and Biochemical Responses in Microalgae Dunaliella salina, Cylindrotheca closterium and Phormidium versicolor NCC466 Exposed to High Salinity and Irradiation. Life, 13(2), 313. https://doi.org/10.3390/life13020313