
The Anticancer Potential of Psidium guajava (Guava) Extracts

 , ,
, ,  and
and
Abstract
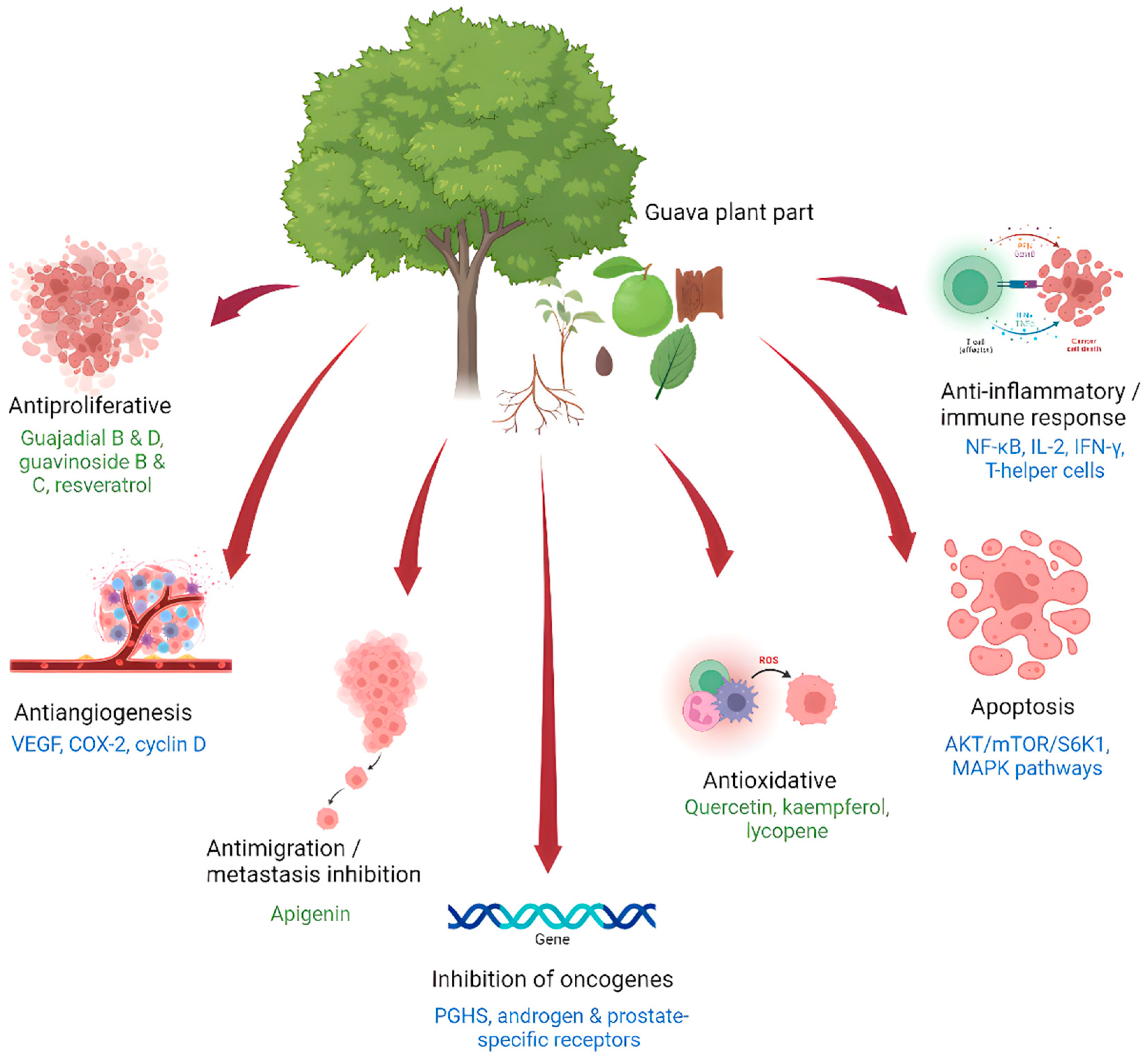
:1. Introduction
2. Anticancer Effects of the Psidium guajava Plant
2.1. Anticancer Effects of Psidium guajava Leaf Extracts
2.2. Anticancer Effects of Psidium guajava Fruit Extracts
2.3. Anticancer Effects of Psidium guajava Seed Extracts
2.4. Anticancer Effects of Psidium guajava Bark and Root Extracts
2.5. In Vivo Anticancer Effects of Psidium guajava Extracts
3. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Morton, J.F. Fruits of Warm Climates; J.F. Morton: Miami, FL, USA, 1987. [Google Scholar]
- Rios, C.; Salazar, C.R.; Cardona, C.; Victoria, K.; Torres, M. Guayaba. In Frutales. Manual de Asistencia Técnica, 2nd ed.; Instituto Colombiano Agropecuario: Bogotá, Colombia, 1977; pp. 4221–4248. [Google Scholar]
- Postman, J.; Hummer, K.; Stover, E.; Krueger, R.; Forsline, P.; Grauke, L.J.; Zee, F.; Ayala-Silva, T.; Irish, B. Fruit and nut genebanks in the US National Plant Germplasm System. HortScience 2006, 41, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Stone, B.C. The Flora of Guam; University of Guam: Mangilao, GU, USA, 1970. [Google Scholar]
- Gutiérrez, R.M.P.; Mitchell, S.; Solis, R.V. Psidium guajava: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Farinazzi-Machado, F.M.; de Alvares Goulart, R.; Brunnati, A.C.S.; Otoboni, A.M.; Ottoboni, B.J.M.A.P. Psidium guajava (Guava): A plant of multipurpose medicinal applications. Med. Aromat. Plants 2012, 1, 1000104. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Cancer Fact Sheet. 2018. Available online: http://www.who.int/en/news-room/fact-sheets/detail/cancer (accessed on 13 August 2018).
- Rizzo, L.; Longato, G.B.; Ruiz, A.L.T.G.; Tinti, S.V.; Possenti, A.; Vendramini-Costa, D.B.; Sartoratto, A.; Figueira, G.M.; Silva, F.L.N.; Eberlin, M.N.; et al. In vitro, in vivo and in silico analysis of the anticancer and estrogen-like activity of guava leaf extracts. Curr. Med. Chem. 2014, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Nakamura, T.; Hosokawa, T.; Suzuki-Yamamoto, T.; Yamashita, H.; Kimoto, M.; Tsuji, H.; Yoshida, H.; Hada, T.; Takahashi, Y. Antiproliferative activity of guava leaf extract via inhibition of prostaglandin endoperoxide H synthase isoforms. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and P388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef]
- Morgan, D.M. Tetrazolium (MTT) assay for cellular viability and activity. In Polyamine Protocols; Springer: New York, NY, USA, 1998; pp. 179–184. [Google Scholar]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Borenfreund, E.; Puerner, J.A. A simple quantitative procedure using monolayer cultures for cytotoxicity assays (HTD/NR-90). Methods Cell Sci. 1985, 9, 7–9. [Google Scholar] [CrossRef]
- Haugland, R. Assays for cell viability, proliferation and function. In The Handbook of Flourescent Probes and Research Chemicals; Molecular Probes: Eugene, OR, USA, 1996; pp. 365–398. [Google Scholar]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 1997, 21, A.3B.1–A.3B.2. [Google Scholar] [CrossRef]
- Qin, X.-J.; Yu, Q.; Yan, H.; Khan, A.; Feng, M.-Y.; Li, P.-P.; Hao, X.-J.; An, L.-K.; Liu, H.-Y. Meroterpenoids with antitumor activities from guava (Psidium guajava). J. Agric. Food Chem. 2017, 65, 4993–4999. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, G.-T.; Li, Y.; Hai, P.; Wang, F.; Liu, J.-K. Guajadials CF, four unusual meroterpenoids from Psidium guajava. Nat. Prod. Bioprospecting 2013, 3, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ouyang, W.; Pan, C.; Gao, Z.; Han, Y.; Song, M.; Feng, K.; Xiao, H.; Cao, Y. Identification of a new benzophenone from Psidium guajava L. leaves and its antineoplastic effects on human colon cancer cells. Food Funct. 2019, 10, 4189–4198. [Google Scholar] [CrossRef] [PubMed]
- Bazioli, J.M.; Costa, J.H.; Shiozawa, L.; Ruiz, A.L.T.G.; Foglio, M.A.; Carvalho, J.E.d. Anti-estrogenic activity of guajadial fraction, from guava leaves (Psidium guajava L.). Molecules 2020, 25, 1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.-H.; Wang, Z.-H.; Meng, D.-L.; Li, X. Cytotoxic and antioxidant constituents from the leaves of Psidium guajava. Bioorganic Med. Chem. Lett. 2015, 25, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Kaileh, M.; Berghe, W.V.; Boone, E.; Essawi, T.; Haegeman, G. Screening of indigenous Palestinian medicinal plants for potential anti-inflammatory and cytotoxic activity. J. Ethnopharmacol. 2007, 113, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Sukanya, S.; Sinthu, K.; Shazhni, J.; Priya, F.S. Qualitative phytochemical screening and assessment of antimicrobial activity of Psidium guajava and its cytotoxic studies. Asian J. Appl. Sci. Technol. 2017, 1, 414–420. [Google Scholar]
- Manikyam, H.K.; Joshi, S.K.; Vakadi, S.; Patil, S.B. Anticancer activity of terpenoid saponin extract of Psidium guajava on MCF-7 cancer cell line using DAPI and MTT assays. Afr. J. Pharm. Pharmacol. 2021, 15, 206–211. [Google Scholar] [CrossRef]
- Sulain, M.D.; Zazali, K.E.; Ahmad, N. Screening on anti-proliferative activity of Psidium guajava leaves extract towards selected cancer cell lines. J. US-China Med Sci. 2012, 9, 30–37. [Google Scholar]
- Radice, M.; Chiurato, M.; Guerrini, A.; Lozupone, F. Antiproliferative activity of Psidium guajava essential oil: A preliminary study. In Proceedings of the Conference on Molecular, Biomedical and Computational Sciences and Engineering (MOL2NET’17), Basel, Switzerland, 15 January–15 December 2017. [Google Scholar]
- Lok, B.; Sandai, D.; Baharetha, H.M.; Nazari, V.M.; Asif, M.; Tan, C.S.; Majid, A.A. Anticancer effect of Psidium guajava (Guava) leaf extracts against colorectal cancer through inhibition of angiogenesis. Asian Pac. J. Trop. Biomed. 2020, 10, 293. [Google Scholar] [CrossRef]
- Braga, T.V.; das Dores, R.G.R.; Ramos, C.S.; Evangelista, F.C.G.; da Silva Tinoco, L.M.; de Pilla Varotti, F.; das Graças Carvalho, M.; de Paula Sabino, A. Antioxidant, antibacterial and antitumor activity of ethanolic extract of the Psidium guajava leaves. Am. J. Plant Sci. 2014, 05, 3492–3500. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-C.; Hsieh, C.-L.; Peng, C.-C.; Hsieh-Li, H.-M.; Chiang, H.-S.; Huang, K.-D.; Peng, R.Y. Brain derived metastatic prostate cancer DU-145 cells are effectively inhibited in vitro by guava (Psidium gujava L.) leaf extracts. Nutr. Cancer 2007, 58, 93–106. [Google Scholar] [CrossRef]
- Chen, K.-C.; Hsieh, C.-L.; Huang, K.-D.; Ker, Y.-B.; Chyau, C.-C.; Peng, R.Y. Anticancer activity of rhamnoallosan against DU-145 cells is kinetically complementary to coexisting polyphenolics in Psidium guajava budding leaves. J. Agric. Food Chem. 2009, 57, 6114–6122. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-C.; Peng, C.-H.; Chen, K.-C.; Hsieh, C.-L.; Peng, R.Y. The aqueous soluble polyphenolic fraction of Psidium guajava leaves exhibits potent anti-angiogenesis and anti-migration actions on DU145 cells. Evid.-Based Complement. Altern. Med. 2011, 2011, 2190069. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-C.; Peng, C.-C.; Chiu, W.-T.; Cheng, Y.-T.; Huang, G.-T.; Hsieh, C.-L.; Peng, R.Y. Action mechanism and signal pathways of Psidium guajava L. aqueous extract in killing prostate cancer LNCaP cells. Nutr. Cancer 2010, 62, 260–270. [Google Scholar] [CrossRef]
- Ryu, N.H.; Park, K.-R.; Kim, S.-M.; Yun, H.-M.; Nam, D.; Lee, S.-G.; Jang, H.-J.; Ahn, K.S.; Kim, S.-H.; Shim, B.S. A hexane fraction of guava leaves (Psidium guajava L.) induces anticancer activity by suppressing AKT/mammalian target of rapamycin/ribosomal p70 S6 kinase in human prostate cancer cells. J. Med. Food 2012, 15, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.-T.; Ko, S.-C.; Oh, G.-W.; Heo, S.-Y.; Jung, W.-K. Effects of Psidium guajava leaf extract on apoptosis induction through mitochondrial dysfunction in HepG2 cells. Microbiol. Biotechnol. Lett. 2019, 47, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Joseph, B.; Priya, R.M. Preliminary phytochemicals of Psidium guajava L. leaf of methanol extract and its cytotoxic study on HeLa cell lines. Inventi Rapid Ethnopharmacol. 2010, 1, 1–3. [Google Scholar]
- Joseph, B.; Priya, R.M.; Helen, P.M.; Sujatha, S. Bio-active compounds in essential oil and its effects of antimicrobial, cytotoxic activity from the Psidium guajava (L.) leaf. J. Adv. Biotechnol. 2010, 9, 10–14. [Google Scholar]
- Fathilah, A.; Sujata, R.; Norhanom, A.; Adenan, M. Antiproliferative activity of aqueous extract of Piper betle L. and Psidium guajava L. on KB and HeLa cell lines. J. Med. Plants Res. 2010, 4, 987–990. [Google Scholar] [CrossRef]
- Levy, A.S.; Carley, S. Cytotoxic activity of hexane extracts of Psidium guajava L. (Myrtaceae) and Cassia alata L. (Caesalpineaceae) in Kasumi-1 and OV2008 cancer cell lines. Trop. J. Pharm. Res. 2012, 11, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, A.; Sarfraz, R.A.; Rashid, M.A.; Mahmood, A.; Shahid, M.; Noor, N. Chemical composition, antioxidant, antitumor, anticancer and cytotoxic effects of Psidium guajava leaf extracts. Pharm. Biol. 2016, 54, 1971–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phonarknguen, R.; Nobsathian, S.; Assawasuparerk, K. Effect of Betulinic acid Extraction from Guava (Psidium guajava Linn.) Leaves Against Human Cholangiocarcinoma Cells. Asian Pac. J. Cancer Prev. 2022, 23, 583–590. [Google Scholar] [CrossRef]
- Dos Santos, R.C.; Ombredane, A.S.; Souza, J.M.T.; Vasconcelos, A.G.; Plácido, A.; das GN Amorim, A.; Barbosa, E.A.; Lima, F.C.; Ropke, C.D.; Alves, M.M. Lycopene-rich extract from red guava (Psidium guajava L.) displays cytotoxic effect against human breast adenocarcinoma cell line MCF-7 via an apoptotic-like pathway. Food Res. Int. 2018, 105, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.G.; Valim, M.O.; Amorim, A.G.; do Amaral, C.P.; de Almeida, M.P.; Borges, T.K.; Socodato, R.; Portugal, C.C.; Brand, G.D.; Mattos, J.S.C. Cytotoxic activity of poly-ɛ-caprolactone lipid-core nanocapsules loaded with lycopene-rich extract from red guava (Psidium guajava L.) on breast cancer cells. Food Res. Int. 2020, 136, 109548. [Google Scholar] [CrossRef]
- Vasconcelos, A.G.; Barros, A.L.A.; Cabral, W.F.; Moreira, D.C.; da Silva, I.G.M.; Silva-Carvalho, A.É.; de Almeida, M.P.; Albuquerque, L.F.; dos Santos, R.C.; S Brito, A.K. Promising self-emulsifying drug delivery system loaded with lycopene from red guava (Psidium guajava L.): In vivo toxicity, biodistribution and cytotoxicity on DU-145 prostate cancer cells. Cancer Nanotechnol. 2021, 12, 30. [Google Scholar] [CrossRef]
- Karia, P.D.; Patil, L.A.; Vakani, M.S.; Chauhan, G.M.; Gupta, S.S.; Rathod, S.P.; Patel, K.V. Chemoprevention of breast cancer by Psidium guajava Linn. Asian J. Pharm. Pharmacol. 2018, 5, 58–68. [Google Scholar] [CrossRef]
- Chadarat, A.; Siriporn, O.; Songyot, A. Cytotoxicity of extracts from fruit plants against leukemic cell lines. Afr. J. Pharm. Pharmacol. 2010, 4, 13–21. [Google Scholar] [CrossRef]
- Bontempo, P.; Doto, A.; Miceli, M.; Mita, L.; Benedetti, R.; Nebbioso, A.; Veglione, M.; Rigano, D.; Cioffi, M.; Sica, V. Psidium guajava L. anti-neoplastic effects: Induction of apoptosis and cell differentiation. Cell Prolif. 2011, 45, 22–31. [Google Scholar] [CrossRef]
- Priam, F.; Marcelin, O.; Marcus, R.; Wijkhuisen, A.; Smith-Ravin, E.J. Evaluation of Anti-Cancer Effects of Lycopene extracted from Pink Guava Psidium guajava L. and its Combination with Apigenin or Resveratrol. Res. Sq. 2021. Preprint. [Google Scholar] [CrossRef]
- Salib, J.Y.; Michael, H.N. Cytotoxic phenylethanol glycosides from Psidium guaijava seeds. Phytochemistry 2004, 65, 2091–2093. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.S.; Yadav, S.A.; Surendren, L.K. Evaluation of in vitro anticancer potential in Punica granatum, Psidium guajava, and Vitis vinifera seed extracts. Int. J. Res. Pharm. Sci. 2019, 10, 165–169. [Google Scholar] [CrossRef]
- Lin, H.-C.; Lin, J.-Y. Immune cell–conditioned media suppress prostate cancer PC-3 cell growth correlating with decreased proinflammatory/anti-inflammatory cytokine ratios in the media using 5 selected crude polysaccharides. Integr. Cancer Ther. 2016, 15, NP13–NP25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-C.; Lin, J.-Y. GSF3, a polysaccharide from guava (Psidium guajava L.) seeds, inhibits MCF-7 breast cancer cell growth via increasing Bax/Bcl-2 ratio or Fas mRNA expression levels. Int. J. Biol. Macromol. 2020, 161, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-C.; Lin, J.-Y. Pharmacological effects of guava (Psidium guajava L.) seed polysaccharides: GSF3 inhibits PC-3 prostate cancer cell growth through immunotherapy in vitro. Int. J. Mol. Sci. 2021, 22, 3631. [Google Scholar] [CrossRef] [PubMed]
- Cates, R.G.; Prestwich, B.; Innes, A.; Rowe, M.; Stanley, M.; Williams, S.; Thompson, A.; McDonald, S.; Cates, S.; Shrestha, G. Evaluation of the activity of Guatemalan medicinal plants against cancer cell lines and microbes. J. Med. Plant Res. 2013, 4, 2616–2627. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Manekeng, H.T.; Nguenang, G.S.; Dzotam, J.K.; Kuete, V.; Efferth, T. Cytotoxicity of 18 Cameroonian medicinal plants against drug sensitive and multi-factorial drug resistant cancer cells. J. Ethnopharmacol. 2018, 222, 21–33. [Google Scholar] [CrossRef]
- Lee, S.-B.; Park, H.-R. Anticancer activity of guava (Psidium guajava L.) branch extracts against HT-29 human colon cancer cells. J. Med. Plant Res. 2010, 4, 891–896. [Google Scholar] [CrossRef]
- Seo, N.; Ito, T.; Wang, N.; Yao, X.; Tokura, Y.; Furukawa, F.; Takigawa, M.; Kitanaka, S. Anti-allergic Psidium guajava extracts exert an antitumor effect by inhibition of T regulatory cells and resultant augmentation of Th1 cells. Anticancer Res. 2005, 25, 3763–3770. [Google Scholar] [PubMed]
- Wang, Y.; Duan, M.; Zhao, L.; Ma, P. Guajadial inhibits NSCLC growth and migration following activation of the VEGF receptor-2. Fitoterapia 2018, 129, 73–77. [Google Scholar] [CrossRef]
- Beniston, R.G.; Campo, M.S. Quercetin elevates p27Kip1 and arrests both primary and HPV16 E6/E7 transformed human keratinocytes in G1. Oncogene 2003, 22, 5504–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-A.; Kim, J.-Y.; Lee, J.-Y.; Kang, C.-M.; Kwon, H.-J.; Yoo, Y.-D.; Kim, T.-W.; Lee, Y.-S.; Lee, S.-J. Induction of cell cycle arrest and apoptosis in human breast cancer cells by quercetin. Int. J. Oncol. 2001, 19, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.S.; Tran, E.; Nguyen, T.T.; Ong, C.K.; Lee, S.K.; Lee, J.J.; Ng, C.P.; Leong, C.; Huynh, H. Quercetin-induced growth inhibition and cell death in nasopharyngeal carcinoma cells are associated with increase in Bad and hypophosphorylated retinoblastoma expressions. Oncol. Rep. 2004, 11, 727–733. [Google Scholar] [CrossRef]
- Baud, V.; Karin, M. Is NF-κB a good target for cancer therapy? Hopes and pitfalls. Nat. Rev. Drug Discov. 2009, 8, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Takahashi, Y.; Sakashita, T.; Inoue, H.; Tanabe, T.; Yoshimoto, T. Growth stimulation and induction of epidermal growth factor receptor by overexpression of cyclooxygenases 1 and 2 in human colon carcinoma cells. Acta (BBA)-Mol. Cell Biol. Lipids 1999, 1438, 120–130. [Google Scholar] [CrossRef]
- Menter, D.G.; DuBois, R.N. Prostaglandins in cancer cell adhesion, migration, and invasion. Int. J. Cell Biol. 2012, 2012, 723419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, F.; Kannangai, R.; Adegbola, O.; Wang, J.; Su, G.; Torbenson, M. mTOR and P70 S6 kinase expression in primary liver neoplasms. Clin. Cancer Res. 2004, 10, 8421–8425. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Zhang, W.; Bertram, P.; Zheng, X.F.; McLeod, H. Pharmacogenomic profiling of the PI3K/PTEN-AKT-mTOR pathway in common human tumors. Int. J. Oncol. 2004, 24, 893–900. [Google Scholar] [CrossRef]
- Testa, J.R.; Bellacosa, A. AKT plays a central role in tumorigenesis. Proc. Natl. Acad. Sci. USA 2001, 98, 10983–10985. [Google Scholar] [CrossRef] [Green Version]
- Wendel, H.-G.; Stanchina, E.d.; Fridman, J.S.; Malina, A.; Ray, S.; Kogan, S.; Cordon-Cardo, C.; Pelletier, J.; Lowe, S.W. Survival signaling by Akt and eIF4E in oncogenesis and cancer therapy. Nature 2004, 428, 332–337. [Google Scholar] [CrossRef]
- Knight, T.; Irving, J.A.E. Ras/Raf/MEK/ERK pathway activation in childhood acute lymphoblastic leukemia and its therapeutic targeting. Front. Oncol. 2014, 4, 160. [Google Scholar] [CrossRef] [PubMed]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK–RAS–RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhouser, M.L. Dietary flavonoids and cancer risk: Evidence from human population studies. Nutr. Cancer 2004, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Rankin, G.O.; Liu, L.; Daddysman, M.K.; Jiang, B.-H.; Chen, Y.C. Kaempferol inhibits angiogenesis and VEGF expression through both HIF dependent and independent pathways in human ovarian cancer cells. Nutr. Cancer 2009, 61, 554–563. [Google Scholar] [CrossRef]
- Marfe, G.; Tafani, M.; Indelicato, M.; Sinibaldi-Salimei, P.; Reali, V.; Pucci, B.; Fini, M.; Russo, M.A. Kaempferol induces apoptosis in two different cell lines via Akt inactivation, Bax and SIRT3 activation, and mitochondrial dysfunction. J. Cell. Biochem. 2009, 106, 643–650. [Google Scholar] [CrossRef]
- Yi, X.; Zuo, J.; Tan, C.; Xian, S.; Luo, C.; Chen, S.; Yu, L.; Luo, Y. Kaempferol, a flavonoid compound from gynura medica induced apoptosis and growth inhibition in MCF-7 breast cancer cell. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Giovannucci, E.; Ascherio, A.; Rimm, E.B.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Intake of carotenoids and retino in relation to risk of prostate cancer. J. Natl. Cancer Inst. 1995, 87, 1767–1776. [Google Scholar] [CrossRef]
- Gloria, N.F.; Soares, N.; Brand, C.; Oliveira, F.L.; Borojevic, R.; Teodoro, A.J. Lycopene and beta-carotene induce cell-cycle arrest and apoptosis in human breast cancer cell lines. Anticancer Res. 2014, 34, 1377–1386. [Google Scholar]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Takeshima, M.; Nakano, S. Mechanism of the anticancer effect of lycopene (tetraterpenoids). Enzymes 2015, 37, 139–166. [Google Scholar] [CrossRef]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer molecular mechanisms of resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, D.A.; Ruscetti, F.W.; Gallo, R. Selective in vitro growth of T lymphocytes from normal human bone marrows. Science 1976, 193, 1007–1008. [Google Scholar] [CrossRef]
- Zhou, Y.; Weyman, C.M.; Liu, H.; Almasan, A.; Zhou, A. IFN-γ induces apoptosis in HL-60 cells through decreased Bcl-2 and increased Bak expression. J. Interf. Cytokine Res. 2008, 28, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Lomelí, P.; Bravo-Cuellar, A.; Hernández-Flores, G.; Jave-Suárez, L.F.; Aguilar-Lemarroy, A.; Lerma-Díaz, J.M.; Domínguez-Rodríguez, J.R.; Sánchez-Reyes, K.; Ortiz-Lazareno, P.C. Increase of IFN-γ and TNF-γ production in CD107a+ NK-92 cells co-cultured with cervical cancer cell lines pre-treated with the HO-1 inhibitor. Cancer Cell Int. 2014, 14, 100. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Jaw, J.J.; Stutzman, N.C.; Zou, Z.; Sun, P.D. Natural killer cell-produced IFN-γ and TNF-α induce target cell cytolysis through up-regulation of ICAM-1. J. Leukoc. Biol. 2011, 91, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Beatty, G.L.; Paterson, Y. IFN-γ-dependent inhibition of tumor angiogenesis by tumor-infiltrating CD4+ T cells requires tumor responsiveness to IFN-γ. J. Immunol. 2001, 166, 2276–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.L.; Jang, J.W.; Lee, S.W.; Yoo, S.H.; Kwon, J.H.; Nam, S.W.; Bae, S.H.; Choi, J.Y.; Han, N.I.; Yoon, S.K. Inflammatory cytokines and change of Th1/Th2 balance as prognostic indicators for hepatocellular carcinoma in patients treated with transarterial chemoembolization. Sci. Rep. 2019, 9, 3260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostoglou-Athanassiou, I.; Ntalles, K.; Markopoulos, C.; Athanassiou, P.; Gogas, J.; Proukakis, C. Thyroid function in postmenopausal women with breast cancer on tamoxifen. Eur. J. Gynaecol. Oncol. 1998, 19, 150–154. [Google Scholar]
- Lim, T.K.; Khoo, K.C. Guava in Malaysia: Production, Pests and Diseases; Tropical Press: Kuala Lumpur, Malaysia, 1990. [Google Scholar]
- Akerele, O. Nature’s medicinal bounty: Don’t throw it away. World Health Forum 1993, 14, 390–395. [Google Scholar]
- Wei, L.; Li, Z.; Chen, B. Clinical study on treatment of infantile rotaviral enteritis with Psidium guajava L. Chin. J. Integr. Tradit. West. Med. 2001, 7, 86–89. [Google Scholar] [CrossRef]
- Lozoya, X.; Reyes-Morales, H.; Chávez-Soto, M.A.; del Carmen Martínez-García, M.a.; Soto-González, Y.; Doubova, S.V. Intestinal anti-spasmodic effect of a phytodrug of Psidium guajava folia in the treatment of acute diarrheic disease. J. Ethnopharmacol. 2002, 83, 19–24. [Google Scholar] [CrossRef]
- Echemendía Salís, C.E.; Morón Rodríguez, F.J. Tintura de hojas de Psidium guajava L. en pacientes con diarrea aguda simple. Rev. Cuba. Plantas Med. 2004, 9. [Google Scholar]
- Cheng, J.-T.; Yang, R.-S. Hypoglycemic effect of guava juice in mice and human subjects. Am. J. Chin. Med. 1983, 11, 74–76. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Schwarzinger, B.; Stadlbauer, V.; Lanzerstorfer, P.; Iken, M.; Schwarzinger, C.; Kolb, P.; Schwarzinger, S.; Mörwald, K.; Brunner, S. Guava (Psidium guajava) fruit extract prepared by supercritical CO2 extraction inhibits intestinal glucose resorption in a double-blind, randomized clinical study. Nutrients 2019, 11, 1512. [Google Scholar] [CrossRef] [Green Version]
- Kakuo, S.; Fushimi, T.; Kawasaki, K.; Nakamura, J.; Ota, N. Effects of Psidium guajava Linn. leaf extract in Japanese subjects with knee pain: A randomized, double-blind, placebo-controlled, parallel pilot study. Aging Clin. Exp. Res. 2018, 30, 1391–1398. [Google Scholar] [CrossRef]
- Guintu, F.Z.; Chua, A.H. Effectivity of guava leaves (Psidium guajava) as mouthwash for patients with aphthous ulcers. Philipp. J. Otolaryngol. Head Neck Surg. 2013, 28, 8–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part of Plant | Type of Extracts | Type of Cancer | References |
|---|---|---|---|
| Leaves | Dichloromethane | NCI-H460 (lung cancer), MCF-7 (breast cancer), HT-29 (colon cancer), PC-3 (prostate cancer), K562 (leukemia), 786-0 (kidney cancer), OVCAR-3, NCI/ADR-RES (ovarian cancer), and UACC-62 (melanoma) | Rizzo et al., 2014 [8] |
| Leaves | Dichloromethane | MCF-7 and MCF-7 BUS (breast cancer) | Bazioli et al., 2020 [20] |
| Leaves | Methanol | A549 (lung cancer), MCF-7 (breast cancer), SMMC-7721 (cervical cancer), and HL60 (leukemia) | Gao et al., 2013 [18] |
| Leaves | 75% ethanol | A549 (lung cancer), HeLa (cervical cancer), and SGC-7901 (gastric cancer) | Feng et al., 2015 [21] |
| Leaves | Dichloromethane and methanol (1:1, v/v) | MCF-7 (breast cancer) | Kaileh et al., 2007 [22] |
| Leaves | Aqueous | MCF-7 (breast cancer) | Sukanya et al., 2017 [23] |
| Leaves | Methanol | MCF-7 (breast cancer) | Manikyam et al., 2021 [24] |
| Leaves | Petroleum ether, methanol, water | MDA-MB-231 (breast cancer), MG-63 (bone osteosarcoma) | Sulain et al., 2012 [25] |
| Leaves | Methanol | MDA-MB-231 (breast cancer), MG-63 (bone osteosarcoma) | Sulain et al., 2012 [25] |
| Leaves | Aqueous | MDA-MB-231 (breast cancer), MG-63 (bone osteosarcoma) | Sulain et al., 2012 [25] |
| Leaves | Polyphenol-rich fraction 50% ethanol (v/v) extract | COLO320DM (colon cancer) | Kawakami et al., 2009 [9] |
| Leaves | Essential oil | HCT116, Caco-2, and SW620 (colon cancer) | Radice et al., 2017 [26] |
| Leaves | 70% ethanol | HCT116 and HT-29 (colon cancer) | Zhu et al., 2019 [19] |
| Leaves | Aqueous, Ethanol | HCT116 (colon cancer) | Lok et al., 2020 [27] |
| Leaves | Ethanol | RKO-AS45-1 (colorectal cancer) and HeLa (cervical cancer) | Braga et al., 2014 [28] |
| Leaves | Aqueous | DU145 (prostate cancer) | Chen et al., 2007 [29]; Chen et al., 2009 [30] |
| Leaves | Aqueous | DU145 (prostate cancer) | Peng et al., 2011 [31] |
| Leaves | Aqueous | LNCaP (prostate cancer) | Chen et al., 2010 [32] |
| Leaves | Hexane fraction of methanol extract | PC-3 (prostate cancer) | Ryu et al., 2012 [33] |
| Leaves | 80% aqueous methanol | HepG2 (liver cancer) | Nguyen et al., 2019 [34] |
| Leaves | Methanol | HeLa (cervical cancer) | Joseph and Priya, 2010 [35] |
| Leaves | Essential oil | HeLa (cervical cancer) | Joseph et al., 2010 [36] |
| Leaves | Aqueous | HeLa (cervical cancer) and KB (mouth epidermal cancer) | Fathilah et al., 2010 [37] |
| Leaves | Hexane fraction of methanol extract | Kasumi-1 (leukemia) | Levy and Carley, 2012 [38] |
| Leaves | Essential oil | P388 (leukemia) and KB (mouth epidermal cancer) | Manosroi et al., 2006 [10] |
| Leaves | Hexane | KBM5 (leukemia), U266 (multiple myeloma), and SCC4 (tongue carcinoma) | Ashraf et al., 2016 [39] |
| Leaves | Chloroform | U266 (multiple myeloma) | Ashraf et al., 2016 [39] |
| Leaves | 70% ethanol | HuCCA (cholangiocarcinoma) | Phonarknguen et al., 2022 [40] |
| Fruits | Petroleum ether | A549 (lung cancer), HCT116 (colon cancer), CCRF-CEM (leukemia), DU145 (prostate cancer), and Huh7 (liver cancer) | Qin et al., 2017 [17] |
| Fruits | Ethanol | MCF-7 (breast cancer) | dos Santos et al., 2018 [41] |
| Fruits | Ethanol | MCF-7 (breast cancer) | Vasconcelos et al., 2020 [42] |
| Fruits | Ethanol | DU145 (prostate cancer) | Vasconcelos et al., 2021 [43] |
| Fruits, pulp | - | MCF-7, MDA-MB-231, and MDA-MB-453 (breast cancer) | Karia et al., 2019 [44] |
| Fruits | Ethanol | HL-60 (leukemia) | Chadarat et al., 2010 [45] |
| Fruits, pulp | Acetone | MDA-MB 231 (breast cancer) and NB4 (leukemia) | Bontempo et al., 2012 [46] |
| Fruits | Carotenoid | LNCaP (prostate cancer) and UACC257 (melanoma) | Priam et al., 2021 [47] |
| Seeds | 80% Aqueous acetone | EAC (skin cancer) and P388 (leukemia) | Salib and Michael, 2004 [48] |
| Seeds | Methanol, ethyl acetate | A549 (lung cancer) | Nelson et al., 2019 [49] |
| Seeds | Aqueous | MCF-7 (breast cancer), PC-3 (prostate cancer) | Lin and Lin, 2016 [50], 2020 [51], 2021 [52] |
| Barks | Acetone | MCF-7 (breast cancer), CAL 27 (tongue carcinoma) | Cates et al., 2013 [53] |
| Barks | Methanol | MDA-MB-231 (breast cancer), HCT116 (colon cancer), HepG2 (liver cancer), CCRF-CEM (leukemia), and U87MG (brain cancer) | Mbaveng et al., 2018 [54] |
| Branches | Acetone | HT-29 (colon cancer) | Lee and Park, 2010 [55] |
| Leaves, bark, and roots | Ethanol | B16 melanoma (skin cancer) | Seo et al., 2005 [56] |
| Bioactive Molecules of Interest | Part of Plant | Type of Cancer Cells | Anticancer Mechanism(s) | References |
|---|---|---|---|---|
| 3,5-dihydroxy-2,4-dimethyl-1-O-(6’-O-galloyl-β-d-glucopyranosyl)-benzophenone | Leaves | HCT116 and HT-29 (colon cancer) | Antiproliferative, apoptosis | Zhu et al., 2019 [19] |
| 4,5-Diepipsidial A | Fruits | DU145 (prostate cancer), Huh7 (liver cancer) | Antiproliferative | Qin et al., 2017 [17] |
| Apigenin | Fruits | LNCaP (prostate cancer) and UACC257 (melanoma) | Antiproliferative, apoptosis, antimigration, and immune response | Priam et al., 2021 [47] |
| Betulinic acid | Leaves | HuCCA (cholangiocarcinoma) | Antiproliferative and apoptosis | Phonarknguen et al., 2022 [40] |
| Guajadial | Leaves | A549 and H1650 (lung cancer), MCF-7 and MCF-7 BUS (breast cancer) | Antiproliferative and antimigration | Rizzo et al., 2014 [8] |
| Guajadial B | Fruits | Huh7 (liver cancer) | Antiproliferative | Qin et al., 2017 [17] |
| Guajadial D | Leaves, Fruits | A549 (lung cancer), CCRF-CEM and HL60 (leukemia), HCT116 (colon cancer), and SMMC-7721 (cervical cancer) | Antiproliferative | Gao et al., 2013 [18] and Qin et al., 2017 [17] |
| Guajadial F | Leaves | A549 (lung cancer), MCF-7 (breast cancer), HL60 (leukemia), and SMMC-7721 (cervical cancer) | Antiproliferative | Gao et al., 2013 [18] |
| Guavinoside B | Leaves | HCT116 and HT-29 (colon cancer) | Antiproliferative | Zhu et al., 2019 [19] |
| Guavinoside C (Avicularin) | Leaves | A549 (lung cancer), HeLa (cervical cancer), and SGC-7901 (gastric cancer) | Antiproliferative | Feng et al., 2015 [21] |
| Kaempferol | Leaves | Kasumi-1 (leukemia) | Apoptosis, antiangiogenesis, and antimigration | Levy and Carley, 2012 [38] |
| Lycopene | Fruits | DU145 (prostate cancer), LNCaP (prostate cancer), MCF-7 (breast cancer), and UACC257 (melanoma) | Antiproliferative, antioxidative, antiangiogenesis, and antimigration | dos Santos et al., 2018 [41], Vasconcelos et al., 2020 [42], Vasconcelos et al., 2021 [43], and Priam et al., 2021 [47] |
| Psiguadial D | Fruits | DU145 (prostate cancer) | Antiproliferative | Qin et al., 2017 [17] |
| Quercetin | Leaves | A549 (lung cancer), HeLa (cervical cancer), Kasumi-1 (leukemia), and SGC-7901 (gastric cancer) | Antiproliferative, apoptosis, and antioxidative | Feng et al., 2015 [21] and Levy and Carley, 2012 [38] |
| Resveratrol | Fruits | UACC257 (melanoma) | Antiproliferative, apoptosis, antiangiogenesis, and antimigration | Priam et al., 2021 [47] |
| Part of Plant | Type of Extracts | Type of Cancer | In Vivo Animal Model | References |
|---|---|---|---|---|
| Leaves, bark, and roots | Ethanol | B16 melanoma (skin cancer) | Subcutaneous inoculation of cancer cells into female B6 and BALB/c mice | Seo et al., 2005 [56] |
| Leaves | Aqueous | LNCaP (prostate cancer) | Subcutaneous inoculation of cancer cells into male BALB/c mice | Chen et al., 2010 [32] |
| Leaves | Dichloromethane | EAC (skin cancer) | Subcutaneous inoculation of cancer cells into female BALB/c mice | Rizzo et al., 2014 [8] |
| Fruits, pulp | - | Breast cancer | MNU-induced rat mammary tumors in female Sprague Dawley rats | Karia et al., 2019 [44] |
| Seeds | 80% Aqueous acetone | EAC (skin cancer) | Intraperitoneal inoculation into female Swiss albino mice | Salib and Michael, 2004 [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lok, B.; Babu, D.; Tabana, Y.; Dahham, S.S.; Adam, M.A.A.; Barakat, K.; Sandai, D. The Anticancer Potential of Psidium guajava (Guava) Extracts. Life 2023, 13, 346. https://doi.org/10.3390/life13020346
Lok B, Babu D, Tabana Y, Dahham SS, Adam MAA, Barakat K, Sandai D. The Anticancer Potential of Psidium guajava (Guava) Extracts. Life. 2023; 13(2):346. https://doi.org/10.3390/life13020346
Chicago/Turabian StyleLok, Bronwyn, Dinesh Babu, Yasser Tabana, Saad Sabbar Dahham, Mowaffaq Adam Ahmed Adam, Khaled Barakat, and Doblin Sandai. 2023. "The Anticancer Potential of Psidium guajava (Guava) Extracts" Life 13, no. 2: 346. https://doi.org/10.3390/life13020346
APA StyleLok, B., Babu, D., Tabana, Y., Dahham, S. S., Adam, M. A. A., Barakat, K., & Sandai, D. (2023). The Anticancer Potential of Psidium guajava (Guava) Extracts. Life, 13(2), 346. https://doi.org/10.3390/life13020346