
Sleep and Core Body Temperature Alterations Induced by Space Radiation in Rats

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Irradiation Procedure
2.3. Surgery
2.4. Sleep Recording and Scoring
2.5. EEG Analyses
2.6. Activity Analyses
2.7. Temperature Analyses
2.8. Statistical Analyses
3. Results
3.1. Sleep Amounts
3.2. EEG Spectra
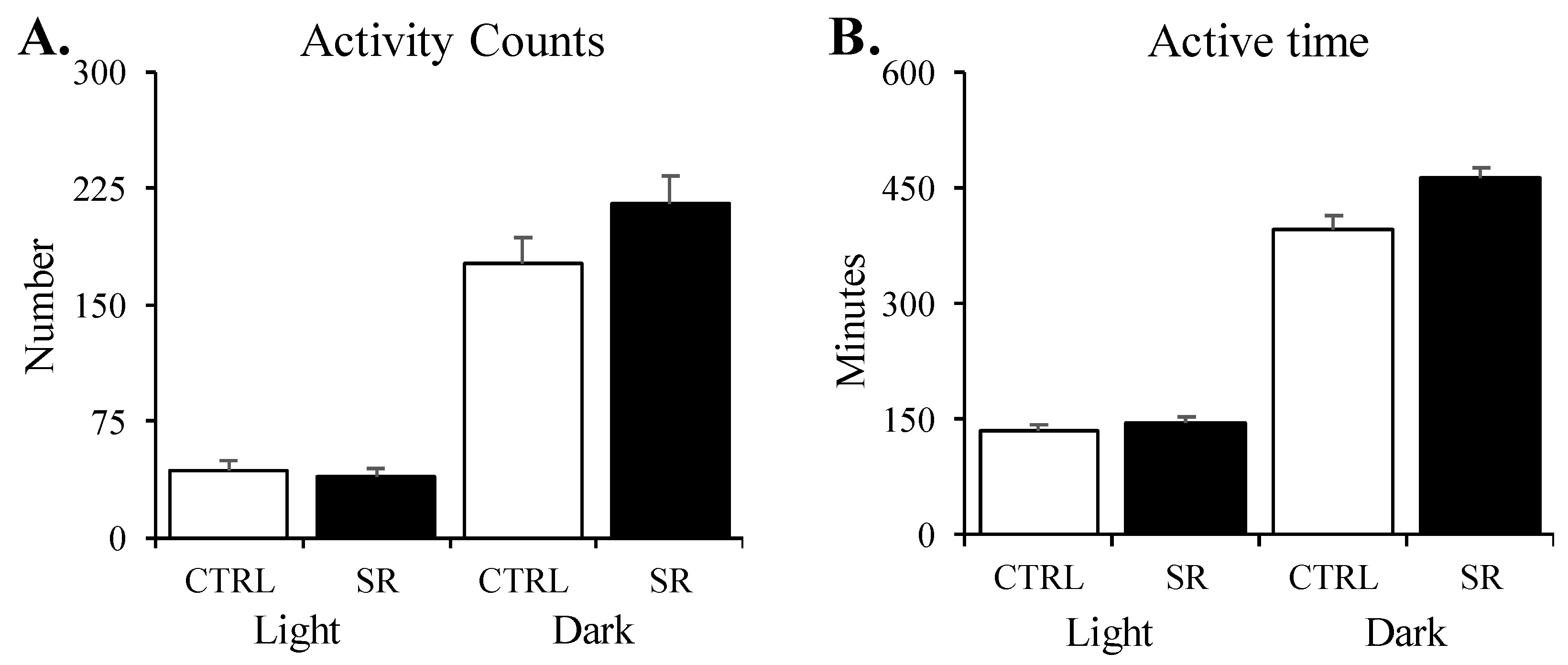
3.3. Activity
3.4. Temperature
3.5. Hourly Alterations in Activity, Sleep, and Temperature
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ranjbaran, Z.; Keefer, L.; Stepanski, E.; Farhadi, A.; Keshavarzian, A. The relevance of sleep abnormalities to chronic inflammatory conditions. Inflamm. Res. 2007, 56, 51–57. [Google Scholar] [CrossRef]
- Nahrendorf, M.; Swirski, F.K. Lifestyle effects on hematopoiesis and atherosclerosis. Circ. Res. 2015, 116, 884–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.P. Sleep, memory and emotion. Prog. Brain Res. 2010, 185, 49–68. [Google Scholar] [CrossRef]
- Rasch, B.; Born, J. About sleep’s role in memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef]
- Ognjanovski, N.; Broussard, C.; Zochowski, M.; Aton, S.J. Hippocampal Network Oscillations Rescue Memory Consolidation Deficits Caused by Sleep Loss. Cereb. Cortex 2018, 28, 3711–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havekes, R.; Meerlo, P.; Abel, T. Animal Studies on the Role of Sleep in Memory: From Behavioral Performance to Molecular Mechanisms. Curr. Top Behav. Neurosci. 2015, 25, 183–206. [Google Scholar] [CrossRef]
- Barger, L.K.; Flynn-Evans, E.E.; Kubey, A.; Walsh, L.; Ronda, J.M.; Wang, W.; Wright, K.P., Jr.; Czeisler, C.A. Prevalence of sleep deficiency and use of hypnotic drugs in astronauts before, during, and after spaceflight: An observational study. Lancet Neurol. 2014, 13, 904–912. [Google Scholar] [CrossRef] [Green Version]
- Frost, J.D., Jr.; Shumate, W.H.; Salamy, J.G.; Booher, C.R. Sleep monitoring: The second manned Skylab mission. Aviat. Space Environ. Med. 1976, 47, 372–382. [Google Scholar] [PubMed]
- Santy, P.A.; Kapanka, H.; Davis, J.R.; Stewart, D.F. Analysis of sleep on Shuttle missions. Aviat. Space Environ. Med. 1988, 59, 1094–1097. [Google Scholar]
- NASA/TM-2013-217378; Sleep Quality Questionnaire Short-Duration Flyers. Johnson Space Center: Houston, TX, USA, 2013.
- Basner, M.; Dinges, D.F. Lost in space: Sleep. Lancet Neurol. 2014, 13, 860–862. [Google Scholar] [CrossRef]
- Zhou, Y.; Ni, H.; Li, M.; Sanzari, J.K.; Diffenderfer, E.S.; Lin, L.; Kennedy, A.R.; Weissman, D. Effect of solar particle event radiation and hindlimb suspension on gastrointestinal tract bacterial translocation and immune activation. PLoS ONE 2012, 7, e44329. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Holmes, V.; Zhou, Y.; Ni, H.; Sanzari, J.K.; Kennedy, A.R.; Weissman, D. Hindlimb suspension and SPE-like radiation impairs clearance of bacterial infections. PLoS ONE 2014, 9, e85665. [Google Scholar] [CrossRef]
- Musacchia, X.J.; Fagette, S. Weightlessness simulations for cardiovascular and muscle systems: Validity of rat models. J. Gravit. Physiol. 1997, 4, 49–59. [Google Scholar]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading of growing rats: A model for predicting skeletal changes during space flight. Bone 1998, 22, 83S–88S. [Google Scholar] [CrossRef]
- Doty, S.B.; Vico, L.; Wronski, T.; Morey-Holton, E. Use of animal models to study skeletal effects of space flight. Adv. Space Biol. Med. 2005, 10, 209–224. [Google Scholar] [PubMed]
- Mader, T.H.; Gibson, C.R.; Pass, A.F.; Kramer, L.A.; Lee, A.G.; Fogarty, J.; Tarver, W.J.; Dervay, J.P.; Hamilton, D.R.; Sargsyan, A.; et al. Optic disc edema, globe flattening, choroidal folds, and hyperopic shifts observed in astronauts after long-duration space flight. Ophthalmology 2011, 118, 2058–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ombergen, A.; Demertzi, A.; Tomilovskaya, E.; Jeurissen, B.; Sijbers, J.; Kozlovskaya, I.B.; Parizel, P.M.; Van de Heyning, P.H.; Sunaert, S.; Laureys, S.; et al. The effect of spaceflight and microgravity on the human brain. J. Neurol. 2017, 264, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedler, B.; Crapser, J.; McCullough, L. One is the deadliest number: The detrimental effects of social isolation on cerebrovascular diseases and cognition. Acta Neuropathol. 2015, 129, 493–509. [Google Scholar] [CrossRef] [Green Version]
- Slaba, T.C.; Blattnig, S.R.; Norbury, J.W.; Rusek, A.; La Tessa, C. Reference field specification and preliminary beam selection strategy for accelerator-based GCR simulation. Life Sci. Space Res. 2016, 8, 52–67. [Google Scholar] [CrossRef]
- Britten, R.A.; Wellman, L.L.; Sanford, L.D. Progressive increase in the complexity and translatability of rodent testing to assess space-radiation induced cognitive impairment. Neurosci. Biobehav. Rev. 2021, 126, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Whoolery, C.W.; Yun, S.; Reynolds, R.P.; Lucero, M.J.; Soler, I.; Tran, F.H.; Ito, N.; Redfield, R.L.; Richardson, D.R.; Shih, H.Y.; et al. Multi-domain cognitive assessment of male mice shows space radiation is not harmful to high-level cognition and actually improves pattern separation. Sci. Rep. 2020, 10, 2737. [Google Scholar] [CrossRef] [Green Version]
- Kiffer, F.; Boerma, M.; Allen, A. Behavioral effects of space radiation: A comprehensive review of animal studies. Life Sci. Space Res. 2019, 21, 1–21. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Cacao, E. Risks of cognitive detriments after low dose heavy ion and proton exposures. Int. J. Radiat. Biol. 2019, 95, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Cekanaviciute, E.; Rosi, S.; Costes, S.V. Central Nervous System Responses to Simulated Galactic Cosmic Rays. Int. J. Mol. Sci. 2018, 19, 3669. [Google Scholar] [CrossRef] [Green Version]
- Burket, J.A.; Matar, M.; Fesshaye, A.; Pickle, J.C.; Britten, R.A. Exposure to Low (</=10 cGy) Doses of 4He Particles Leads to Increased Social Withdrawal and Loss of Executive Function Performance. Radiat. Res. 2021, 196, 345–354. [Google Scholar] [CrossRef]
- Howe, A.; Kiffer, F.; Alexander, T.C.; Sridharan, V.; Wang, J.; Ntagwabira, F.; Rodriguez, A.; Boerma, M.; Allen, A.R. Long-Term Changes in Cognition and Physiology after Low-Dose (16)O Irradiation. Int. J. Mol. Sci. 2019, 20, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.B.; Mange, A.; Granata, L.; Johnson, B.; Hienz, R.D.; Davis, C.M. Short and Long-Term Changes in Social Odor Recognition and Plasma Cytokine Levels Following Oxygen ((16)O) Ion Radiation Exposure. Int. J. Mol. Sci. 2019, 20, 339. [Google Scholar] [CrossRef] [Green Version]
- Krukowski, K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; Rosi, S. Peripheral T Cells as a Biomarker for Oxygen-Ion-Radiation-Induced Social Impairments. Radiat. Res. 2018, 190, 186–193. [Google Scholar] [CrossRef]
- Krukowski, K.; Grue, K.; Frias, E.S.; Pietrykowski, J.; Jones, T.; Nelson, G.; Rosi, S. Female mice are protected from space radiation-induced maladaptive responses. Brain Behav. Immun. 2018, 74, 106–120. [Google Scholar] [CrossRef]
- Mange, A.; Cao, Y.; Zhang, S.; Hienz, R.D.; Davis, C.M. Whole-Body Oxygen ((16)O) Ion-Exposure-Induced Impairments in Social Odor Recognition Memory in Rats are Dose and Time Dependent. Radiat. Res. 2018, 189, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.M.; DeCicco-Skinner, K.L.; Roma, P.G.; Hienz, R.D. Individual differences in attentional deficits and dopaminergic protein levels following exposure to proton radiation. Radiat. Res. 2014, 181, 258–271. [Google Scholar] [CrossRef]
- Rice, O.V.; Grande, A.V.; Dehktyar, N.; Bruneus, M.; Robinson, J.K.; Gatley, S.J. Long-term effects of irradiation with iron-56 particles on the nigrostriatal dopamine system. Radiat. Environ. Biophys. 2009, 48, 215–225. [Google Scholar] [CrossRef]
- Joseph, J.A.; Hunt, W.A.; Rabin, B.M.; Dalton, T.K.; Harris, A.H. Deficits in the sensitivity of striatal muscarinic receptors induced by 56Fe heavy-particle irradiation: Further “age-radiation” parallels. Radiat. Res. 1993, 135, 257–261. [Google Scholar] [CrossRef]
- Joseph, J.A.; Hunt, W.A.; Rabin, B.M.; Dalton, T.K. Possible “accelerated striatal aging” induced by 56Fe heavy-particle irradiation: Implications for manned space flights. Radiat. Res. 1992, 130, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, A.A.; Schell, B.D.; Osterlund Oltmanns, J.R.; Whishaw, I.Q.; Ton, S.T.; Adamczyk, N.S.; Kartje, G.L.; Britten, R.A.; Wallace, D.G. Skilled movement and posture deficits in rat string-pulling behavior following low dose space radiation ((28)Si) exposure. Behav. Brain Res. 2021, 400, 113010. [Google Scholar] [CrossRef]
- Blackwell, A.A.; Fesshaye, A.; Tidmore, A.; Lake, R.I.; Wallace, D.G.; Britten, R.A. Rapid loss of fine motor skills after low dose space radiation exposure. Behav. Brain Res. 2022, 430, 113907. [Google Scholar] [CrossRef] [PubMed]
- Szymusiak, R.; McGinty, D. Control of slow wave sleep by thermoregulatory mechanisms. Prog. Clin. Biol. Res. 1990, 345, 53–64; discussion 65–66. [Google Scholar]
- McGinty, D.; Szymusiak, R. Keeping cool: A hypothesis about the mechanisms and functions of slow-wave sleep. Trends Neurosci. 1990, 13, 480–487. [Google Scholar] [CrossRef]
- McGinty, D.; Alam, M.N.; Szymusiak, R.; Nakao, M.; Yamamoto, M. Hypothalamic sleep-promoting mechanisms: Coupling to thermoregulation. Arch. Ital. Biol. 2001, 139, 63–75. [Google Scholar]
- Stahn, A.C.; Werner, A.; Opatz, O.; Maggioni, M.A.; Steinach, M.; von Ahlefeld, V.W.; Moore, A.; Crucian, B.E.; Smith, S.M.; Zwart, S.R.; et al. Increased core body temperature in astronauts during long-duration space missions. Sci. Rep. 2017, 7, 16180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinkers, C.H.; van Oorschot, R.; Olivier, B.; Groenink, L. Stress-Induced Hyperthermia in the Mouse. In Mood and Anxiety Related Phenotypes in Mice, Neuromethods; Gould, T.D., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 139–152. [Google Scholar]
- La Tessa, C.; Sivertz, M.; Chiang, I.H.; Lowenstein, D.; Rusek, A. Overview of the NASA space radiation laboratory. Life Sci. Space Res. 2016, 11, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundel, A.; Polyakov, V.V.; Zulley, J. The alteration of human sleep and circadian rhythms during spaceflight. J. Sleep Res. 1997, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans-Flynn, E.; Gregory, K.; Arsintescu, L.; Whitmire, A.; Leveton, L.; Vessey, W. Risk of performance decrements and adverse health outcomes resulting from sleep loss, circadian desynchronization, and work overload. In NASA; Johnson Space Center: Houston, TX, USA, 2015. [Google Scholar]
- Moldofsky, H.; Lue, F.; MacFarlane, J.; Jiang, C.; Poplonski, L.; Ponomoreva, I.; Larina, I.; Gorczynski, R. Long-term effects of microgravity on human sleep, cytokine, and endocrines. Gravitat. Space Biol. Bull. 2000, 14, 41. [Google Scholar]
- Stickgold, R.; Hobson, J.A. REM sleep and sleep efficiency are reduced during space flight. Sleep 1999, 22, 82. [Google Scholar]
- Petit, G.; Cebolla, A.M.; Fattinger, S.; Petieau, M.; Summerer, L.; Cheron, G.; Huber, R. Local sleep-like events during wakefulness and their relationship to decreased alertness in astronauts on ISS. NPJ Microgravity 2019, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanford, L.D.; Suchecki, D.; Meerlo, P. Stress, arousal, and sleep. Curr. Top Behav. Neurosci. 2015, 25, 379–410. [Google Scholar] [CrossRef]
- Sanford, L.D.; Tang, X. Effect of stress on sleep and its relationship to PTSD. In Neurobiology of PTSD; Shiromani, P., Keane, T., LeDoux, J., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 231–253. [Google Scholar]
- Bryant, R.A.; Creamer, M.; O’Donnell, M.; Silove, D.; McFarlane, A.C. Sleep disturbance immediately prior to trauma predicts subsequent psychiatric disorder. Sleep 2010, 33, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Gehrman, P.; Seelig, A.D.; Jacobson, I.G.; Boyko, E.J.; Hooper, T.I.; Gackstetter, G.D.; Ulmer, C.S.; Smith, T.C. Predeployment sleep duration and insomnia symptoms as risk factors for new-onset mental health disorders following military deployment. Sleep 2013, 36, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Koren, D.; Arnon, I.; Lavie, P.; Klein, E. Sleep complaints as early predictors of posttraumatic stress disorder: A 1-year prospective study of injured survivors of motor vehicle accidents. Am. J. Psychiatry 2002, 159, 855–857. [Google Scholar] [CrossRef] [Green Version]
- Lavie, P. Sleep disturbances in the wake of traumatic events. N. Engl. J. Med. 2001, 345, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Minkel, J.; Moreta, M.; Muto, J.; Htaik, O.; Jones, C.; Basner, M.; Dinges, D. Sleep deprivation potentiates HPA axis stress reactivity in healthy adults. Health Psychol. 2014, 33, 1430–1434. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.M.; Majde, J.A. Sleep as a host defense: Its regulation by microbial products and cytokines. Clin. Immunol. Immunopathol. 1990, 57, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Opp, M.R. Cytokines and sleep. Sleep Med. Rev. 2005, 9, 355–364. [Google Scholar] [CrossRef]
- Toth, L.A.; Tolley, E.A.; Krueger, J.M. Sleep as a prognostic indicator during infectious disease in rabbits. Proc. Soc. Exp. Biol. Med. 1993, 203, 179–192. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Opp, M.R.; Krueger, J.M. Sleep and immune function: Glial contributions and consequences of aging. Curr. Opin. Neurobiol. 2013, 23, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Poroyko, V.A.; Carreras, A.; Khalyfa, A.; Khalyfa, A.A.; Leone, V.; Peris, E.; Almendros, I.; Gileles-Hillel, A.; Qiao, Z.; Hubert, N.; et al. Chronic Sleep Disruption Alters Gut Microbiota, Induces Systemic and Adipose Tissue Inflammation and Insulin Resistance in Mice. Sci. Rep. 2016, 6, 35405. [Google Scholar] [CrossRef] [Green Version]
- Medina-Flores, F.; Hurtado-Alvarado, G.; Contis-Montes de Oca, A.; Lopez-Cervantes, S.P.; Konigsberg, M.; Deli, M.A.; Gomez-Gonzalez, B. Sleep loss disrupts pericyte-brain endothelial cell interactions impairing blood-brain barrier function. Brain Behav. Immun. 2020, 89, 118–132. [Google Scholar] [CrossRef]
- Hurtado-Alvarado, G.; Becerril-Villanueva, E.; Contis-Montes de Oca, A.; Dominguez-Salazar, E.; Salinas-Jazmin, N.; Perez-Tapia, S.M.; Pavon, L.; Velazquez-Moctezuma, J.; Gomez-Gonzalez, B. The yin/yang of inflammatory status: Blood-brain barrier regulation during sleep. Brain Behav. Immun. 2018, 69, 154–166. [Google Scholar] [CrossRef]
- Gomez-Gonzalez, B.; Hurtado-Alvarado, G.; Esqueda-Leon, E.; Santana-Miranda, R.; Rojas-Zamorano, J.A.; Velazquez-Moctezuma, J. REM sleep loss and recovery regulates blood-brain barrier function. Curr. Neurovasc. Res. 2013, 10, 197–207. [Google Scholar] [CrossRef]
- Hurtado-Alvarado, G.; Dominguez-Salazar, E.; Pavon, L.; Velazquez-Moctezuma, J.; Gomez-Gonzalez, B. Blood-Brain Barrier Disruption Induced by Chronic Sleep Loss: Low-Grade Inflammation May Be the Link. J. Immunol. Res. 2016, 2016, 4576012. [Google Scholar] [CrossRef] [Green Version]
- Lim, A.S.P.; Kowgier, M.; Yu, L.; Buchman, A.S.; Bennett, D.A. Sleep Fragmentation and the Risk of Incident Alzheimer’s Disease and Cognitive Decline in Older Persons. Sleep 2013, 36, 1027–1032. [Google Scholar] [CrossRef] [Green Version]
- Carskadon, M.A.; Brown, E.D.; Dement, W.C. Sleep fragmentation in the elderly: Relationship to daytime sleep tendency. Neurobiol. Aging 1982, 3, 321–327. [Google Scholar] [CrossRef]
- Ohayon, M.M.; Carskadon, M.A.; Guilleminault, C.; Vitiello, M.V. Meta-analysis of quantitative sleep parameters from childhood to old age in healthy individuals: Developing normative sleep values across the human lifespan. Sleep 2004, 27, 1255–1273. [Google Scholar] [CrossRef]
- Taillard, J.; Gronfier, C.; Bioulac, S.; Philip, P.; Sagaspe, P. Sleep in Normal Aging, Homeostatic and Circadian Regulation and Vulnerability to Sleep Deprivation. Brain Sci. 2021, 11, 1003. [Google Scholar] [CrossRef]
- Zepelin, H.; Whitehead, W.E.; Rechtschaffen, A. Aging and sleep in the albino rat. Behav. Biol. 1972, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Witting, W.; Mirmiran, M.; Bos, N.P.; Swaab, D.F. Effect of light intensity on diurnal sleep-wake distribution in young and old rats. Brain Res. Bull. 1993, 30, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Mendelson, W.B.; Bergmann, B.M. Age-related changes in sleep in the rat. Sleep 1999, 22, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, R.S.; Zepelin, H.; Rechtschaffen, A. Sleep in young and old rats. J. Gerontol. 1979, 34, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Borbely, A.A. Sleep regulation. Introduction. Hum. Neurobiol. 1982, 1, 161–162. [Google Scholar]
- Pappenheimer, J.R.; Koski, G.; Fencl, V.; Karnovsky, M.L.; Krueger, J. Extraction of sleep-promoting factor S from cerebrospinal fluid and from brains of sleep-deprived animals. J. Neurophysiol. 1975, 38, 1299–1311. [Google Scholar] [CrossRef]
- Borbely, A.A.; Tobler, I.; Hanagasioglu, M. Effect of sleep deprivation on sleep and EEG power spectra in the rat. Behav. Brain Res. 1984, 14, 171–182. [Google Scholar] [CrossRef]
- Freedman, R.R. EEG power spectra in sleep-onset insomnia. Electroencephalogr. Clin. Neurophysiol. 1986, 63, 408–413. [Google Scholar] [CrossRef]
- Perlis, M.L.; Smith, M.T.; Andrews, P.J.; Orff, H.; Giles, D.E. Beta/Gamma EEG activity in patients with primary and secondary insomnia and good sleeper controls. Sleep 2001, 24, 110–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krystal, A.D.; Edinger, J.D.; Wohlgemuth, W.K.; Marsh, G.R. NREM sleep EEG frequency spectral correlates of sleep complaints in primary insomnia subtypes. Sleep 2002, 25, 630–640. [Google Scholar] [PubMed] [Green Version]
- Britten, R.A.; Hienz, R.D.; Davis, C.M.; Yoon, H.; Machida, M.; Wellman, L.L.; Sanford, L.D. Hadron-induced impairment of Executive Function: Role of perturbed neurotransmission and the exacerbating impact of sleep deprivation. Grant application to NASA Specialized Centers of Research (NSCORs) for Ground-Based Studies in Cancer Risks and Cognitive and Behavioral Central Nervous System Risks from Space Radiation. 2014; unpublished data. [Google Scholar]
- Buzsaki, G.; Leung, L.W.; Vanderwolf, C.H. Cellular bases of hippocampal EEG in the behaving rat. Brain Res. 1983, 287, 139–171. [Google Scholar] [CrossRef] [PubMed]
- Klemm, W.R.; Naugle, N.W. Oscillatory electrographic activity in the hippocampus: A mathematical model. Neurosci. Biobehav. Rev. 1980, 4, 437–449. [Google Scholar] [CrossRef]
- Jouvet, M. Biogenic amines and the states of sleep. Science 1969, 163, 32–41. [Google Scholar] [CrossRef]
- Buzsaki, G. The hippocampo-neocortical dialogue. Cereb. Cortex 1996, 6, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Buzsaki, G. Theta oscillations in the hippocampus. Neuron 2002, 33, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Boyce, R.; Glasgow, S.D.; Williams, S.; Adamantidis, A. Causal evidence for the role of REM sleep theta rhythm in contextual memory consolidation. Science 2016, 352, 812–816. [Google Scholar] [CrossRef]
- Lesting, J.; Narayanan, R.T.; Kluge, C.; Sangha, S.; Seidenbecher, T.; Pape, H.C. Patterns of coupled theta activity in amygdala-hippocampal-prefrontal cortical circuits during fear extinction. PLoS ONE 2011, 6, e21714. [Google Scholar] [CrossRef] [Green Version]
- Hancock, P.A.; Ross, J.M.; Szalma, J.L. A meta-analysis of performance response under thermal stressors. Hum. Factors 2007, 49, 851–877. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.; Watkins, S.L.; Marshall, H.; Dascombe, B.J.; Foster, J. The Impact of Different Environmental Conditions on Cognitive Function: A Focused Review. Front. Physiol. 2015, 6, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymusiak, R. Body temperature and sleep. Handb. Clin. Neurol. 2018, 156, 341–351. [Google Scholar] [CrossRef]
- McGinty, D.; Szymusiak, R.; Thomson, D. Preoptic/anterior hypothalamic warming increases EEG delta frequency activity within non-rapid eye movement sleep. Brain Res. 1994, 667, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; McGinty, D.; Szymusiak, R. Preoptic/anterior hypothalamic neurons: Thermosensitivity in wakefulness and non rapid eye movement sleep. Brain Res. 1996, 718, 76–82. [Google Scholar] [CrossRef]
- Laiakis, E.C.; Pinheiro, M.; Nguyen, T.; Nguyen, H.; Beheshti, A.; Dutta, S.M.; Russell, W.K.; Emmett, M.R.; Britten, R.A. Quantitative proteomic analytic approaches to identify metabolic changes in the medial prefrontal cortex of rats exposed to space radiation. Front. Physiol. 2022, 13, 971282. [Google Scholar] [CrossRef] [PubMed]
- Soler, I.; Yun, S.; Reynolds, R.P.; Whoolery, C.W.; Tran, F.H.; Kumar, P.L.; Rong, Y.; DeSalle, M.J.; Gibson, A.D.; Stowe, A.M.; et al. Multi-Domain Touchscreen-Based Cognitive Assessment of C57BL/6J Female Mice Shows Whole-Body Exposure to (56)Fe Particle Space Radiation in Maturity Improves Discrimination Learning Yet Impairs Stimulus-Response Rule-Based Habit Learning. Front. Behav. Neurosci. 2021, 15, 722780. [Google Scholar] [CrossRef]
- Rabin, B.M.; Miller, M.G.; Larsen, A.; Spadafora, C.; Zolnerowich, N.N.; Dell’Acqua, L.A.; Shukitt-Hale, B. Effects of exposure to (12)C and (4)He particles on cognitive performance of intact and ovariectomized female rats. Life Sci. Space Res. 2019, 22, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Krukowski, K.; Grue, K.; Becker, M.; Elizarraras, E.; Frias, E.S.; Halvorsen, A.; Koenig-Zanoff, M.; Frattini, V.; Nimmagadda, H.; Feng, X.; et al. The impact of deep space radiation on cognitive performance: From biological sex to biomarkers to countermeasures. Sci. Adv. 2021, 7, eabg6702. [Google Scholar] [CrossRef] [PubMed]
- Krukowski, K.; Feng, X.; Paladini, M.S.; Chou, A.; Sacramento, K.; Grue, K.; Riparip, L.K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; et al. Temporary microglia-depletion after cosmic radiation modifies phagocytic activity and prevents cognitive deficits. Sci. Rep. 2018, 8, 7857. [Google Scholar] [CrossRef] [PubMed]
- Kiffer, F.; Alexander, T.; Anderson, J.E.; Groves, T.; Wang, J.; Sridharan, V.; Boerma, M.; Allen, A.R. Late Effects of (16)O-Particle Radiation on Female Social and Cognitive Behavior and Hippocampal Physiology. Radiat. Res. 2019, 191, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Fesshaye, A.; Ihle, P.; Wheeler, A.; Baulch, J.E.; Limoli, C.L.; Stark, C.E. Dissecting Differential Complex Behavioral Responses to Simulated Space Radiation Exposures. Radiat. Res. 2022, 197, 289–297. [Google Scholar] [CrossRef]
- Sweeten, B.L.W.; Sutton, A.M.; Wellman, L.L.; Sanford, L.D. Predicting stress resilience and vulnerability: Brain-derived neurotrophic factor and rapid eye movement sleep as potential biomarkers of individual stress responses. Sleep 2020, 43, zsz199. [Google Scholar] [CrossRef]
- Wellman, L.L.; Fitzpatrick, M.E.; Hallum, O.Y.; Sutton, A.M.; Williams, B.L.; Sanford, L.D. Individual Differences in Animal Stress Models: Considering Resilience, Vulnerability, and the Amygdala in Mediating the Effects of Stress and Conditioned Fear on Sleep. Sleep 2016, 39, 1293–1303. [Google Scholar] [CrossRef] [Green Version]
- Wellman, L.L.; Fitzpatrick, M.E.; Hallum, O.Y.; Sutton, A.M.; Williams, B.L.; Sanford, L.D. The basolateral amygdala can mediate the effects of fear memory on sleep independently of fear behavior and the peripheral stress response. Neurobiol. Learn. Mem. 2017, 137, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Wellman, L.L.; Fitzpatrick, M.E.; Sutton, A.M.; Williams, B.L.; Machida, M.; Sanford, L.D. Antagonism of corticotropin releasing factor in the basolateral amygdala of resilient and vulnerable rats: Effects on fear-conditioned sleep, temperature and freezing. Horm. Behav. 2018, 100, 20–28. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanford, L.D.; Adkins, A.M.; Boden, A.F.; Gotthold, J.D.; Harris, R.D.; Shuboni-Mulligan, D.; Wellman, L.L.; Britten, R.A. Sleep and Core Body Temperature Alterations Induced by Space Radiation in Rats. Life 2023, 13, 1002. https://doi.org/10.3390/life13041002
Sanford LD, Adkins AM, Boden AF, Gotthold JD, Harris RD, Shuboni-Mulligan D, Wellman LL, Britten RA. Sleep and Core Body Temperature Alterations Induced by Space Radiation in Rats. Life. 2023; 13(4):1002. https://doi.org/10.3390/life13041002
Chicago/Turabian StyleSanford, Larry D., Austin M. Adkins, Alea F. Boden, Justin D. Gotthold, Ryan D. Harris, Dorela Shuboni-Mulligan, Laurie L. Wellman, and Richard A. Britten. 2023. "Sleep and Core Body Temperature Alterations Induced by Space Radiation in Rats" Life 13, no. 4: 1002. https://doi.org/10.3390/life13041002
APA StyleSanford, L. D., Adkins, A. M., Boden, A. F., Gotthold, J. D., Harris, R. D., Shuboni-Mulligan, D., Wellman, L. L., & Britten, R. A. (2023). Sleep and Core Body Temperature Alterations Induced by Space Radiation in Rats. Life, 13(4), 1002. https://doi.org/10.3390/life13041002