
Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars



Abstract
:1. Introduction
2. Materials and Methods
2.1. Propagation of Rosemary Cuttings
2.2. AMF Trials Design
2.3. Plant Growth Measurements
2.4. Total Antioxidant Content and Phenolic Content Assays
2.5. Root Staining and Microscopy
2.6. RNA Extraction from Rosemary Leaves
2.7. Reverse Transcription and Quantitative PCR
2.8. Statistical Analysis
3. Results
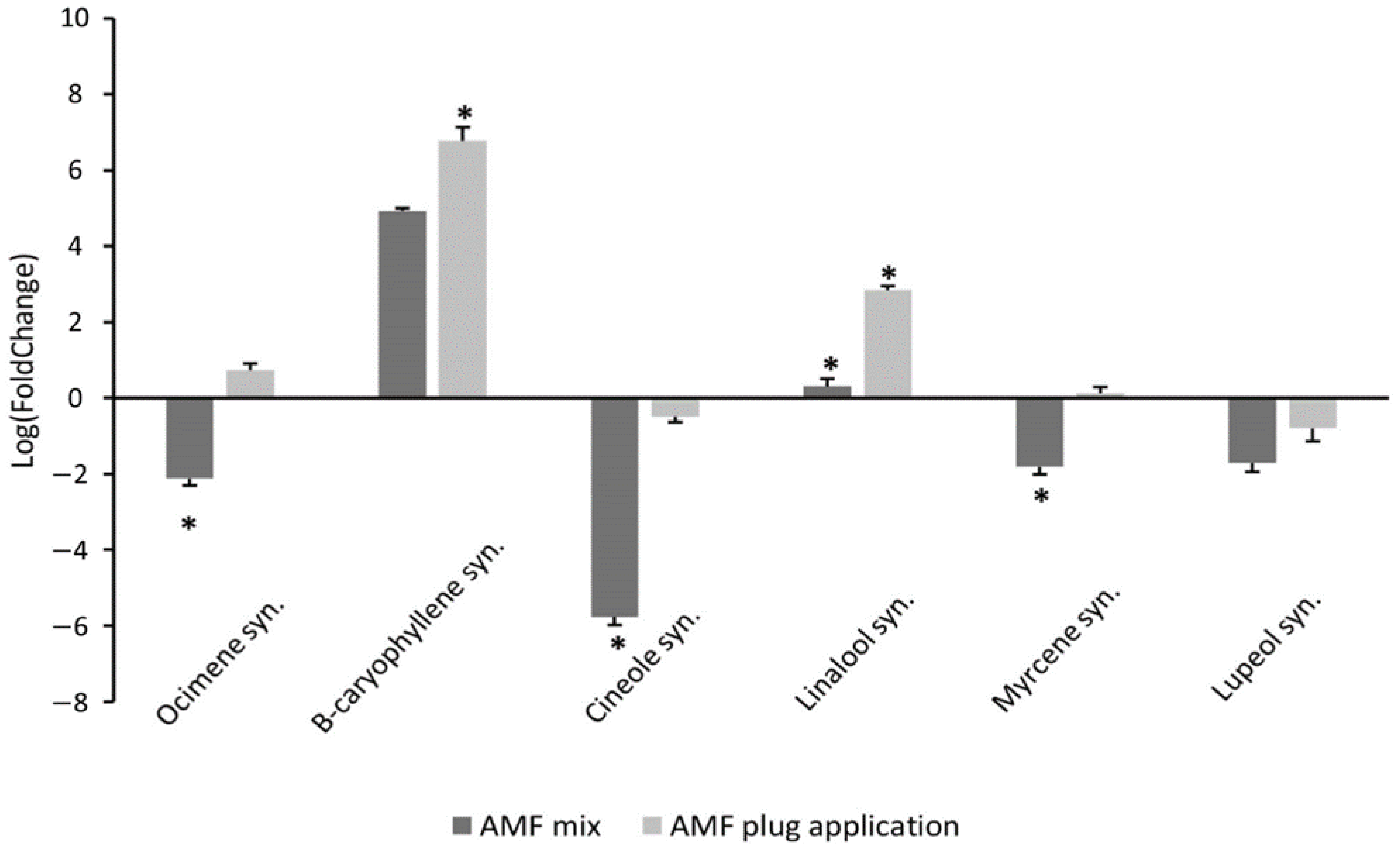
3.1. AMF Addition to the Substrate Increases Phenolic Content and Alters Gene Expression of Terpene Synthases without Altering the Biomass or Morphology of Rosemary ‘Perigord’
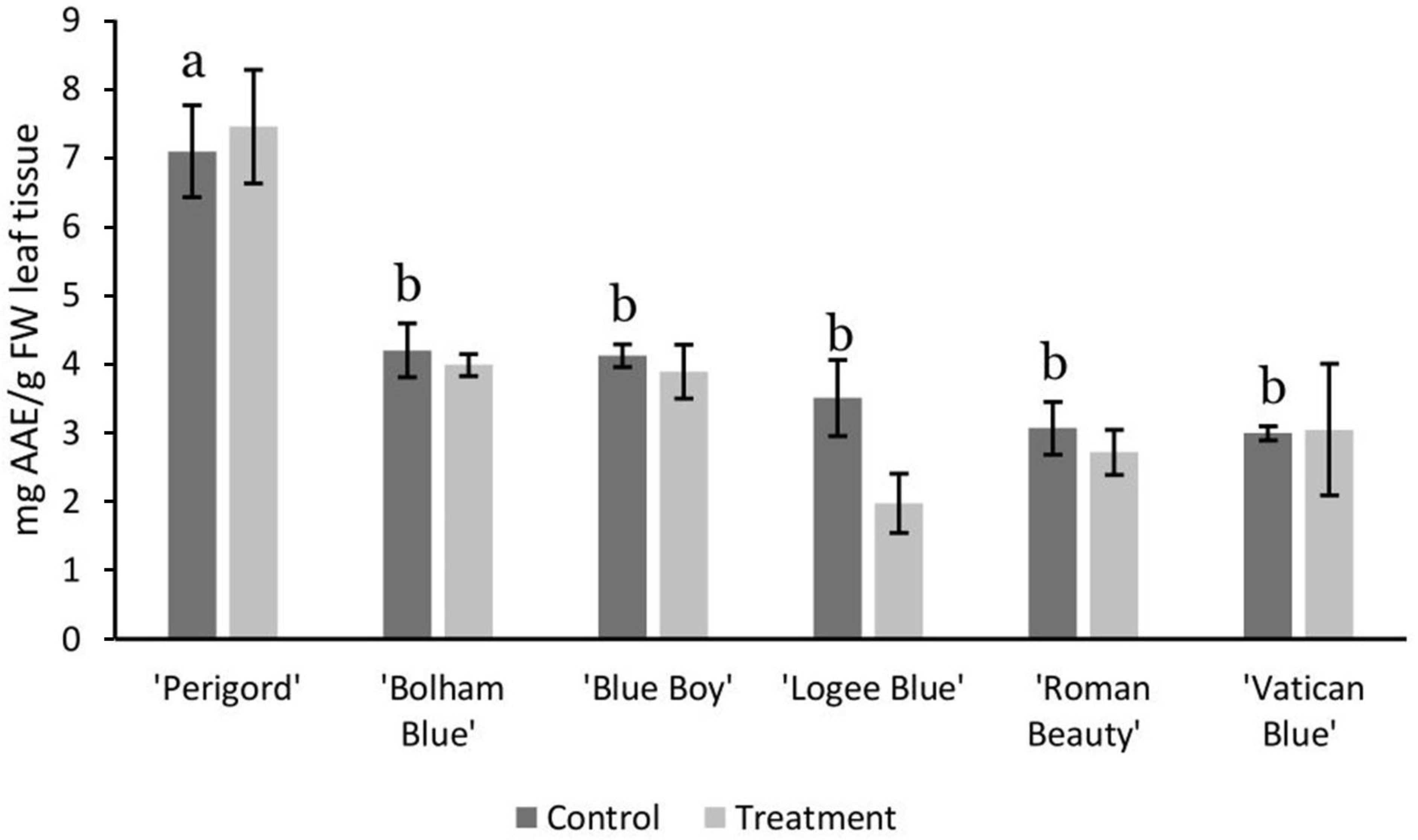
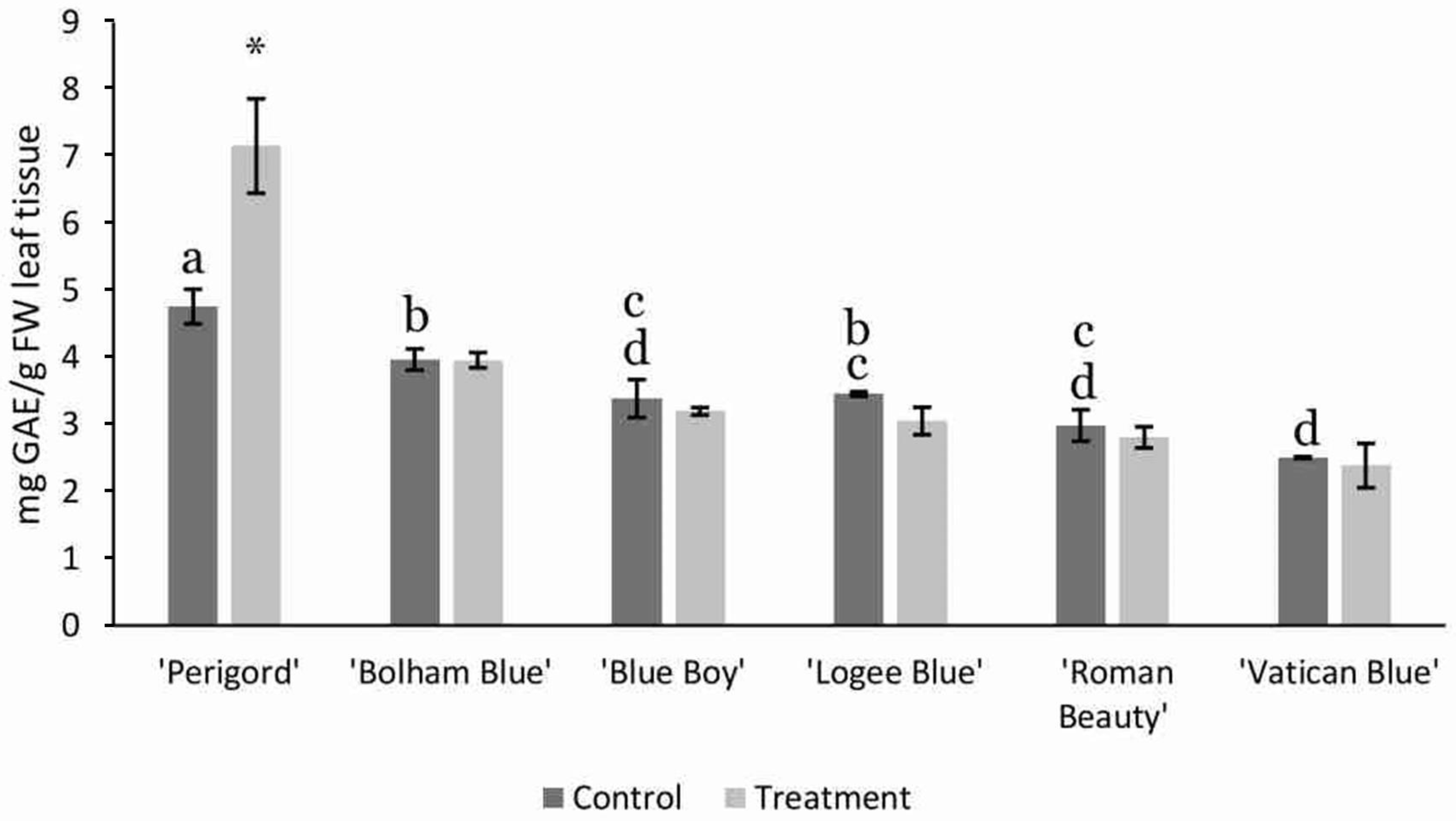
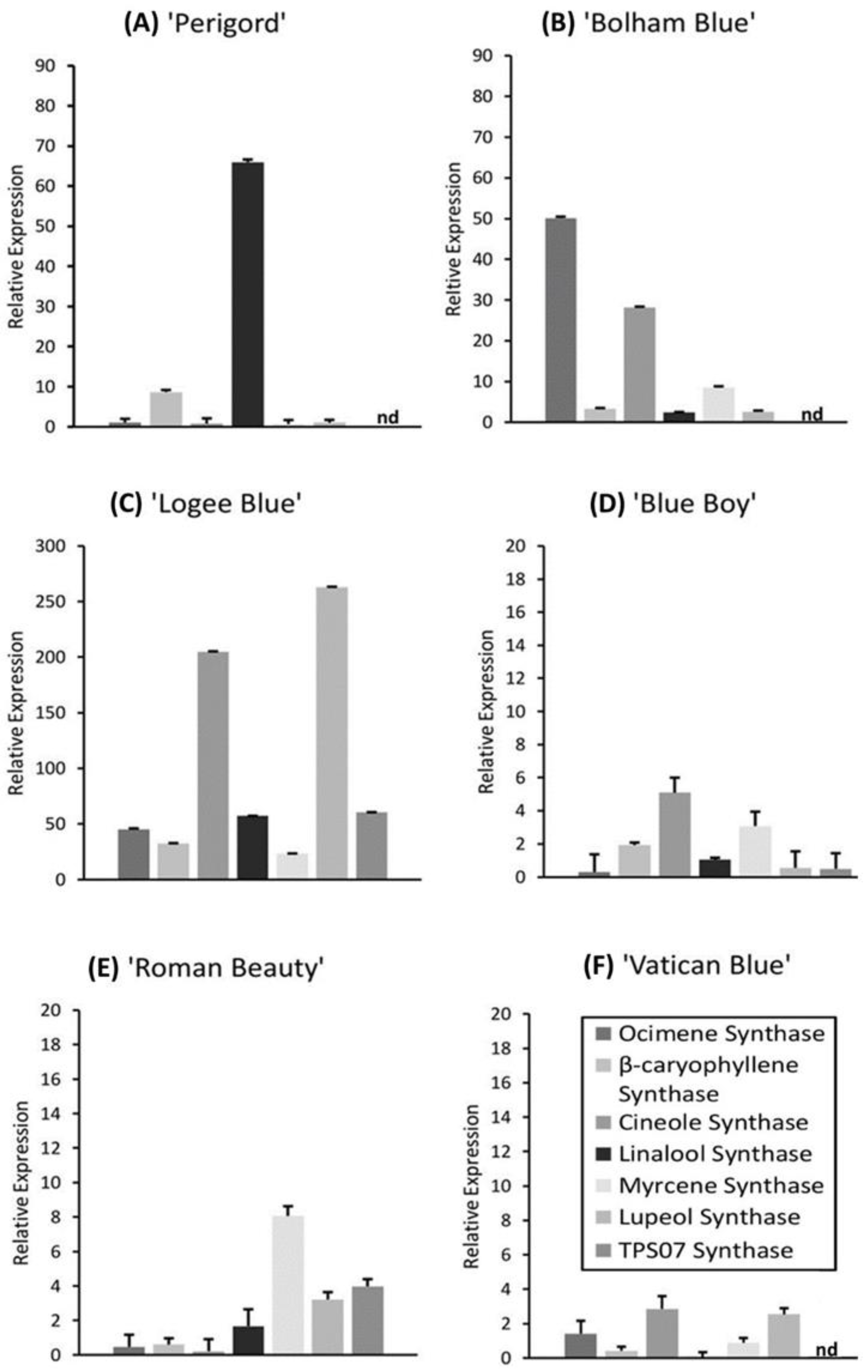
3.2. AMF Substrate Mixture Alters Gene Expression of Terpene Synthases in Six Rosemary Cultivars
4. Discussion
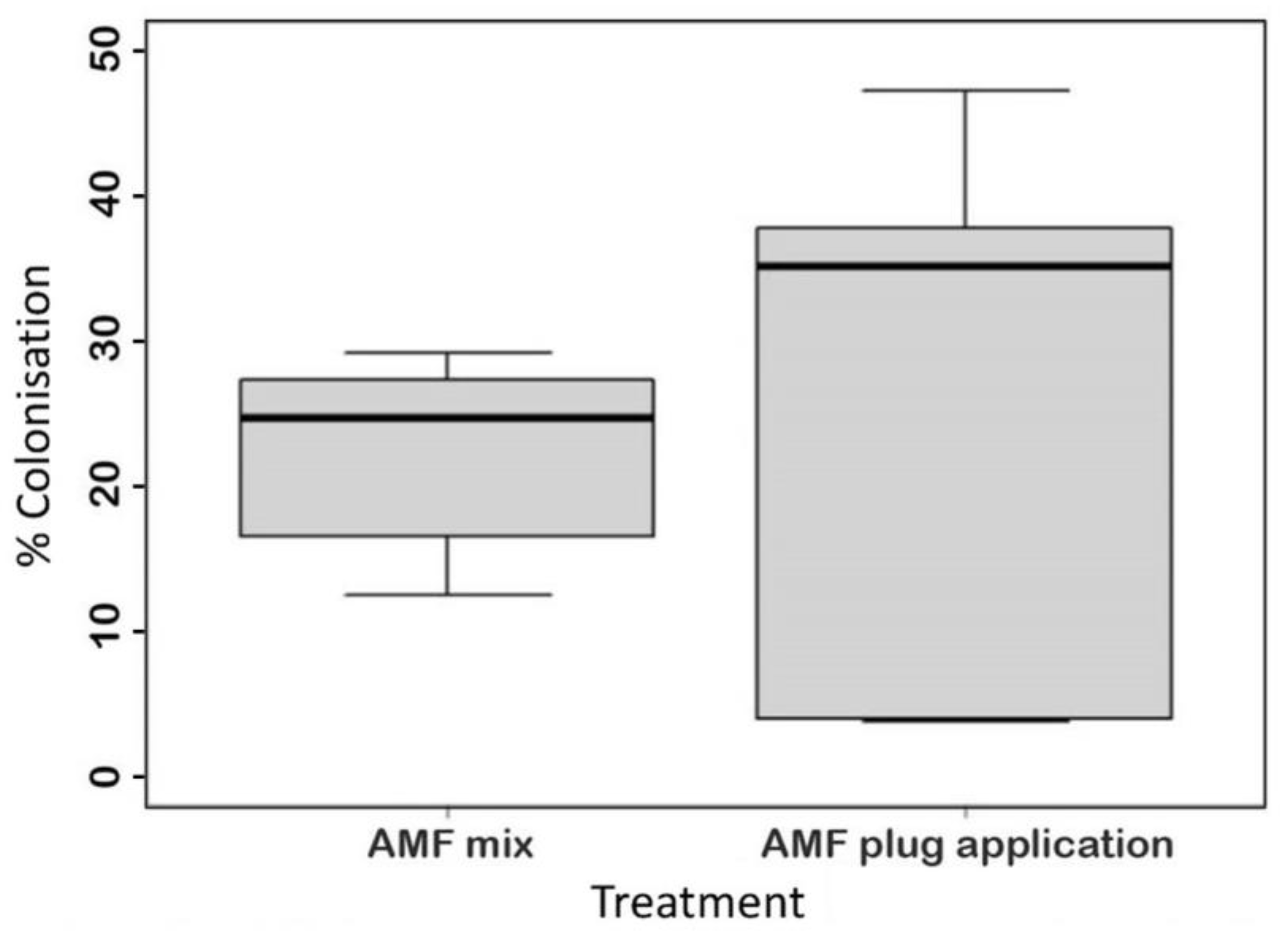
4.1. Root Colonisation Patterns Differed between the Two AMF Application Methods
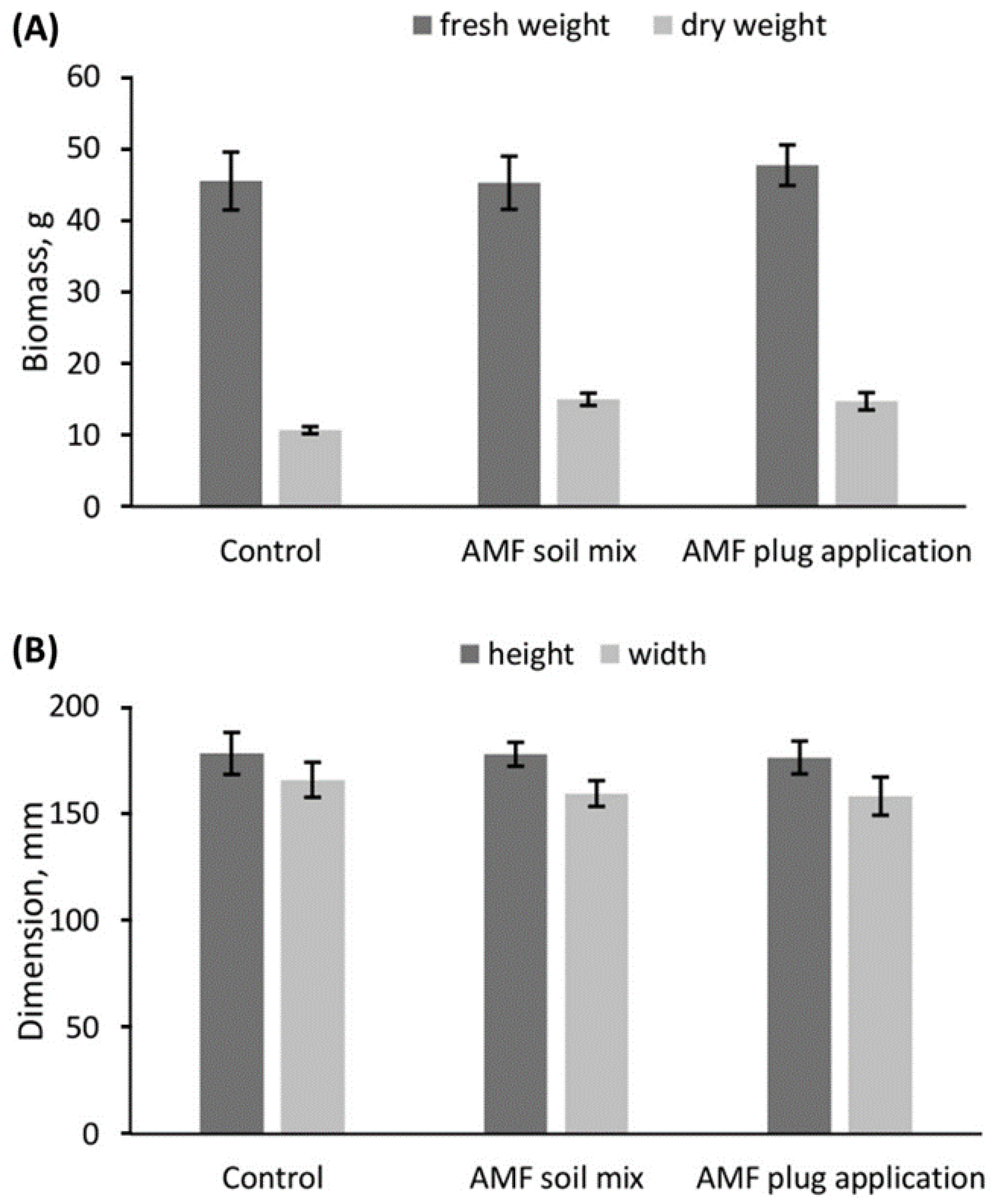
4.2. AMF Application Enhanced Specialised Metabolism but Did Not Change Biomass or Morphology
4.3. AMF Enhances Gene Expression of Terpene Synthases in Rosemary Cultivars
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- TechNavio. Global Greenhouse Horticulture Market 2021–2025; TechNavio (Infiniti Research Ltd.): London, UK, 2021. [Google Scholar]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2018, 249, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K.; Sasaki, K.; Tsurumaru, Y. Prenylation of aromatic compounds, a key diversification of plant secondary metabolites. Phytochemistry 2009, 70, 1739–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazouki, L.; Niinemets, U. Multi-Substrate Terpene Synthases: Their Occurrence and Physiological Significance. Front. Plant Sci. 2016, 7, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özcan, M.M.; Chalchat, J.-C. Chemical composition and antifungal activity of rosemary (Rosmarinus officinalis L.) oil from Turkey. Int. J. Food Sci. Nutr. 2008, 59, 691–698. [Google Scholar] [CrossRef]
- Bornowski, N.; Hamilton, J.P.; Liao, P.; Wood, J.C.; Dudareva, N.; Buell, C.R. Genome sequencing of four culinary herbs reveals terpenoid genes underlying chemodiversity in the Nepetoideae. DNA Res. 2020, 27, dsaa016. [Google Scholar] [CrossRef]
- Tarraf, W.; Ruta, C.; De Cillis, F.; Tagarelli, A.; Tedone, L.; De Mastro, G. Effects of mycorrhiza on growth and essential oil production in selected aromatic plants. Ital. J. Agron. 2015, 10, 160–162. [Google Scholar] [CrossRef] [Green Version]
- Shenata, A.; Nosir, W.; Ahmed, A. Using some biofertilizers treatments to promote growth and oil yield of rosemary plant (Rosmarinus officinalis L.) grown in sandy calcareous soil. Future J. Biol. 2019, 3, 26–33. [Google Scholar]
- Begum, N.; Akhtar, K.; Ahanger, M.A.; Iqbal, M.; Wang, P.; Mustafa, N.S.; Zhang, L. Arbuscular mycorrhizal fungi improve growth, essential oil, secondary metabolism, and yield of tobacco (Nicotiana tabacum L.) under drought stress conditions. Environ. Sci. Pollut. Res. 2021, 28, 45276–45295. [Google Scholar] [CrossRef]
- Saia, S.; Corrado, G.; Vitaglione, P.; Colla, G.; Bonini, P.; Giordano, M.; Stasio, E.; Raimondi, G.; Sacchi, R.; Rouphael, Y. An Endophytic Fungi-Based Biostimulant Modulates Volatile and Non-Volatile Secondary Metabolites and Yield of Greenhouse Basil (Ocimum basilicum L.) through Variable Mechanisms Dependent on Salinity Stress Level. Pathogens 2021, 10, 797. [Google Scholar] [CrossRef]
- Bustamante, M.; Michelozzi, M.; Caracciolo, A.B.; Grenni, P.; Verbokkem, J.; Geerdink, P.; Safi, C.; Nogues, I. Effects of Soil Fertilization on Terpenoids and Other Carbon-Based Secondary Metabolites in Rosmarinus officinalis Plants: A Comparative Study. Plants 2020, 9, 830. [Google Scholar] [CrossRef]
- Seró, R.; Núñez, N.; Núñez, O.; Camprubí, A.; Grases, J.M.; Saurina, J.; Moyano, E.; Calvet, C. Modified distribution in the polyphenolic profile of rosemary leaves induced by plant inoculation with an arbuscular mycorrhizal fungus. J. Sci. Food Agric. 2018, 99, 2966–2973. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- McGONIGLE, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- MacKenzie, D.J.; McLean, M.A.; Mukerji, S.; Green, M. Improved RNA Extraction from Woody Plants for the Detection of Viral Pathogens by Reverse Transcription-Polymerase Chain Reaction. Plant Dis. 1997, 81, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2 (-delta delta CT) method for quantitative re-al-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio PBC: Boston, MA, USA, 2021. [Google Scholar]
- Scagel, C.; Reddy, K.; Armstrong, J. Mycorrhizal Fungi in Rooting Substrate Influences the Quantity and Quality of Roots on Stem Cuttings of Hick’s Yew. Horttechnology 2003, 13, 62–66. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- The Sandy Mush Herb Nursery. The Sandy Mush Herb Nursery Handbook, 9th ed.; Sandy Mush Herb Nursery: Leicester, NC, USA, 2004. [Google Scholar]
- Blerot, B.; Martinelli, L.; Prunier, C.; Saint-Marcoux, D.; Legrand, S.; Bony, A.; Sarrabère, L.; Gros, F.; Boyer, N.; Caissard, J.-C.; et al. Functional Analysis of Four Terpene Synthases in Rose-Scented Pelargonium Cultivars (Pelargonium × hybridum) and Evolution of Scent in the Pelargonium Genus. Front. Plant Sci. 2018, 9, 1435. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-Q.; Fu, N.-N.; Zhou, J.-J.; Wang, G.-R. Functional characterization of (E)-β-caryophyllene synthase from lima bean and its up-regulation by spider mites and alamethicin. J. Integr. Agric. 2017, 16, 2231–2238. [Google Scholar] [CrossRef]
- Duc, N.H.; Vo, A.T.; Haddidi, I.; Daood, H.; Posta, K. Arbuscular Mycorrhizal Fungi Improve Tolerance of the Medicinal Plant Eclipta prostrata (L.) and Induce Major Changes in Polyphenol Profiles Under Salt Stresses. Front. Plant Sci. 2021, 11, 612299. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Upadhyay, S.; Wajid, S.; Ram, M.; Jain, D.C.; Singh, V.P.; Abdin, M.Z.; Kapoor, R. Arbuscular mycorrhiza increase artemisinin accumulation in Artemisia annua by higher expression of key biosynthesis genes via enhanced jasmonic acid levels. Mycorrhiza 2014, 25, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M. Sites of synthesis, biochemistry and functional role of plant volatiles. South Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef] [Green Version]
- Kobae, Y. Dynamic Phosphate Uptake in Arbuscular Mycorrhizal Roots Under Field Conditions. Front. Environ. Sci. 2019, 6, 159. [Google Scholar] [CrossRef] [Green Version]
- Peçanha, D.A.; Freitas, M.S.M.; Vieira, M.E.; Cunha, J.M.; de Jesus, A.C. Phosphorus fertilization affects growth, essential oil yield and quality of true lavender in Brazil. Ind. Crop. Prod. 2021, 170, 113803. [Google Scholar] [CrossRef]
- Motamedi, M.; Zahedi, M.; Karimmojeni, H.; Motamedi, H.; Mastinu, A. Effect of rhizosphere bacteria on antioxidant enzymes and some biochemical characteristics of Medicago sativa L. subjected to herbicide stress. Acta Physiol. Plant. 2022, 44, 1–12. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Arabidopsis Ortholog | Forward Primer | Reverse Primer |
|---|---|---|---|
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH | AAGCATCGGAGACCAAGCTC | CGCGAGAACTGTAACCCCAT |
| Ocimene synthase | TPS03 | GGTACCACACGGGGCATAAA | CAAGATCATCTGCAAGCCGC |
| β-caryophyllene synthase | TPS12 | AGACTGGCCGTAGCAAACTC | CCGATTGTTCAGGCAACACG |
| Cineole synthase | TPS27 | CAGGCATCCTTGCCACATGA | GCCAAACGTTGAGAAAGCCC |
| Linalool synthase | TPS14 | GCCAAATTCAGAGAGGCCCTT | TTGTCCGAGAAGGAAGCACG |
| Myrcene synthase | TPS24 | TGACGCGAACCCTATTCTGG | CAAACCCCAACTTTTCCGGC |
| Lupeol synthase | LUP2 | CTGGCTCTTCCCTTCCGTTT | TAAAACGACGTCGGTGAGGG |
| Terpene synthase 07 | TPS07 | CGATGTTCGTGTTCTTGCCC | CCTTCAAATCTCCTCCCCCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leggatt, E.; Griffiths, A.; Budge, S.; Stead, A.D.; Gange, A.C.; Devlin, P.F. Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars. Life 2023, 13, 315. https://doi.org/10.3390/life13020315
Leggatt E, Griffiths A, Budge S, Stead AD, Gange AC, Devlin PF. Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars. Life. 2023; 13(2):315. https://doi.org/10.3390/life13020315
Chicago/Turabian StyleLeggatt, Emily, Alistair Griffiths, Simon Budge, Anthony D. Stead, Alan C. Gange, and Paul F. Devlin. 2023. "Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars" Life 13, no. 2: 315. https://doi.org/10.3390/life13020315
APA StyleLeggatt, E., Griffiths, A., Budge, S., Stead, A. D., Gange, A. C., & Devlin, P. F. (2023). Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars. Life, 13(2), 315. https://doi.org/10.3390/life13020315