
Low-Level Laser Therapy Induces Melanoma Tumor Growth by Promoting Angiogenesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model and Cell Culture
2.2. Laser Irradiation
2.3. Cell Proliferation Assay
2.4. Tissue Collection
2.5. Immunohistochemical Analysis
2.6. Immunofluorescence Analysis
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
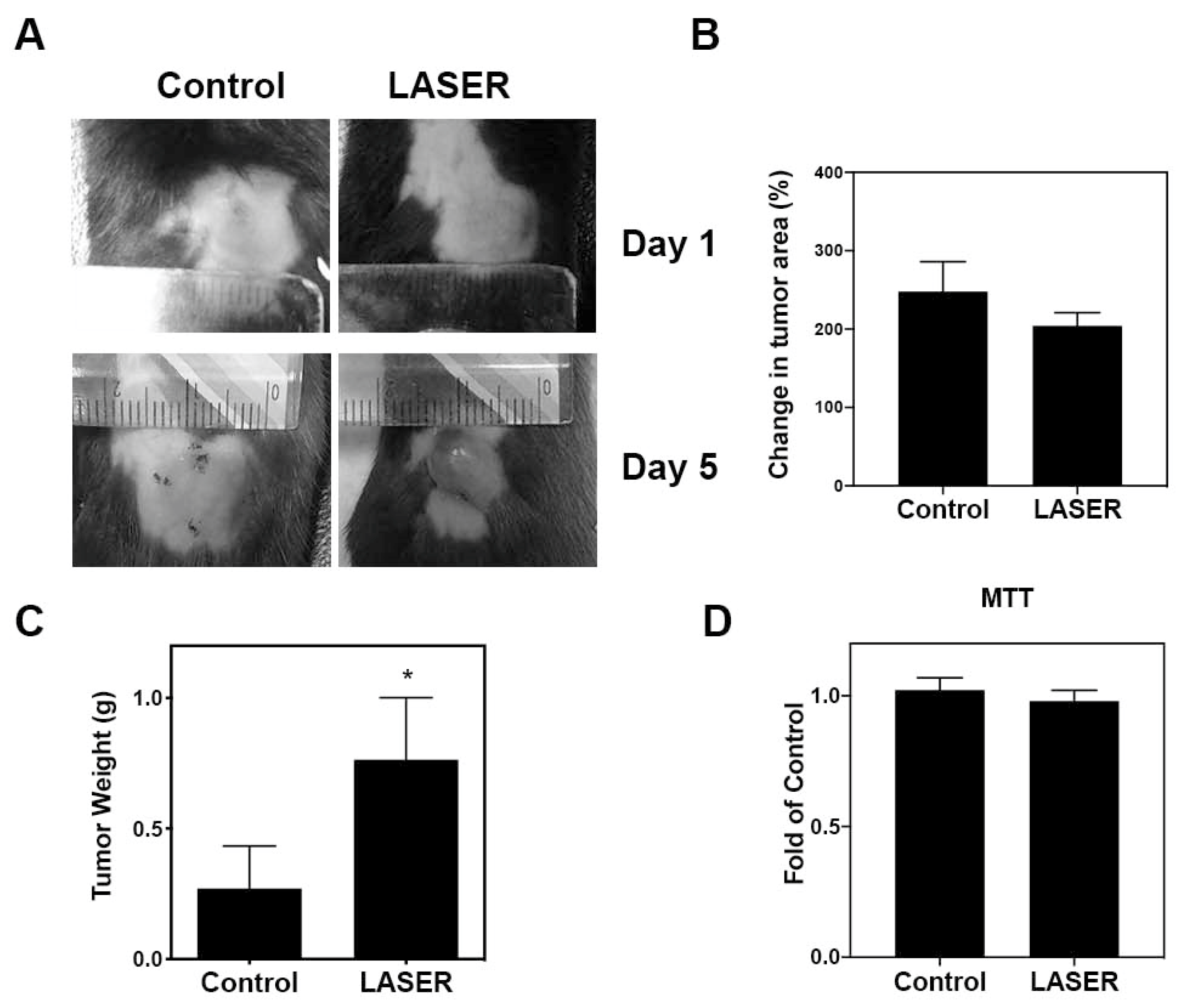
3.1. LLLT Enhanced Tumor Progression In Vivo, but Not Cell Proliferation In Vitro
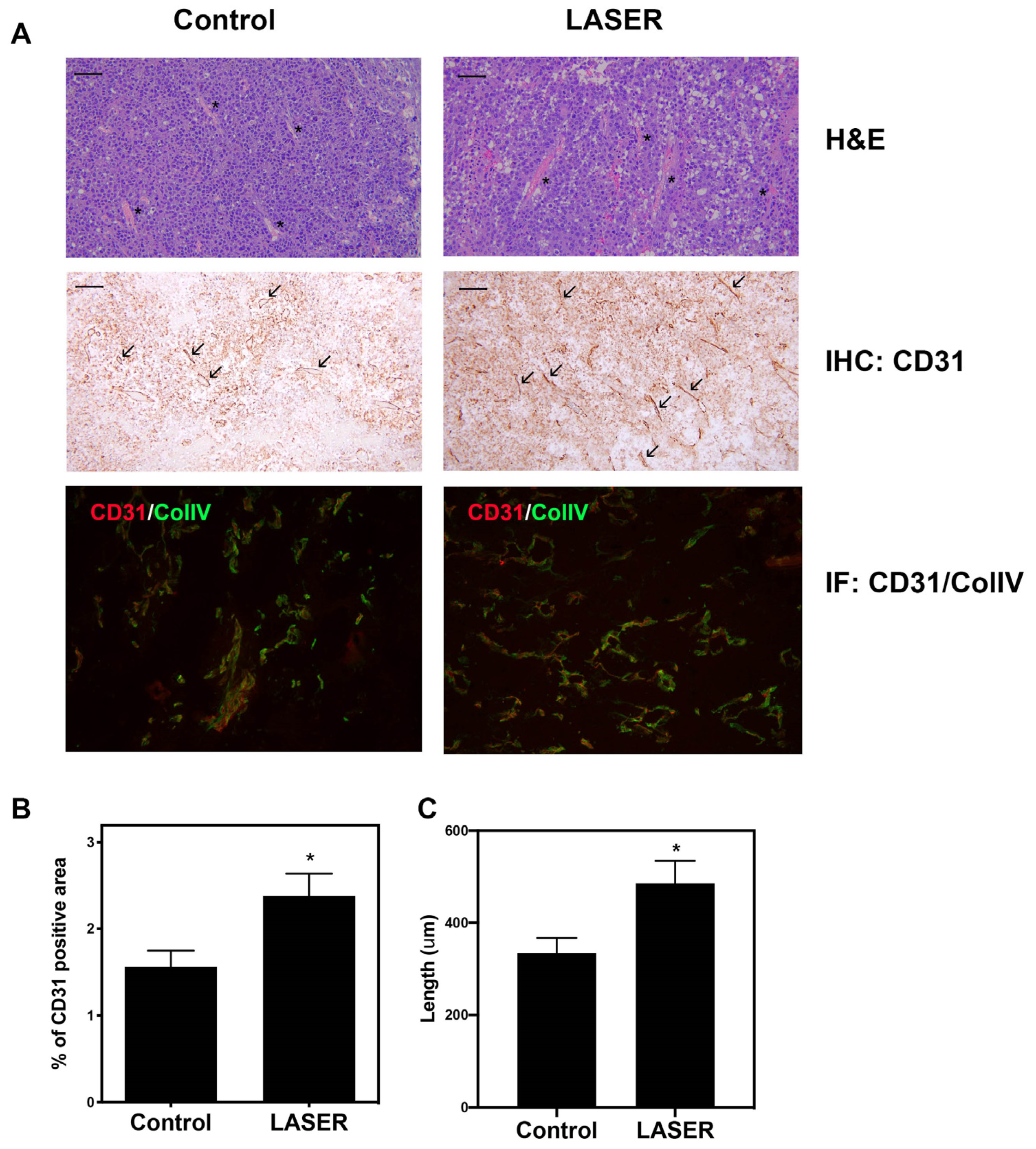
3.2. LLLT Promoted Angiogenesis and Expanded CD31-Positive Vascular Area in Tumors
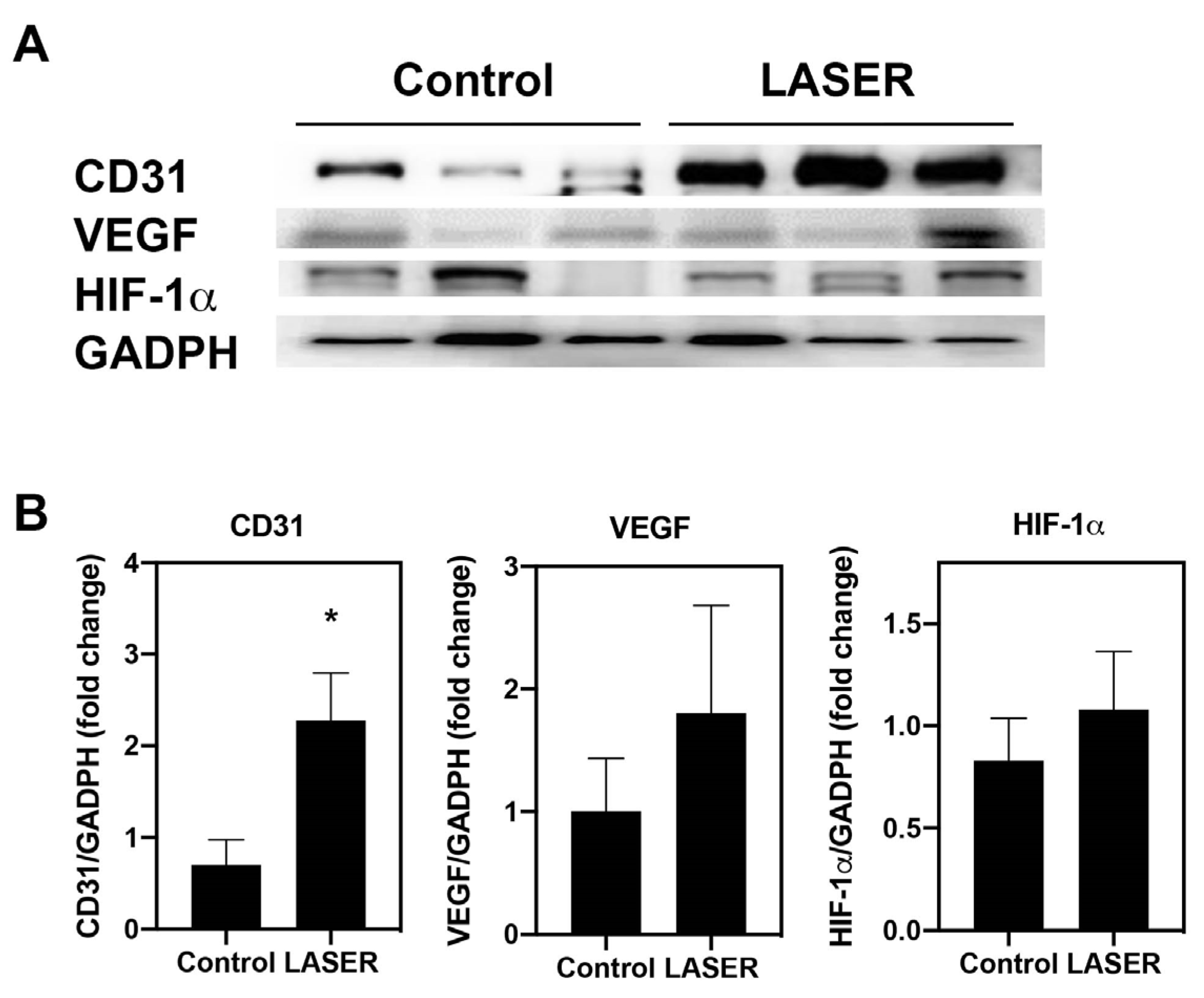
3.3. Angiogenesis-Related Molecules in Tumors
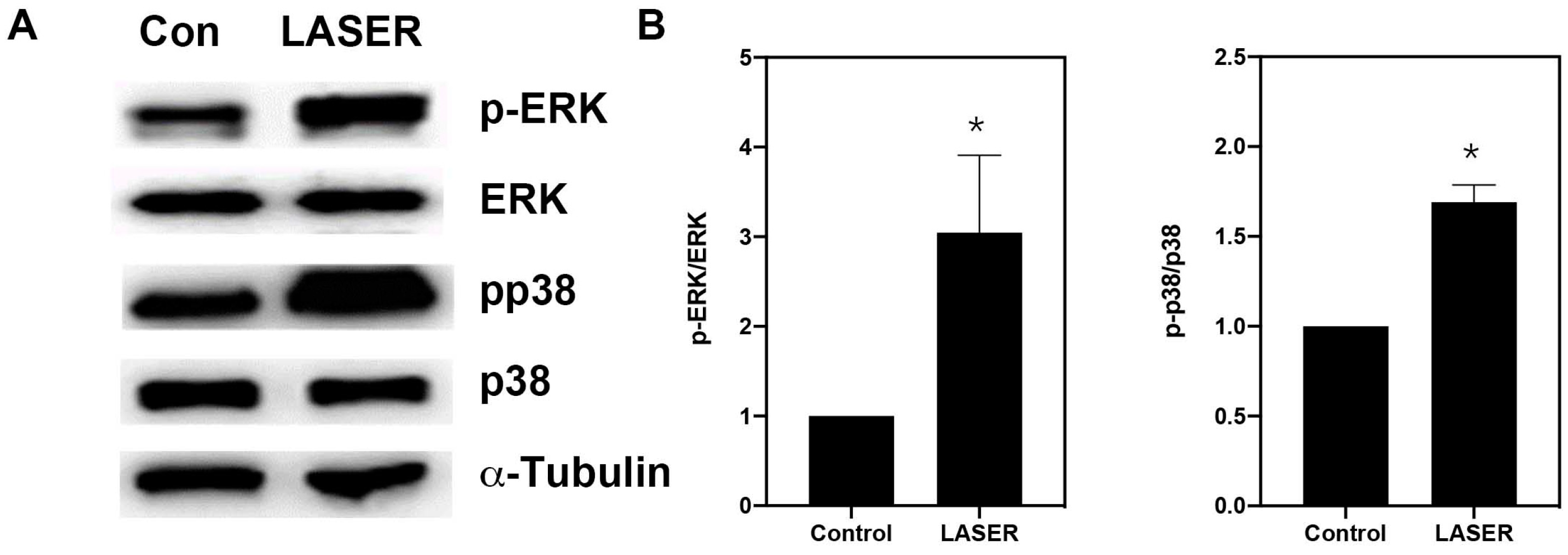
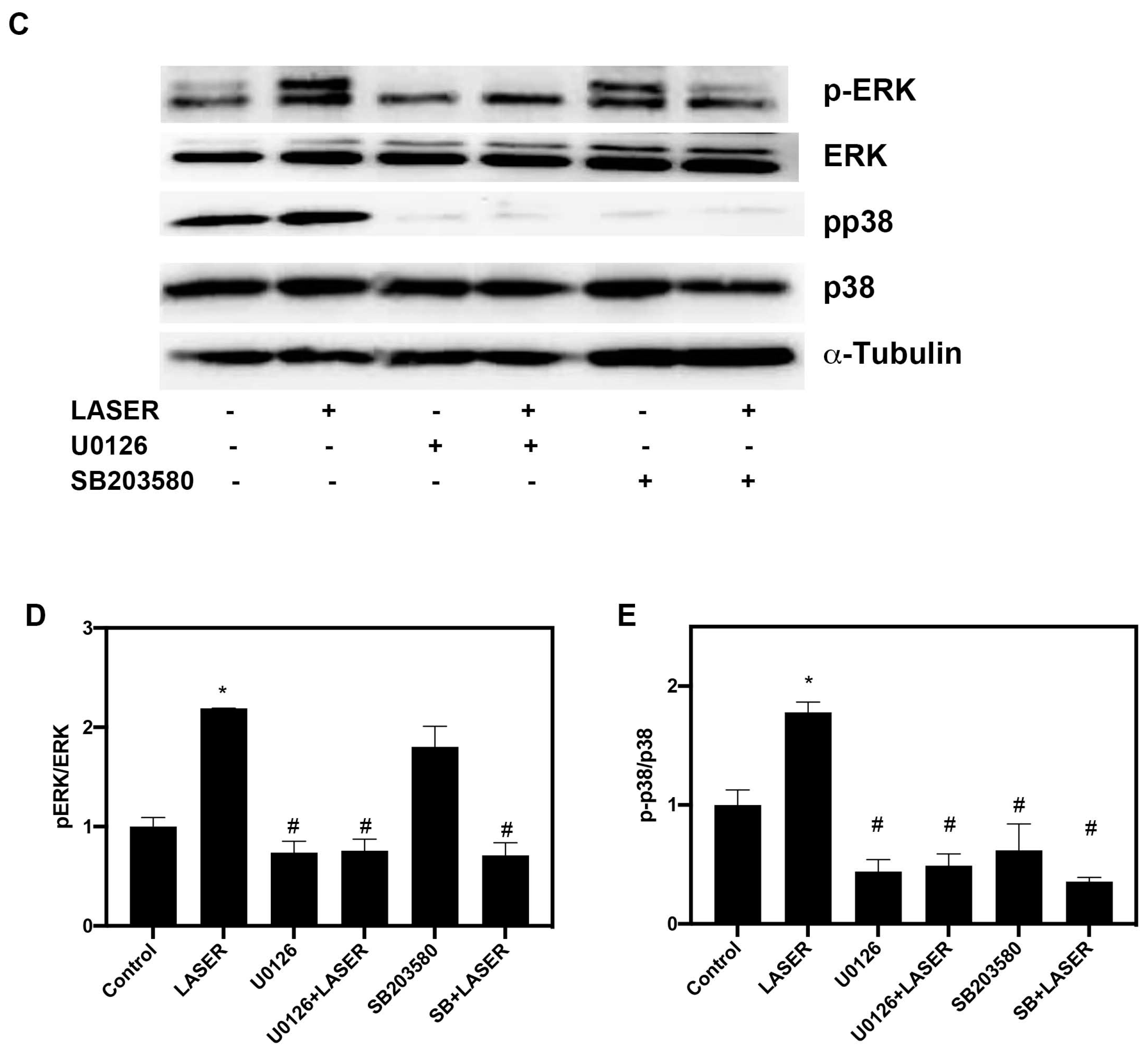
3.4. LLLT Induced the Phosphorylation of ERK and p38 MAPK in B16F10 Cells
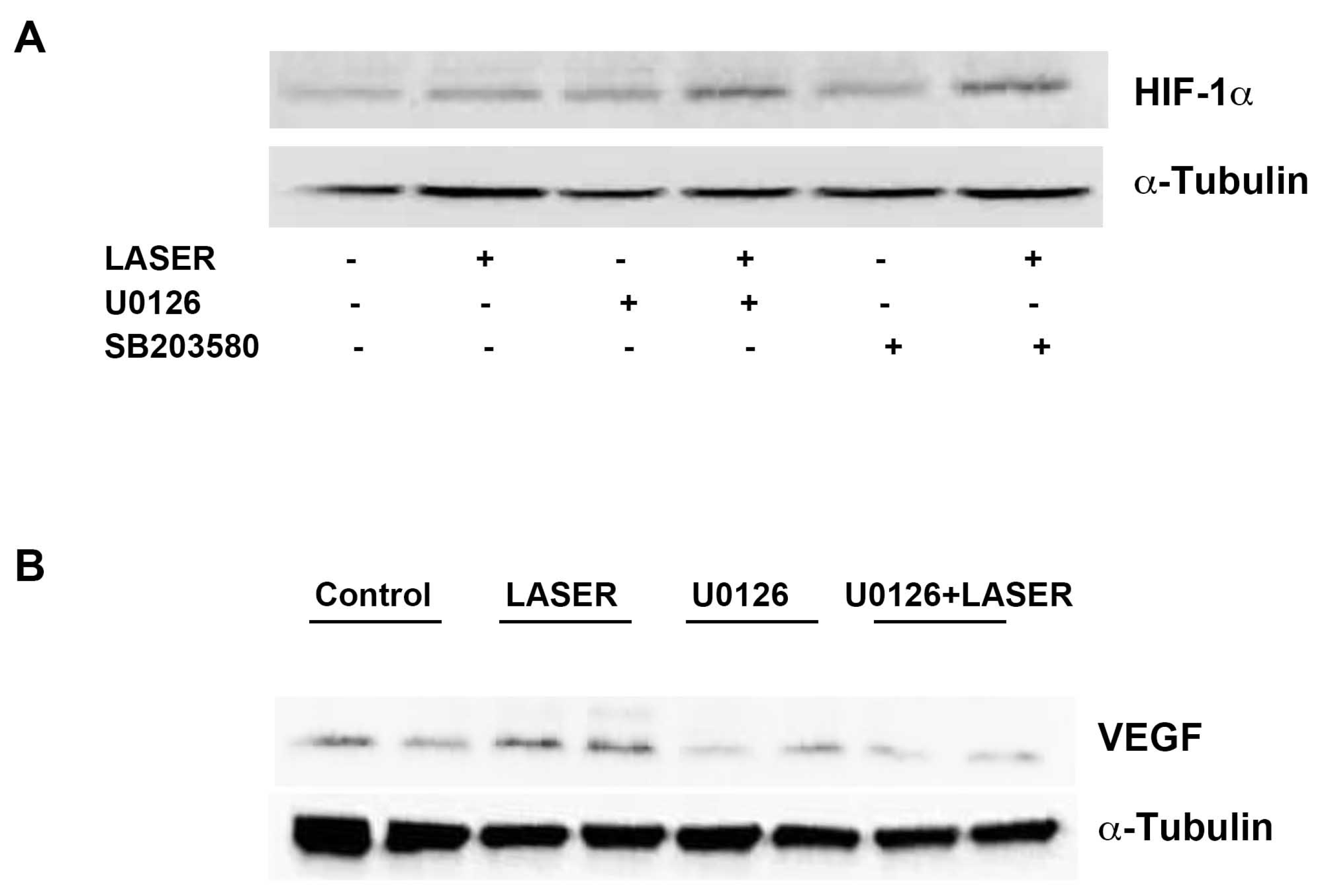
3.5. LLLT Induced the Expression of VEGF, but Not HIF-1α, through ERK Signaling
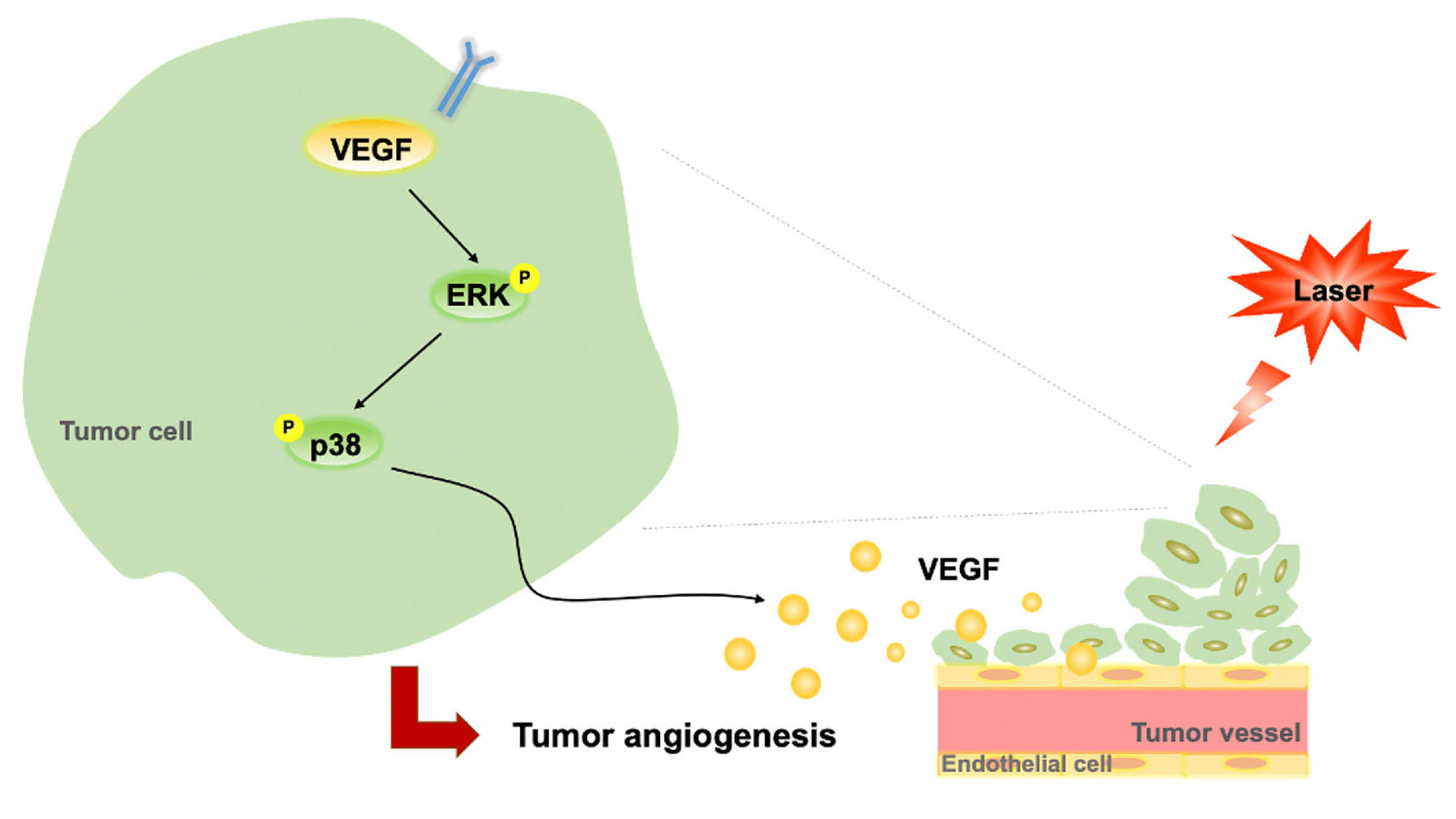
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taradaj, J.; Shay, B.; Dymarek, R.; Sopel, M.; Walewicz, K.; Beeckman, D.; Schoonhoven, L.; Gefen, A.; Rosinczuk, J. Effect of laser therapy on expression of angio- and fibrogenic factors, and cytokine concentrations during the healing process of human pressure ulcers. Int. J. Med. Sci. 2018, 15, 1105–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruh, A.C.; Frigo, L.; Cavalcanti, M.; Svidnicki, P.; Vicari, V.N.; Lopes-Martins, R.A.B.; Leal Junior, E.C.P.; De Isla, N.; Diomede, F.; Trubiani, O.; et al. Laser photobiomodulation in pressure ulcer healing of human diabetic patients: Gene expression analysis of inflammatory biochemical markers. Lasers Med. Sci. 2018, 33, 165–171. [Google Scholar] [CrossRef]
- Taradaj, J.; Rajfur, K.; Shay, B.; Rajfur, J.; Ptaszkowski, K.; Walewicz, K.; Dymarek, R.; Sopel, M.; Rosinczuk, J. Photobiomodulation using high- or low-level laser irradiations in patients with lumbar disc degenerative changes: Disappointing outcomes and remarks. Clin. Interv. Aging 2018, 13, 1445–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avci, P.; Gupta, A.; Sadasivam, M.; Vecchio, D.; Pam, Z.; Pam, N.; Hamblin, M.R. Low-level laser (light) therapy (LLLT) in skin: Stimulating, healing, restoring. Semin. Cutan. Med. Surg. 2013, 32, 41–52. [Google Scholar] [PubMed]
- Lim, W.; Lee, S.; Kim, I.; Chung, M.; Kim, M.; Lim, H.; Park, J.; Kim, O.; Choi, H. The anti-inflammatory mechanism of 635 nm light-emitting-diode irradiation compared with existing COX inhibitors. Lasers Surg. Med. 2007, 39, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Bjordal, J.M.; Johnson, M.I.; Iversen, V.; Aimbire, F.; Lopes-Martins, R.A. Low-level laser therapy in acute pain: A systematic review of possible mechanisms of action and clinical effects in randomized placebo-controlled trials. Photomed. Laser Surg. 2006, 24, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Enwemeka, C.S.; Parker, J.C.; Dowdy, D.S.; Harkness, E.E.; Sanford, L.E.; Woodruff, L.D. The efficacy of low-power lasers in tissue repair and pain control: A meta-analysis study. Photomed. Laser Surg. 2004, 22, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, J.T.; McLoda, T.A.; Seegmiller, J.G.; David Baxter, G. Low-Level Laser Therapy Facilitates Superficial Wound Healing in Humans: A Triple-Blind, Sham-Controlled Study. J. Athl. Train. 2004, 39, 223–229. [Google Scholar]
- Sroka, R.; Schaffer, M.; Fuchs, C.; Pongratz, T.; Schrader-Reichard, U.; Busch, M.; Schaffer, P.M.; Duhmke, E.; Baumgartner, R. Effects on the mitosis of normal and tumor cells induced by light treatment of different wavelengths. Lasers Surg. Med. 1999, 25, 263–271. [Google Scholar] [CrossRef]
- Dastanpour, S.; Momen Beitollahi, J.; Saber, K. The effect of low-level laser therapy on human leukemic cells. J. Lasers Med. Sci. 2015, 6, 74–79. [Google Scholar]
- Tian, Y.; Lee, Y.; Kim, H.; Kang, H.W. In vitro anti-tumor effect of low-power laser irradiation (LPLI) on gastroenterological carcinoma cells. Lasers Med. Sci. 2020, 35, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, M.; Sroka, R.; Fuchs, C.; Schrader-Reichardt, U.; Schaffer, P.M.; Busch, M.; Duhmke, E. Biomodulative effects induced by 805 nm laser light irradiation of normal and tumor cells. J. Photochem. Photobiol. B 1997, 40, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Frigo, L.; Luppi, J.S.; Favero, G.M.; Maria, D.A.; Penna, S.C.; Bjordal, J.M.; Bensadoun, R.J.; Lopes-Martins, R.A. The effect of low-level laser irradiation (In-Ga-Al-AsP—660 nm) on melanoma in vitro and in vivo. BMC Cancer 2009, 9, 404. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.H.; Moon, J.H.; Choi, S.H.; Ahn, J.C. Low-Level Laser Therapy Promoted Aggressive Proliferation and Angiogenesis Through Decreasing of Transforming Growth Factor-beta1 and Increasing of Akt/Hypoxia Inducible Factor-1alpha in Anaplastic Thyroid Cancer. Photomed. Laser Surg. 2016, 34, 229–235. [Google Scholar] [CrossRef]
- Ottaviani, G.; Martinelli, V.; Rupel, K.; Caronni, N.; Naseem, A.; Zandona, L.; Perinetti, G.; Gobbo, M.; Di Lenarda, R.; Bussani, R.; et al. Laser Therapy Inhibits Tumor Growth in Mice by Promoting Immune Surveillance and Vessel Normalization. EBioMedicine 2016, 11, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Austin, E.; Huang, A.; Wang, J.Y.; Cohen, M.; Heilman, E.; Maverakis, E.; Michl, J.; Jagdeo, J. Red Light Phototherapy Using Light-Emitting Diodes Inhibits Melanoma Proliferation and Alters Tumor Microenvironments. Front. Oncol. 2022, 12, 928484. [Google Scholar] [CrossRef]
- Antunes, H.S.; Herchenhorn, D.; Small, I.A.; Araujo, C.M.M.; Viegas, C.M.P.; de Assis Ramos, G.; Dias, F.L.; Ferreira, C.G. Long-term survival of a randomized phase III trial of head and neck cancer patients receiving concurrent chemoradiation therapy with or without low-level laser therapy (LLLT) to prevent oral mucositis. Oral Oncol. 2017, 71, 11–15. [Google Scholar] [CrossRef]
- Karu, T.I.; Pyatibrat, L.V.; Kolyakov, S.F.; Afanasyeva, N.I. Absorption measurements of a cell monolayer relevant to phototherapy: Reduction of cytochrome c oxidase under near IR radiation. J. Photochem. Photobiol. B 2005, 81, 98–106. [Google Scholar] [CrossRef]
- Tafur, J.; Mills, P.J. Low-intensity light therapy: Exploring the role of redox mechanisms. Photomed. Laser Surg. 2008, 26, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.H.; Chen, S.Y.; Hsieh, Y.L.; Teng, Y.H.; Cheng, Y.J. Low-level laser therapy prevents endothelial cells from TNF-alpha/cycloheximide-induced apoptosis. Lasers Med. Sci. 2018, 33, 279–286. [Google Scholar] [CrossRef]
- de Loura Santana, C.; Silva Dde, F.; Deana, A.M.; Prates, R.A.; Souza, A.P.; Gomes, M.T.; de Azevedo Sampaio, B.P.; Shibuya, J.F.; Bussadori, S.K.; Mesquita-Ferrari, R.A.; et al. Tissue responses to postoperative laser therapy in diabetic rats submitted to excisional wounds. PLoS ONE 2015, 10, e0122042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loevschall, H.; Arenholt-Bindslev, D. Effect of low level diode laser irradiation of human oral mucosa fibroblasts in vitro. Lasers Surg. Med. 1994, 14, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Lapchak, P.A.; De Taboada, L. Transcranial near infrared laser treatment (NILT) increases cortical adenosine-5′-triphosphate (ATP) content following embolic strokes in rabbits. Brain Res. 2010, 1306, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Goralczyk, K.; Szymanska, J.; Lukowicz, M.; Drela, E.; Kotzbach, R.; Dubiel, M.; Michalska, M.; Goralczyk, B.; Zajac, A.; Rosc, D. Effect of LLLT on endothelial cells culture. Lasers Med. Sci. 2015, 30, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Cury, V.; Moretti, A.I.; Assis, L.; Bossini, P.; Crusca Jde, S.; Neto, C.B.; Fangel, R.; de Souza, H.P.; Hamblin, M.R.; Parizotto, N.A. Low level laser therapy increases angiogenesis in a model of ischemic skin flap in rats mediated by VEGF, HIF-1alpha and MMP-2. J. Photochem. Photobiol. B 2013, 125, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folkman, J. Seminars in Medicine of the Beth Israel Hospital, Boston. Clinical applications of research on angiogenesis. N. Engl. J. Med. 1995, 333, 1757–1763. [Google Scholar] [CrossRef]
- Hicklin, D.J.; Ellis, L.M. Role of the vascular endothelial growth factor pathway in tumor growth and angiogenesis. J. Clin. Oncol. 2005, 23, 1011–1027. [Google Scholar] [CrossRef]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1alpha and HIF2alpha: Sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 2011, 12, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Burke, B.; Tang, N.; Corke, K.P.; Tazzyman, D.; Ameri, K.; Wells, M.; Lewis, C.E. Expression of HIF-1alpha by human macrophages: Implications for the use of macrophages in hypoxia-regulated cancer gene therapy. J. Pathol. 2002, 196, 204–212. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Pastushenko, I.; Van den Eynden, G.G.; Vicente-Arregui, S.; Prieto-Torres, L.; Alvarez-Alegret, R.; Querol, I.; Dirix, L.Y.; Carapeto, F.J.; Vermeulen, P.B.; Van Laere, S.J. Increased Angiogenesis and Lymphangiogenesis in Metastatic Sentinel Lymph Nodes Is Associated With Nonsentinel Lymph Node Involvement and Distant Metastasis in Patients with Melanoma. Am. J. Dermatopathol. 2016, 38, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Streit, M.; Detmar, M. Angiogenesis, lymphangiogenesis, and melanoma metastasis. Oncogene 2003, 22, 3172–3179. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Lee, S.Y.; Teng, Y.H.; Lee, S.D.; Cheng, Y.J. Photobiomodulation Therapy to Promote Angiogenesis in Diabetic Mice with Hindlimb Ischemia. Photobiomodul. Photomed. Laser Surg. 2021, 39, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y.; Lin, J.S.; Lin, H.C.; Shan, Y.S.; Cheng, Y.J.; Yang, B.C. Dependence of fibroblast infiltration in tumor stroma on type IV collagen-initiated integrin signal through induction of platelet-derived growth factor. Biochim. Biophys. Acta 2015, 1853, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.Y.; Chen, Y.J.; Lu, Y.C.; Liao, C.T.; Chen, I.H.; Chang, J.T.; Huang, Y.C.; Chen, W.H.; Huang, C.C.; Tsai, C.Y.; et al. Fascin is a circulating tumor marker for head and neck cancer as determined by a proteomic analysis of interstitial fluid from the tumor microenvironment. Clin. Chem. Lab. Med. 2015, 53, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R.; Nelson, S.T.; Strahan, J.R. Photobiomodulation and Cancer: What Is the Truth? Photomed. Laser Surg. 2018, 36, 241–245. [Google Scholar] [CrossRef]
- Fidler, I.J.; Nicolson, G.L. Fate of recirculating B16 melanoma metastatic variant cells in parabiotic syngeneic recipients. J. Natl. Cancer Inst. 1977, 58, 1867–1872. [Google Scholar] [CrossRef]
- Eddy, K.; Chen, S. Overcoming Immune Evasion in Melanoma. Int. J. Mol. Sci. 2020, 21, 8984. [Google Scholar] [CrossRef]
- das Neves, L.M.; Leite, G.P.; Marcolino, A.M.; Pinfildi, C.E.; Garcia, S.B.; de Araujo, J.E.; Guirro, E.C. Laser photobiomodulation (830 and 660 nm) in mast cells, VEGF, FGF, and CD34 of the musculocutaneous flap in rats submitted to nicotine. Lasers Med. Sci. 2017, 32, 335–341. [Google Scholar] [CrossRef]
- Zhang, W.G.; Wu, C.Y.; Pan, W.X.; Tian, L.; Xia, J.L. Low-power Helium-Neon laser irradiation enhances the expression of VEGF in murine myocardium. Chin. Med. J. 2004, 117, 1476–1480. [Google Scholar]
- Kawano, Y.; Utsunomiya-Kai, Y.; Kai, K.; Miyakawa, I.; Ohshiro, T.; Narahara, H. The production of VEGF involving MAP kinase activation by low level laser therapy in human granulosa cells. Laser Ther. 2012, 21, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.W.; Mayo, L.D.; Dunbar, J.D.; Kessler, K.M.; Baerwald, M.R.; Jaffe, E.A.; Wang, D.; Warren, R.S.; Donner, D.B. Utilization of distinct signaling pathways by receptors for vascular endothelial cell growth factor and other mitogens in the induction of endothelial cell proliferation. J. Biol. Chem. 2000, 275, 5096–5103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berra, E.; Milanini, J.; Richard, D.E.; Le Gall, M.; Vinals, F.; Gothie, E.; Roux, D.; Pages, G.; Pouyssegur, J. Signaling angiogenesis via p42/p44 MAP kinase and hypoxia. Biochem. Pharmacol. 2000, 60, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Mylonis, I.; Chachami, G.; Samiotaki, M.; Panayotou, G.; Paraskeva, E.; Kalousi, A.; Georgatsou, E.; Bonanou, S.; Simos, G. Identification of MAPK phosphorylation sites and their role in the localization and activity of hypoxia-inducible factor-1alpha. J. Biol. Chem. 2006, 281, 33095–33106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almalki, S.G.; Agrawal, D.K. ERK signaling is required for VEGF-A/VEGFR2-induced differentiation of porcine adipose-derived mesenchymal stem cells into endothelial cells. Stem Cell Res. Ther. 2017, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Schluter, A.; Weller, P.; Kanaan, O.; Nel, I.; Heusgen, L.; Hoing, B.; Hasskamp, P.; Zander, S.; Mandapathil, M.; Dominas, N.; et al. CD31 and VEGF are prognostic biomarkers in early-stage, but not in late-stage, laryngeal squamous cell carcinoma. BMC Cancer 2018, 18, 272. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Kang, G.; Wang, T.; Huang, H. Tumor angiogenesis and anti-angiogenic gene therapy for cancer. Oncol. Lett. 2018, 16, 687–702. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Sorenson, C.M.; Sheibani, N. PECAM-1 isoforms, eNOS and endoglin axis in regulation of angiogenesis. Clin. Sci. 2015, 129, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Zhou, F.; Wu, S.; Liu, L.; Xing, D. Phototherapy-Induced Antitumor Immunity: Long-Term Tumor Suppression Effects via Photoinactivation of Respiratory Chain Oxidase-Triggered Superoxide Anion Burst. Antioxid. Redox Signal. 2016, 24, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zhou, F.; Wei, Y.; Chen, W.R.; Chen, Q.; Xing, D. Cancer phototherapy via selective photoinactivation of respiratory chain oxidase to trigger a fatal superoxide anion burst. Antioxid. Redox Signal. 2014, 20, 733–746. [Google Scholar] [CrossRef] [Green Version]
- de Castro, J.L.; Pinheiro, A.L.; Werneck, C.E.; Soares, C.P. The effect of laser therapy on the proliferation of oral KB carcinoma cells: An in vitro study. Photomed. Laser Surg. 2005, 23, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Langenkamp, E.; Vom Hagen, F.M.; Zwiers, P.J.; Moorlag, H.E.; Schouten, J.P.; Hammes, H.P.; Gouw, A.S.; Molema, G. Tumor Vascular Morphology Undergoes Dramatic Changes during Outgrowth of B16 Melanoma While Proangiogenic Gene Expression Remains Unchanged. ISRN Oncol. 2011, 2011, 409308. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ERK | Cell Signaling #9102 |
| p-ERK | Cell Signaling #9101 |
| P38 MAPK | Cell Signaling #9212 |
| p-p38 MAPK | Cell Signaling #9211 |
| CD31 | Invitrogen #PA5-16301 |
| HIF-1α | Invitrogen # MA1-516 |
| β-actin | GeneTex #GTX109639 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-Y.; Lee, S.-Y.; Cheng, Y.-J. Low-Level Laser Therapy Induces Melanoma Tumor Growth by Promoting Angiogenesis. Life 2023, 13, 320. https://doi.org/10.3390/life13020320
Lin Y-Y, Lee S-Y, Cheng Y-J. Low-Level Laser Therapy Induces Melanoma Tumor Growth by Promoting Angiogenesis. Life. 2023; 13(2):320. https://doi.org/10.3390/life13020320
Chicago/Turabian StyleLin, Yi-Yuan, Shin-Yi Lee, and Yu-Jung Cheng. 2023. "Low-Level Laser Therapy Induces Melanoma Tumor Growth by Promoting Angiogenesis" Life 13, no. 2: 320. https://doi.org/10.3390/life13020320
APA StyleLin, Y. -Y., Lee, S. -Y., & Cheng, Y. -J. (2023). Low-Level Laser Therapy Induces Melanoma Tumor Growth by Promoting Angiogenesis. Life, 13(2), 320. https://doi.org/10.3390/life13020320