

Physical Concept to Explain the Regulation of Lipid Membrane Phase Separation under Isothermal Conditions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Free Energy of Phase Separation
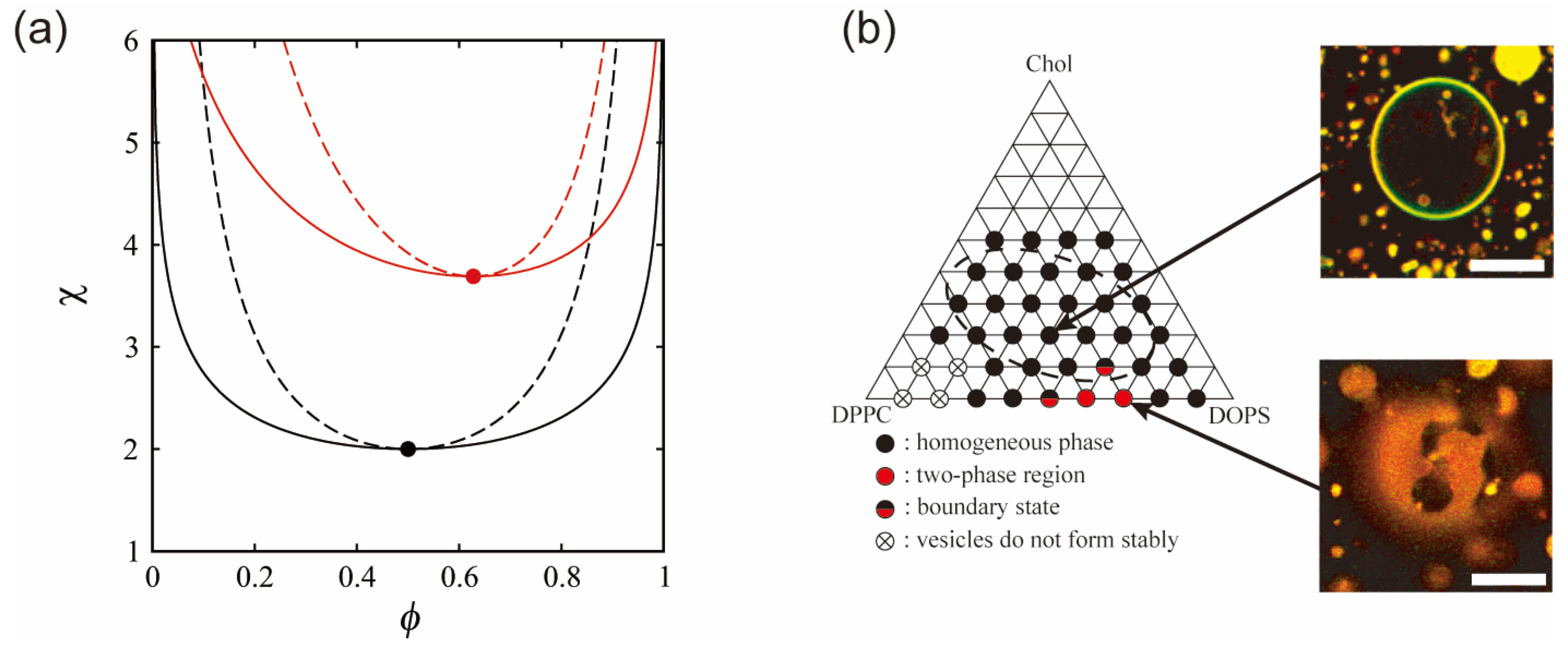
3. Phase Separation in Charged Membranes
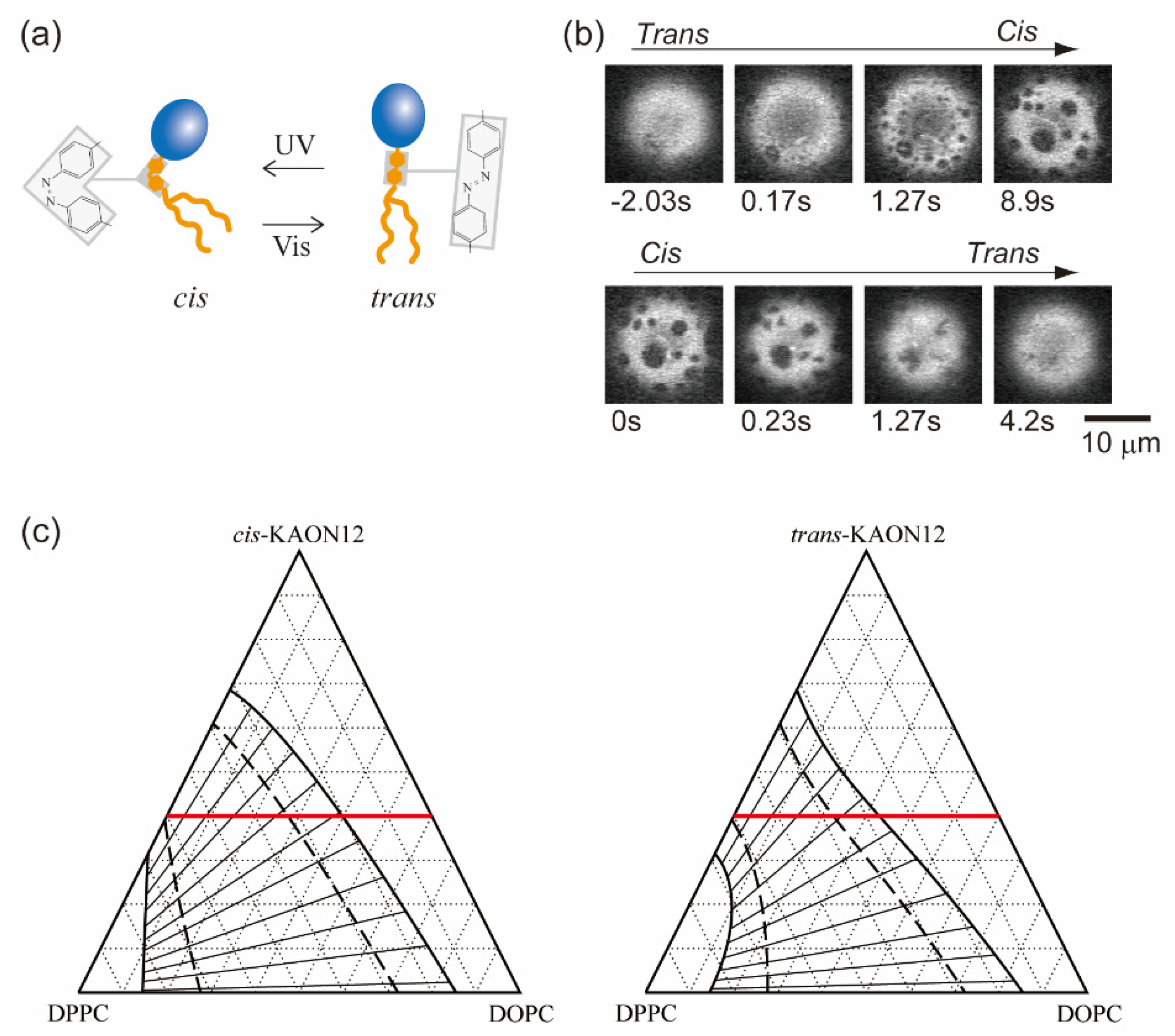
4. Phase Separation Induced by Chemical Reactions within a Bilayer
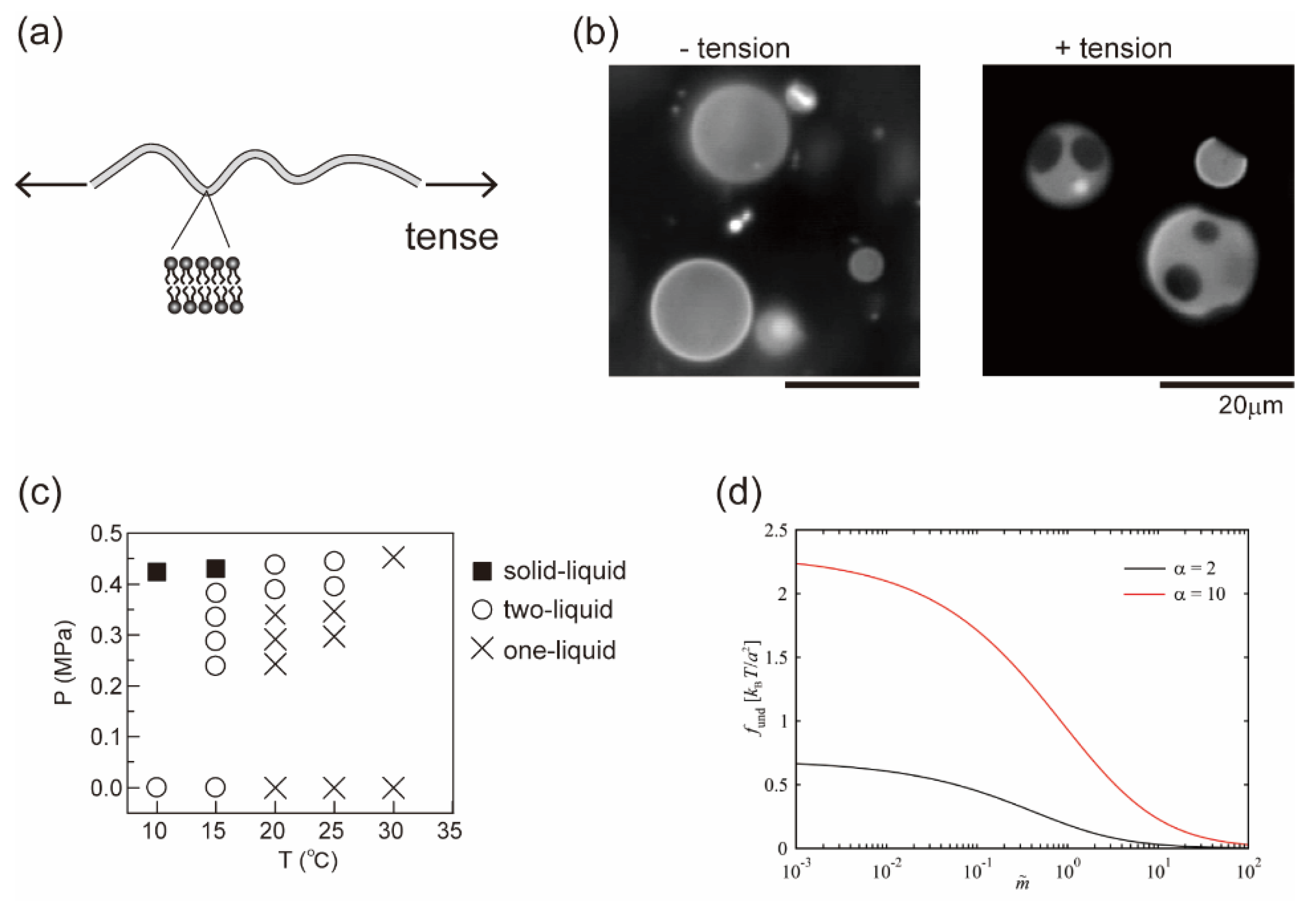
5. Effect of Membrane Tension on Phase Separation

6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberts, B.; Johnson, A.; Walter, P.; Lewis, J.; Raff, M. Molecular Biology of the Cell; Garland Science: New York, NY, USA, 2008. [Google Scholar]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.F. Lipid rafts: Contentious only from simplistic standpoints. Nat. Rev. Mol. Cell Biol. 2006, 7, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Sampaion, J.L. Membrane Organization and Lipid Rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; London, E. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 2000, 275, 17221–17224. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Yadav, V.; Saini, N. Lipid rafts in immune signalling: Current progress and future perspective. Immunology 2016, 149, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-T.; Kiessling, V.; Yamm, L.K. Line tension at lipid phase boundaries as driving force for HIV fusion peptide-mediated fusion. Nat. Commun. 2016, 7, 11401. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, Z.; Zhang, L.; Shi, L.; Shahriar, S.; Chan, R.B.; Paolo, G.D.; Min, W. Metabolic activity induces membrane phase separation in endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2017, 114, 13394–13399. [Google Scholar] [CrossRef]
- King, C.; Sengupta, P.; Seo, A.Y.; Lippincott-Schwartz, J. ER membranes exhibit phase behavior at sites of organelle contact. Proc. Natl. Acad. Sci. USA 2020, 117, 7225–7235. [Google Scholar] [CrossRef]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant Vesicles: Preparations and Applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef]
- Hamada, T.; Yoshikawa, K. Cell-Sized Liposomes and Droplets: Real-World Modeling of Living Cells. Materials 2012, 5, 2292–2305. [Google Scholar] [CrossRef]
- Dimova, R. Giant Vesicles and Their Use in Assays for Assessing Membrane Phase State, Curvature, Mechanics, and Electrical Properties. Annu. Rev. Phys. 2019, 48, 93–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Du, H.; Wang, Z.; Mu, W.; Han, X. Versatile Phospholipid Assemblies for Functional Synthetic Cells and Artificial Tissues. Adv. Mat. 2021, 33, 2002635. [Google Scholar] [CrossRef] [PubMed]
- Miele, Y.; Holló, G.; Lagzi, I.; Rossi, F. Shape Deformation, Budding and Division of Giant Vesicles and Artificial Cells: A Review. Life 2022, 12, 841. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nagai, K.H.; Zinchenko, A.; Hamada, T. Photoinduced Fusion of Lipid Bilayer Membranes. Langmuir 2017, 33, 2671–2676. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C.; Bagatolli, L.A.; Volovyk, Z.N.; Thompson, N.L.; Levi, M.; Jacobson, K.; Gratton, E. Lipid Rafts Reconstituted in Model Membranes. Biophys. J. 2001, 80, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Korlach, J.; Schwille, P.; Webb, W.W.; Feigenson, G.W. Characterization of lipid bilayer phases by confocal microscopy and fluorescence correlation spectroscopy. Proc. Natl. Acad. Sci. USA 1999, 96, 8461–8466. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, T.; Hess, S.T.; Webb, W.W. Imaging coexisting fluid domains in biomembrane models coupling curvature and line tension. Nature 2003, 425, 821–824. [Google Scholar] [CrossRef]
- Baumgart, T.; Hammond, A.T.; Sengupta, P.; Webb, W.W. Large-scale fluid/fluid phase separation of proteins and lipids in giant plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2007, 104, 3165–3170. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Morita, M.; Miyakawa, M.; Sugimoto, R.; Hatanaka, A.; Vestergaard, M.C.; Takagi, M. Size-dependent partitioning of nano/microparticles mediated by membrane lateral heterogeneity. J. Am. Chem. Soc. 2012, 134, 13990–13996. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.L. Soft Condensed Matter; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Veatch, S.L.; Keller, S.L. Separation of Liquid Phases in Giant Vesicles of Ternary Mixtures of Phospholipids and Cholesterol. Biophys. J. 2003, 85, 3074–3083. [Google Scholar] [CrossRef]
- Veatch, S.L.; Keller, S.L. Organization in Lipid Membranes Containing Cholesterol. Phys. Rev. Lett. 2002, 89, 268101. [Google Scholar] [CrossRef] [PubMed]
- Veatch, S.L.; Keller, S.L. Miscibility Phase Diagrams of Giant Vesicles Containing Sphingomyelin. Phys. Rev. Lett. 2005, 94, 148101. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Stinson, B.M.; Go, M.S.; Carmona, L.M.; Reminick, J.I.; Fang, X.; Baumgart, T. Temperature-dependent phase behavior and protein partitioning in giant plasma membrane vesicles. Biochim. Biophys. Acta 2010, 1798, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Miura, Y.; Ishii, K.; Araki, S.; Yoshikawa, K.; Vestergaard, M.; Takagi, M. Dynamic Processes in Endocytic Transformation of a Raft-Exhibiting Giant Liposome. J. Phys. Chem. B 2007, 111, 10853–10857. [Google Scholar] [CrossRef] [PubMed]
- Flory, P.J. Thermodynamics of High Polymer Solutions. J. Chem. Phys. 1942, 10, 51. [Google Scholar] [CrossRef]
- Huggins, M.L. Some Properties of Solutions of Long-chain Compounds. J. Phys. Chem. 1942, 46, 151–158. [Google Scholar] [CrossRef]
- Wagner, J.; Loew, S.; May, S. Influence of monolayer-monolayer coupling on the phase behavior of a fluid lipid bilayer. Biophys. J. 2007, 93, 4268–4277. [Google Scholar] [CrossRef]
- Shimokawa, N.; Hishida, M.; Seto, H.; Yoshikawa, K. Phase separation of a mixture of charged and neutral lipids on a giant vesicle induced by small cations. Chem. Phys. Lett. 2010, 496, 59–63. [Google Scholar] [CrossRef]
- Komura, S.; Andelman, D. Physical aspects of heterogeneities in multi-component lipid membranes. Adv. Colloid Interface Sci. 2014, 208, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Komura, S.; Shirotori, H.; Olmsted, P.D.; Andelman, D. Lateral phase separation in mixtures of lipids and cholesterol. EPL 2004, 67, 321–327. [Google Scholar] [CrossRef]
- Komura, S.; Shirotori, H.; Olmsted, P.D. Phase behaviour of three-component lipid mixtures. J. Phys. Condens. Matter 2005, 17, S2951–S2956. [Google Scholar] [CrossRef]
- Shimokawa, N.; Komura, S.; Andelman, D. The phase behavior of mixed lipid membranes in the presence of the rippled phase. Eur. Phys. J. E 2008, 26, 197–204. [Google Scholar] [CrossRef]
- Shimokawa, N.; Himeno, H.; Hamada, T.; Takagi, M.; Komura, S.; Andelman, D. Phase Diagrams and Ordering in Charged Membranes: Binary Mixtures of Charged and Neutral Lipids. J. Phys. Chem. B 2016, 120, 6358–6367. [Google Scholar] [CrossRef] [PubMed]
- Vist, M.R.; Davis, J.H. Phase equilibria of cholesterol/dipalmitoylphosphatidylcholine mixtures: Deuterium nuclear magnetic resonance and differential scanning calorimetry. Biochemistry 1990, 29, 451–464. [Google Scholar] [CrossRef]
- Putzel, G.G.; Schick, M. Phase Behavior of a Model Bilayer Membrane with Coupled Leaves. Biophys. J. 2008, 94, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Rozovsky, S.; Kaizuka, Y.; Groves, J.T. Formation and Spatio-Temporal Evolution of Periodic Structures in Lipid Bilayers. J. Am. Chem. Soc. 2005, 127, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Shimokawa, N.; Ichikawa, M.; Yoshikawa, K. Micro-segregation induced by bulky-head lipids: Formation of characteristic patterns in a giant vesicle. Soft Matter 2012, 8, 488–495. [Google Scholar] [CrossRef]
- Shimobayashi, S.F.; Ichikawa, M.; Taniguchi, T. Direct observations of transition dynamics from macro- to micro-phase separation in asymmetric lipid bilayers induced by externally added glycolipids. EPL 2016, 113, 56005. [Google Scholar] [CrossRef]
- Shimokawa, N.; Mukai, R.; Nagata, M.; Takagi, M. Formation of modulated phases and domain rigidification in fatty acid-containing lipid membranes. Phys. Chem. Chem. Phys. 2017, 19, 13252–13263. [Google Scholar] [CrossRef]
- Leibler, S.; Andelman, D. Ordered and curved meso-structures in membranes and amphiphilic films. J. Phys. Fr. 1987, 48, 2013–2018. [Google Scholar] [CrossRef]
- Komura, S.; Shimokawa, N.; Andelman, D. Tension-induced morphological transition in mixed lipid bilayers. Langmuir 2006, 22, 6771–6774. [Google Scholar] [CrossRef] [PubMed]
- Hirose, Y.; Komura, S.; Andelman, D. Coupled Modulated Bilayers: A Phenomenological Model. Chem. Phys. Chem. 2009, 10, 2839–2846. [Google Scholar] [CrossRef] [PubMed]
- Brewster, R.; Safran, A. Line Active Hybrid Lipids Determine Domain Size in Phase Separation of Saturated and Unsaturated Lipids. Biophys. J. 2010, 98, L21–L23. [Google Scholar] [CrossRef] [PubMed]
- Scheve, C.S.; Gonzales, P.A.; Momin, N.; Stachowiak, J.C. Steric Pressure between Membrane-Bound Proteins Opposes Lipid Phase Separation. J. Am. Chem. Soc. 2013, 135, 1185–1188. [Google Scholar] [CrossRef]
- Vequi-Suplicy, C.C.; Riske, K.A.; Knorr, R.L.; Dimova, R. Vesicles with charged domains. Biochim. Biophys. Acta 2010, 1798, 1338–1347. [Google Scholar] [CrossRef]
- Blosser, M.C.; Starr, J.B.; Turtle, C.W.; Ashcraft, J.; Keller, S.L. Minimal effect of lipid charge on membrane miscibility phase behavior in three ternary systems. Biophys. J. 2013, 104, 2629–2638. [Google Scholar] [CrossRef]
- Himeno, H.; Shimokawa, N.; Komura, S.; Andelman, D.; Hamada, T.; Takagi, M. Charge-induced phase separation in lipid membranes. Soft Matter 2014, 10, 7959–7967. [Google Scholar] [CrossRef]
- May, S.; Harries, D.; Ben-Shaul, A. Macroion-Induced Compositional Instability of Binary Fluid Membranes. Phys. Rev. Lett. 2002, 89, 268102. [Google Scholar] [CrossRef]
- Harries, D.; May, S.; Ben-Shaul, A. Adsorption of charged macromolecules on mixed fluid membranes. Colloids Surf. A 2002, 208, 41–50. [Google Scholar] [CrossRef]
- Mbamala, E.C.; Ben-Shaul, A.; May, S. Domain Formation Induced by the Adsorption of Charged Proteins on Mixed Lipid Membranes. Biophys. J. 2005, 88, 1702–1714. [Google Scholar] [CrossRef]
- Baciu, C.L.; May, S. Stability of charged, mixed lipid bilayers: Effect of electrostatic coupling between the monolayers. J. Phys. Condens. Matter 2004, 16, S2455. [Google Scholar] [CrossRef]
- Wagner, A.J.; May, S. Electrostatic interactions across a charged lipid bilayer. Eur. Biophys. J. 2007, 36, 293–303. [Google Scholar] [CrossRef]
- Shimokawa, N.; Komura, S.; Andelman, D. Charged bilayer membranes in asymmetric ionic solutions: Phase diagrams and critical behavior. Phys. Rev. E 2011, 84, 031919. [Google Scholar] [CrossRef] [PubMed]
- Verwey, E.J.W.; Overbeek, J.T.G. Theory of the Stability of Lyophobic Colloids; Elsevier: Amsterdam, The Netherlands, 1948. [Google Scholar]
- Evans, D.F.; Wennerström, H. The Colloidal Domain; John Wiley: New York, NY, USA, 1999. [Google Scholar]
- Israelachvili, J. Intermolecular and Surface Forces; University of California-Santa Barbara: Santa Barbara, CA, USA, 2011. [Google Scholar]
- Kubsch, B.; Robinson, T.; Lipowsky, R.; Dimova, R. Solution Asymmetry and Salt Expand Fluid-Fluid Coexistence Regions of Charged Membranes. Biophys. J. 2016, 110, 2581–2584. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Ito, H.; Higuchi, Y.; Bohinc, K.; Shimokawa, N.; Takagi, M. Three-Phase Coexistence in Binary Charged Lipid Membranes in a Hypotonic Solution. Langmuir 2021, 37, 9683–9693. [Google Scholar] [CrossRef]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef]
- Hamada, T.; Sugimoto, R.; Nagasaki, T.; Takagi, M. Photochemical control of membrane raft organization. Soft Matter 2011, 7, 220–224. [Google Scholar] [CrossRef]
- Hamada, T.; Sato, Y.T.; Nagasaki, T.; Yoshikawa, K. Reversible photoswitching in a cell-sized vesicle. Langmuir 2005, 21, 7626–7628. [Google Scholar] [CrossRef]
- Ishii, K.; Hamada, T.; Hatakeyama, M.; Sugimoto, R.; Nagasaki, T.; Takagi, M. Reversible Control of Exo- and Endo-Budding Transitions in a Photosensitive Lipid Membrane. ChemBioChem 2009, 10, 251–256. [Google Scholar] [CrossRef]
- Hamada, T.; Sugimoto, R.; Vestergaard, M.; Nagasaki, T.; Takagi, M. Membrane disc and sphere: Controllable mesoscopic structures for the capture and release of a targeted object. J. Am. Chem. Soc. 2010, 132, 10528–10532. [Google Scholar] [CrossRef]
- Yasuhara, K.; Sasaki, Y.; Kikuchi, J. A photo-responsive cholesterol capable of inducing a morphological transformation of the liquid-ordered microdomain in lipid bilayers. Colloid Polym. Sci. 2008, 286, 1675–1680. [Google Scholar] [CrossRef]
- Urban, P.; Pritzl, S.D.; Konrad, D.B.; Frank, J.A.; Pernpeintner, C.; Roeske, C.R.; Trauner, D.; Lohmuller, T. Light-Controlled Lipid Interaction and Membrane Organization in Photolipid Bilayer Vesicles. Langmuir 2018, 34, 13368–13374. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Franquelim, H.G.; Schwille, P.; Trauner, D. Optical Control of Lipid Rafts with Photoswitchable Ceramides. J. Am. Chem. Soc. 2016, 138, 12981–12986. [Google Scholar] [CrossRef] [PubMed]
- Tsubone, T.M.; Baptista, M.S.; Itri, R. Understanding membrane remodelling initiated by photosensitized lipid oxidation. Biophys. Chem. 2019, 254, 106263. [Google Scholar] [CrossRef]
- Yoda, T.; Vestergaard, M.C.; Akazawa-Ogawa, Y.; Yoshida, Y.; Hamada, T.; Takagi, M. Dynamic response of a cholesterol-containing model membrane to oxidative stress. Chem. Lett. 2010, 39, 1273–1274. [Google Scholar] [CrossRef]
- Meijering, J.L. Segregation in regular ternary solutions PART I. Philips Res. Rep. 1950, 5, 333–356. [Google Scholar]
- Knobler, C.M.; Scott, R.L. Phase Transitions and Critical Phenomena; Domb, C., Lebowitz, J.L., Eds.; Academic Press: New York, NY, USA, 1984; Volume 9, p. 164. [Google Scholar]
- Hammond, A.T.; Heberle, F.A.; Baumgart, T.; Holowka, D.; Baird, B.; Feigenson, G.W. Crosslinking a lipid raft component triggers liquid ordered-liquid disordered phase separation in model plasma membranes. Proc. Natl. Acad. Sci. USA 2005, 102, 6320–6325. [Google Scholar] [CrossRef]
- Roffay, C.; Molinard, G.; Kim, K.; Urbanska, M.; Andrade, V.; Barbarasa, V.; Nowak, P.; Mercier, V.; García-Calvo, J.; Matile, S.; et al. Passive coupling of membrane tension and cell volume during active response of cells to osmosis. Proc. Natl. Acad. Sci. USA 2021, 118, e2103228118. [Google Scholar] [CrossRef]
- Colom, A.; Derivery, E.; Soleimanpour, S.; Tomba, C.; Molin, M.D.; Sakai, N.; González-Gaitán, M.; Matile, S.; Roux, A. A fluorescent membrane tension probe. Nat. Chem. 2018, 10, 1118–1125. [Google Scholar] [CrossRef]
- Sachs, F. Stretch-Activated Ion Channels: What Are They? Physiology 2010, 25, 50–56. [Google Scholar] [CrossRef]
- Sukharev, S.I.; Blount, P.; Martinac, B.; Blattner, F.R.; Kung, C. A large-conductance mechanosensitive channel in E. coli encoded by mscL alone. Nature 1994, 368, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ando, J. Vascular endothelial cell membranes differentiate between stretch and shear stress through transitions in their lipid phases. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1178–H1185. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Kishimoto, Y.; Nagasaki, T.; Takagi, M. Lateral phase separation in tense membranes. Soft Matter 2011, 7, 9061–9068. [Google Scholar] [CrossRef]
- Rathe, V.; Kuckla, D.; Monzel, C. Phase separation in biological membranes: An overview with focus on experimental effects of illumination and osmotic pressure changes. Adv. Biomembr. Lipid Self-Assem. 2021, 34, 31–66. [Google Scholar]
- Li, L.; Cheng, J.-X. Coexisting Stripe and Patch Shaped Domains in Giant Unilamellar Vesicles. Biochemistry 2006, 45, 11819–11826. [Google Scholar] [CrossRef]
- Ayuyan, A.G.; Cohen, F.S. Raft Composition at Physiological Temperature and pH in the Absence of Detergents. Biophys. J. 2008, 94, 2654–2666. [Google Scholar] [CrossRef] [PubMed]
- Oglȩcka, K.; Rangamani, P.; Liedberg, B.; Kraut, R.S.; Parikh, A.N. Oscillatory phase separation in giant lipid vesicles induced by transmembrane osmotic differentials. eLife 2014, 3, e03695. [Google Scholar] [CrossRef]
- Knop, J.M.; Mukherjee, S.; Jaworek, M.W.; Kriegler, S.; Manisegaran, M.; Fetahaj, Z.; Ostermeier, L.; Oliva, R.; Gault, S.; Cockell, C.S.; et al. Life in Multi-Extreme Environments: Brines, Osmotic and Hydrostatic Pressure—A Physicochemical View. Chem. Rev. 2023, 123, 73–104. [Google Scholar] [CrossRef]
- Helfrich, W.Z. Steric Interaction of Fluid Membranes in Multilayer Systems. Naturforsch 1978, 33, 305–315. [Google Scholar] [CrossRef]
- Gordon, V.D.; Deserno, M.; Andrew, C.M.J.; Egelhaaf, S.U.; Poon, W.C.K. Adhesion promotes phase separation in mixed-lipid membranes. EPL 2008, 84, 48003. [Google Scholar] [CrossRef]
- Wongsirojkul, N.; Shimokawa, N.; Opaprakasit, P.; Takagi, M.; Hamada, T. Osmotic-Tension-Induced Membrane Lateral Organization. Langmuir 2020, 36, 2937–2945. [Google Scholar] [CrossRef] [PubMed]
- Wongsirojkul, N.; Masuta, A.; Shimokawa, N.; Takagi, M. Control of Line Tension at Phase-Separated Lipid Domain Boundaries: Monounsaturated Fatty Acids with Different Chain Lengths and Osmotic Pressure. Membranes 2022, 12, 781. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.; Dittrich, P.S. Observations of Membrane Domain Reorganization in Mechanically Compressed Artificial Cells. ChemBioChem 2019, 20, 2666–2673. [Google Scholar] [CrossRef] [PubMed]
- Sturzenegger, F.; Robinson, T.; Hessa, D.; Dittrich, P.S. Membranes under shear stress: Visualization of non-equilibrium domain patterns and domain fusion in a microfluidic device. Soft Matter 2016, 12, 5072–5076. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Mizuno, S.; Kitahata, H. Domain dynamics of phase-separated lipid membranes under shear flow. Soft Matter 2022, 18, 9069–9075. [Google Scholar] [CrossRef] [PubMed]
- Veatch, S.L.; Rogers, N.; Decker, A.; Shelby, S.A. The plasma membrane as an adaptable fluid mosaic. Biochim. Biophys. Acta (BBA)-Biomembr. 2023, 1865, 184114. [Google Scholar] [CrossRef]
- Goh, M.W.S.; Tero, R. Non-raft submicron domain formation in cholesterol-containing lipid bilayers induced by polyunsaturated phosphatidylethanolamine. Colloids Surf. B Biointerfaces 2022, 210, 112235. [Google Scholar] [CrossRef]
- Hirschi, S.; Ward, T.R.; Meier, W.P.; Müller, D.J.; Fotiadis, D. Synthetic Biology: Bottom-Up Assembly of Molecular Systems. Chem. Rev. 2022, 122, 16294–16328. [Google Scholar] [CrossRef]
- Schwille, P.; Spatz, J.; Landfester, K.; Bodenschatz, E.; Herminghaus, S.; Sourjik, V.; Erb, T.J.; Bastiaens, P.; Lipowsky, R.; Hyman, A.; et al. MaxSynBio: Avenues Towards Creating Cells from the Bottom Up. Angew. Chem. 2018, 57, 13382–13392. [Google Scholar] [CrossRef]
- Stano, P. Minimal cells: Relevance and interplay of physical and biochemical factors. Biotech. J. 2011, 6, 850–859. [Google Scholar] [CrossRef]
- Imai, M.; Sakuma, Y.; Kurisu, M.; Walde, P. From vesicles toward protocells and minimal cells. Soft Matter 2022, 18, 4823–4849. [Google Scholar] [CrossRef]
- Lipowsky, R. Remodeling of Membrane Shape and Topology by Curvature Elasticity and Membrane Tension. Adv. Biol. 2022, 6, 102613. [Google Scholar] [CrossRef]
- Hamada, T.; Fujimoto, R.; Shimobayashi, S.F.; Ichikawa, M.; Takagi, M. Molecular behavior of DNA in a cell-sized compartment coated by lipids. Phys. Rev. E 2015, 91, 062717. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hiratsuka, Y.; Kawamata, I.; Murata, S.; Nomura, S.-I.M. Micrometer-sized molecular robot changes its shape in response to signal molecules. Sci. Robot. 2017, 2, eaal3735. [Google Scholar] [CrossRef] [PubMed]
- Takiue, T. Heterogeneity and deformation behavior of lipid vesicles. Curr. Opin. Colloid Interface Sci. 2022, 62, 101646. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimokawa, N.; Hamada, T. Physical Concept to Explain the Regulation of Lipid Membrane Phase Separation under Isothermal Conditions. Life 2023, 13, 1105. https://doi.org/10.3390/life13051105
Shimokawa N, Hamada T. Physical Concept to Explain the Regulation of Lipid Membrane Phase Separation under Isothermal Conditions. Life. 2023; 13(5):1105. https://doi.org/10.3390/life13051105
Chicago/Turabian StyleShimokawa, Naofumi, and Tsutomu Hamada. 2023. "Physical Concept to Explain the Regulation of Lipid Membrane Phase Separation under Isothermal Conditions" Life 13, no. 5: 1105. https://doi.org/10.3390/life13051105
APA StyleShimokawa, N., & Hamada, T. (2023). Physical Concept to Explain the Regulation of Lipid Membrane Phase Separation under Isothermal Conditions. Life, 13(5), 1105. https://doi.org/10.3390/life13051105