

The Polymorphisms in GSTO Genes (GSTO1 rs4925, GSTO2 rs156697, and GSTO2 rs2297235) Affect the Risk for Testicular Germ Cell Tumor Development: A Pilot Study

 ,
,  , ,
, ,  ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethical Approval
2.3. GSTO Genotyping
2.4. Statistical and Haplotype Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oosterhuis, J.W.; Looijenga, L.H.J. Testicular Germ-Cell Tumours in a Broader Perspective. Nat. Rev. Cancer 2005, 5, 210–222. [Google Scholar] [CrossRef]
- Winter, C.; Albers, P. Testicular Germ Cell Tumors: Pathogenesis, Diagnosis and Treatment. Nat. Rev. Endocrinol. 2011, 7, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.J.; Turner, M.C.; Ghadirian, P.; Krewski, D. Epidemiology of Testicular Cancer: An Overview. Int. J. Cancer 2005, 116, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Katabathina, V.S.; Vargas-Zapata, D.; Monge, R.A.; Nazarullah, A.; Ganeshan, D.; Tammisetti, V.; Prasad, S.R. Testicular Germ Cell Tumors: Classification, Pathologic Features, Imaging Findings, and Management. Radiographics 2021, 41, 1698–1716. [Google Scholar] [CrossRef]
- Looijenga, L.H.J.; Van Agthoven, T.; Biermann, K. Development of Malignant Germ Cells—The Genvironmental Hypothesis. Int. J. Dev. Biol. 2013, 57, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Skakkebæk, N.E.; Rajpert-De Meyts, E.; Main, K.M. Testicular Dysgenesis Syndrome: An Increasingly Common Developmental Disorder with Environmental Aspects: Opinion. Human. Reprod. 2001, 16, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Baroni, T.; Arato, I.; Mancuso, F.; Calafiore, R.; Luca, G. On the Origin of Testicular Germ Cell Tumors: From Gonocytes to Testicular Cancer. Front. Endocrinol. 2019, 10, 343. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione Transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Board, P.G.; Menon, D. Glutathione Transferases, Regulators of Cellular Metabolism and Physiology. Biochim. Biophys. Acta—General. Subj. 2013, 1830, 3267–3288. [Google Scholar] [CrossRef]
- Mijatović, S.; Savić-Radojević, A.; Plješa-Ercegovac, M.; Simić, T.; Nicoletti, F.; Maksimović-Ivanić, D. The Double-Faced Role of Nitric Oxide and Reactive Oxygen Species in Solid Tumors. Antioxidants 2020, 9, 374. [Google Scholar] [CrossRef]
- Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Coric, V.; Radic, T.; Simic, T. Glutathione Transferase Genotypes May Serve as Determinants of Risk and Prognosis in Renal Cell Carcinoma. Biofactors 2020, 46, 229–238. [Google Scholar] [CrossRef]
- Di Pietro, G.; Magno, L.A.V.; Rios-Santos, F. Glutathione S-Transferases: An Overview in Cancer Research. Expert Opin. Drug Metab. Toxicol. 2010, 6, 153–170. [Google Scholar] [CrossRef]
- Simic, T.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Matic, M.; Mimic-Oka, J. Glutathione S-Transferases in Kidney and Urinary Bladder Tumors. Nat. Rev. Urol. 2009, 6, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Mannervik, B.; Danielson, U.H. Glutathione Transferases—Structure and Catalytic Activity. CRC Crit. Rev. Biochem. 1988, 23, 283–337. [Google Scholar] [CrossRef]
- Board, P.G.; Coggan, M.; Chelvanayagam, G.; Easteal, S.; Jermiin, L.S.; Schulte, G.K.; Danley, D.E.; Hoth, L.R.; Griffor, M.C.; Kamath, A.V.; et al. Identification, Characterization, and Crystal Structure of the Omega Class Glutathione Transferases. J. Biol. Chem. 2000, 275, 24798–24806. [Google Scholar] [CrossRef]
- Whitbread, A.K.; Masoumi, A.; Tetlow, N.; Schmuck, E.; Coggan, M.; Board, P.G. Characterization of the Omega Class of Glutathione Transferases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2005; Volume 401, pp. 78–99. ISBN 978-0-12-182806-6. [Google Scholar]
- Board, P.G.; Menon, D. Structure, Function and Disease Relevance of Omega-Class Glutathione Transferases. Arch. Toxicol. 2016, 90, 1049–1067. [Google Scholar] [CrossRef]
- Hughes, M.M.; Hooftman, A.; Angiari, S.; Tummala, P.; Zaslona, Z.; Runtsch, M.C.; McGettrick, A.F.; Sutton, C.E.; Diskin, C.; Rooke, M.; et al. Glutathione Transferase Omega-1 Regulates NLRP3 Inflammasome Activation through NEK7 Deglutathionylation. Cell Rep. 2019, 29, 151–161.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Brock, J.; Liu, D.; Board, P.G.; Oakley, A.J. Structural Insights into the Dehydroascorbate Reductase Activity of Human Omega-Class Glutathione Transferases. J. Mol. Biol. 2012, 420, 190–203. [Google Scholar] [CrossRef]
- Hamilton, L.E.; Zigo, M.; Mao, J.; Xu, W.; Sutovsky, P.; O’Flaherty, C.; Oko, R. GSTO2 Isoforms Participate in the Oxidative Regulation of the Plasmalemma in Eutherian Spermatozoa during Capacitation. Antioxidants 2019, 8, 601. [Google Scholar] [CrossRef] [PubMed]
- Radic, T.M.; Coric, V.M.; Pljesa-Ercegovac, M.S.; Basta-Jovanovic, G.M.; Radojevic-Skodric, S.M.; Dragicevic, D.P.; Matic, M.G.; Bogdanovic, L.M.; Dzamic, Z.M.; Simic, T.P.; et al. Concomitance of Polymorphisms in Glutathione Transferase Omega Genes Is Associated with Risk of Clear Cell Renal Cell Carcinoma. Tohoku J. Exp. Med. 2018, 246, 35–44. [Google Scholar] [CrossRef]
- Radic, T.; Coric, V.; Bukumiric, Z.; Pljesa-Ercegovac, M.; Djukic, T.; Avramovic, N.; Matic, M.; Mihailovic, S.; Dragicevic, D.; Dzamic, Z.; et al. GSTO1*CC Genotype (Rs4925) Predicts Shorter Survival in Clear Cell Renal Cell Carcinoma Male Patients. Cancers 2019, 11, 2038. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.I.; Savic-Radojevic, A.R.; Pekmezovic, T.D.; Matic, M.G.; Pljesa-Ercegovac, M.S.; Coric, V.M.; Radic, T.M.; Suvakov, S.R.; Krivic, B.N.; Dragicevic, D.P.; et al. Glutathione S-Transferase T1, O1 and O2 Polymorphisms Are Associated with Survival in Muscle Invasive Bladder Cancer Patients. PLoS ONE 2013, 8, e74724. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.; Simic, T.; Radic, T.; Matic, M.; Pljesa-Ercegovac, M.; Suvakov, S.; Coric, V.; Pekmezovic, T.; Novakovic, I.; Dragicevic, D.; et al. GSTO1*C/GSTO2*G Haplotype Is Associated with Risk of Transitional Cell Carcinoma of Urinary Bladder. Int. Urol. Nephrol. 2015, 47, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.; Simic, T.; Pljesa-Ercegovac, M.; Matic, M.; Suvakov, S.; Coric, V.; Dragicevic, D.; Savic-Radojevic, A. Upregulated Glutathione Transferase Omega-1 Correlates with Progression of Urinary Bladder Carcinoma. Redox Rep. 2017, 22, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.; Stevanovic, G.; Coric, V.; Bukumiric, Z.; Pljesa-Ercegovac, M.; Matic, M.; Jerotic, D.; Todorovic, N.; Asanin, M.; Ercegovac, M.; et al. GSTO1, GSTO2 and ACE2 Polymorphisms Modify Susceptibility to Developing COVID-19. J. Pers. Med. 2022, 12, 458. [Google Scholar] [CrossRef]
- Tanaka-Kagawa, T.; Jinno, H.; Hasegawa, T.; Makino, Y.; Seko, Y.; Hanioka, N.; Ando, M. Functional Characterization of Two Variant Human GSTO 1-1s (Ala140Asp and Thr217Asn). Biochem. Biophys. Res. Commun. 2003, 301, 516–520. [Google Scholar] [CrossRef]
- Whitbread, A.K.; Tetlow, N.; Eyre, H.J.; Sutherland, G.R.; Board, P.G. Characterization of the Human Omega Class Glutathione Transferase Genes and Associated Polymorphisms. Pharmacogenetics 2003, 13, 131–144. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Yeh, S.-D.; Shen, K.-H.; Shen, C.-H.; Juang, G.-D.; Hsu, L.-I.; Chiou, H.-Y.; Chen, C.-J. A Significantly Joint Effect between Arsenic and Occupational Exposures and Risk Genotypes/Diplotypes of CYP2E1, GSTO1 and GSTO2 on Risk of Urothelial Carcinoma. Toxicol. Appl. Pharmacol. 2009, 241, 111–118. [Google Scholar] [CrossRef]
- Bumbasirevic, U.; Bojanic, N.; Pljesa-Ercegovac, M.; Zivkovic, M.; Djukic, T.; Zekovic, M.; Milojevic, B.; Kajmakovic, B.; Janicic, A.; Simic, T.; et al. The Polymorphisms of Genes Encoding Catalytic Antioxidant Proteins Modulate the Susceptibility and Progression of Testicular Germ Cell Tumor. Cancers 2022, 14, 1068. [Google Scholar] [CrossRef]
- Harries, L.W.; Stubbins, M.J.; Forman, D.; Howard, G.C.; Wolf, C.R. Identification of Genetic Polymorphisms at the Glutathione S-Transferase Pi Locus and Association with Susceptibility to Bladder, Testicular and Prostate Cancer. Carcinogenesis 1997, 18, 641–644. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, J.; Kraggerud, S.M.; Brydøy, M.; Cvancarova, M.; Lothe, R.A.; Fossa, S.D. Association between Long-Term Neuro-Toxicities in Testicular Cancer Survivors and Polymorphisms in Glutathione-s-Transferase-P1 and -M1, a Retrospective Cross Sectional Study. J. Transl. Med. 2007, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Kraggerud, S.M.; Oldenburg, J.; Alnaes, G.I.; Berg, M.; Kristensen, V.N.; Fossa, S.D.; Lothe, R.A. Functional Glutathione S-Transferase Genotypes among Testicular Germ Cell Tumor Survivors: Associations with Primary and Post-Chemotherapy Tumor Histology. Pharm. Genom. 2009, 19, 751–759. [Google Scholar] [CrossRef]
- Roco, A.; Lavanderos, A.; Cayún, J.P.; Acevedo, C.; Celedón, C.; Rubilar, J.C.; Sandoval, C.; Cerpa, L.; García-Martín, E.; Agúndez, J.A.; et al. The Role of Phase I and II Genetic Polymorphisms, Smoking, Alcohol and Cancer Family History, in the Risk of Developing Testicular Cancer. Pharm. Genom. 2019, 29, 159–166. [Google Scholar] [CrossRef]
- Bumbasirevic, U.; Bojanic, N.; Simic, T.; Milojevic, B.; Zivkovic, M.; Kosanovic, T.; Kajmakovic, B.; Janicic, A.; Durutovic, O.; Radovanovic, M.; et al. Interplay between Comprehensive Inflammation Indices and Redox Biomarkers in Testicular Germ-Cell Tumors. J. Pers. Med. 2022, 12, 833. [Google Scholar] [CrossRef]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and Visualization of LD and Haplotype Maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A Web Tool for the Analysis of Association Studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef]
- Hedrick, P.W. Genetics of Populations, 4th ed.; Jones and Bartlett Publishers: Sudbury, MA, USA, 2011; ISBN 978-0-7637-5737-3. [Google Scholar]
- Armstrong, R.N. Glutathione Transferases. In Comprehensive Toxicology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 295–321. ISBN 978-0-08-046884-6. [Google Scholar]
- Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Matic, M.; Coric, V.; Djukic, T.; Radic, T.; Simic, T. Glutathione Transferases: Potential Targets to Overcome Chemoresistance in Solid Tumors. Int. J. Mol. Sci. 2018, 19, 3785. [Google Scholar] [CrossRef]
- Bartolini, D.; Galli, F. The Functional Interactome of GSTP: A Regulatory Biomolecular Network at the Interface with the Nrf2 Adaption Response to Oxidative Stress. J. Chromatogr. B 2016, 1019, 29–44. [Google Scholar] [CrossRef]
- Schmuck, E.M.; Board, P.G.; Whitbread, A.K.; Tetlow, N.; Cavanaugh, J.A.; Blackburn, A.C.; Masoumi, A. Characterization of the Monomethylarsonate Reductase and Dehydroascorbate Reductase Activities of Omega Class Glutathione Transferase Variants: Implications for Arsenic Metabolism and the Age-at-Onset of Alzheimer’s and Parkinson’s Diseases. Pharmacogenet. Genom. 2005, 15, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Zakharyan, R.A.; Sampayo-Reyes, A.; Healy, S.M.; Tsaprailis, G.; Board, P.G.; Liebler, D.C.; Aposhian, H.V. Human Monomethylarsonic Acid (MMA(V)) Reductase Is a Member of the Glutathione-S-Transferase Superfamily. Chem. Res. Toxicol. 2001, 14, 1051–1057. [Google Scholar] [CrossRef]
- Rachamalla, M.; Chinthada, J.; Kushwaha, S.; Putnala, S.K.; Sahu, C.; Jena, G.; Niyogi, S. Contemporary Comprehensive Review on Arsenic-Induced Male Reproductive Toxicity and Mechanisms of Phytonutrient Intervention. Toxics 2022, 10, 744. [Google Scholar] [CrossRef] [PubMed]
- Tabrez, S.; Priyadarshini, M.; Priyamvada, S.; Khan, M.S.; Na, A.; Zaidi, S.K. Gene-Environment Interactions in Heavy Metal and Pesticide Carcinogenesis. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 760, 1–9. [Google Scholar] [CrossRef]
- Dai, G.; Chou, N.; He, L.; Gyamfi, M.A.; Mendy, A.J.; Slitt, A.L.; Klaassen, C.D.; Wan, Y.-J.Y. Retinoid X Receptor α Regulates the Expression of Glutathione S-Transferase Genes and Modulates Acetaminophen-Glutathione Conjugation in Mouse Liver. Mol. Pharmacol. 2005, 68, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Dulhunty, A.; Gage, P.; Curtis, S.; Chelvanayagam, G.; Board, P. The Glutathione Transferase Structural Family Includes a Nuclear Chloride Channel and a Ryanodine Receptor Calcium Release Channel Modulator. J. Biol. Chem. 2001, 276, 3319–3323. [Google Scholar] [CrossRef] [PubMed]
- Laliberte, R.E.; Perregaux, D.G.; Hoth, L.R.; Rosner, P.J.; Jordan, C.K.; Peese, K.M.; Eggler, J.F.; Dombroski, M.A.; Geoghegan, K.F.; Gabel, C.A. Glutathione S-Transferase Omega 1-1 Is a Target of Cytokine Release Inhibitory Drugs and May Be Responsible for Their Effect on Interleukin-1β Posttranslational Processing. J. Biol. Chem. 2003, 278, 16567–16578. [Google Scholar] [CrossRef]
- Piaggi, S.; Raggi, C.; Corti, A.; Pitzalis, E.; Mascherpa, M.C.; Saviozzi, M.; Pompella, A.; Casini, A.F. Glutathione Transferase Omega 1-1 (GSTO1-1) Plays an Anti-Apoptotic Role in Cell Resistance to Cisplatin Toxicity. Carcinogenesis 2010, 31, 804–811. [Google Scholar] [CrossRef]
- Yan, X.; Pan, L.; Yuan, Y.; Lang, J.; Mao, N. Identification of Platinum-Resistance Associated Proteins through Proteomic Analysis of Human Ovarian Cancer Cells and Their Platinum-Resistant Sublines. J. Proteome Res. 2007, 6, 772–780. [Google Scholar] [CrossRef]
- Mukherjee, B.; Salavaggione, O.E.; Pelleymounter, L.L.; Moon, I.; Eckloff, B.W.; Schaid, D.J.; Wieben, E.D.; Weinshilboum, R.M. Glutathione S-Transferase Omega 1 and Omega 2 Pharmacogenomics. Drug Metab. Dispos. 2006, 34, 1237–1246. [Google Scholar] [CrossRef]
- Sanguansin, S.; Petmitr, S.; O-charoenrat, P.; Pongstaporn, W. Association of Glutathione S-Transferase Omega Gene Polymorphisms with Progression of Head and Neck Cancer. Mol. Biol. Rep. 2012, 39, 10915–10920. [Google Scholar] [CrossRef]
- Pongstaporn, W.; Rochanawutanon, M.; Wilailak, S.; Linasamita, V.; Weerakiat, S.; Petmitr, S. Genetic Alterations in Chromosome 10q24.3 and Glutathione S-Transferase Omega 2 Gene Polymorphism in Ovarian Cancer. J. Exp. Clin. Cancer Res. 2006, 25, 107–114. [Google Scholar] [PubMed]
- Lesseur, C.; Gilbert-Diamond, D.; Andrew, A.S.; Ekstrom, R.M.; Li, Z.; Kelsey, K.T.; Marsit, C.J.; Karagas, M.R. A Case-Control Study of Polymorphisms in Xenobiotic and Arsenic Metabolism Genes and Arsenic-Related Bladder Cancer in New Hampshire. Toxicol. Lett. 2012, 210, 100–106. [Google Scholar] [CrossRef]
- Masoudi, M.; Saadat, I.; Omidvari, S.; Saadat, M. Genetic Polymorphisms of GSTO2, GSTM1, and GSTT1 and Risk of Gastric Cancer. Mol. Biol. Rep. 2009, 36, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, M.; Saadat, I.; Omidvari, S.; Saadat, M. Association between N142D Genetic Polymorphism of GSTO2 and Susceptibility to Colorectal Cancer. Mol. Biol. Rep. 2011, 38, 4309–4313. [Google Scholar] [CrossRef]
- Xu, Y.-T.; Wang, J.; Yin, R.; Qiu, M.-T.; Xu, L.; Wang, J.; Xu, L. Genetic Polymorphisms in Glutathione S-Transferase Omega (GSTO) and Cancer Risk: A Meta-Analysis of 20 Studies. Sci. Rep. 2014, 4, 6578. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Allamaneni, S.S.R. The Effect of Sperm DNA Damage on Assisted Reproduction Outcomes. A Review. Minerva Ginecol. 2004, 56, 235–245. [Google Scholar]
- Agarwal, A.; Said, T.M. Oxidative Stress, DNA Damage and Apoptosis in Male Infertility: A Clinical Approach. BJU Int. 2005, 95, 503–507. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative Stress and Male Infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Mustacich, D.; Powis, G. Thioredoxin Reductase. Biochem. J. 2000, 346 Pt 1, 1–8. [Google Scholar] [CrossRef]
- Board, P.G. The Omega-Class Glutathione Transferases: Structure, Function, and Genetics. Drug Metab. Rev. 2011, 43, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, S.; Monaci, P.M.; Polimanti, R.; Manfellotto, D.; Fuciarelli, M. GSTO2*N142D Gene Polymorphism Associated with Hypothyroidism in Italian Patients. Mol. Biol. Rep. 2013, 40, 1967–1971. [Google Scholar] [CrossRef] [PubMed]


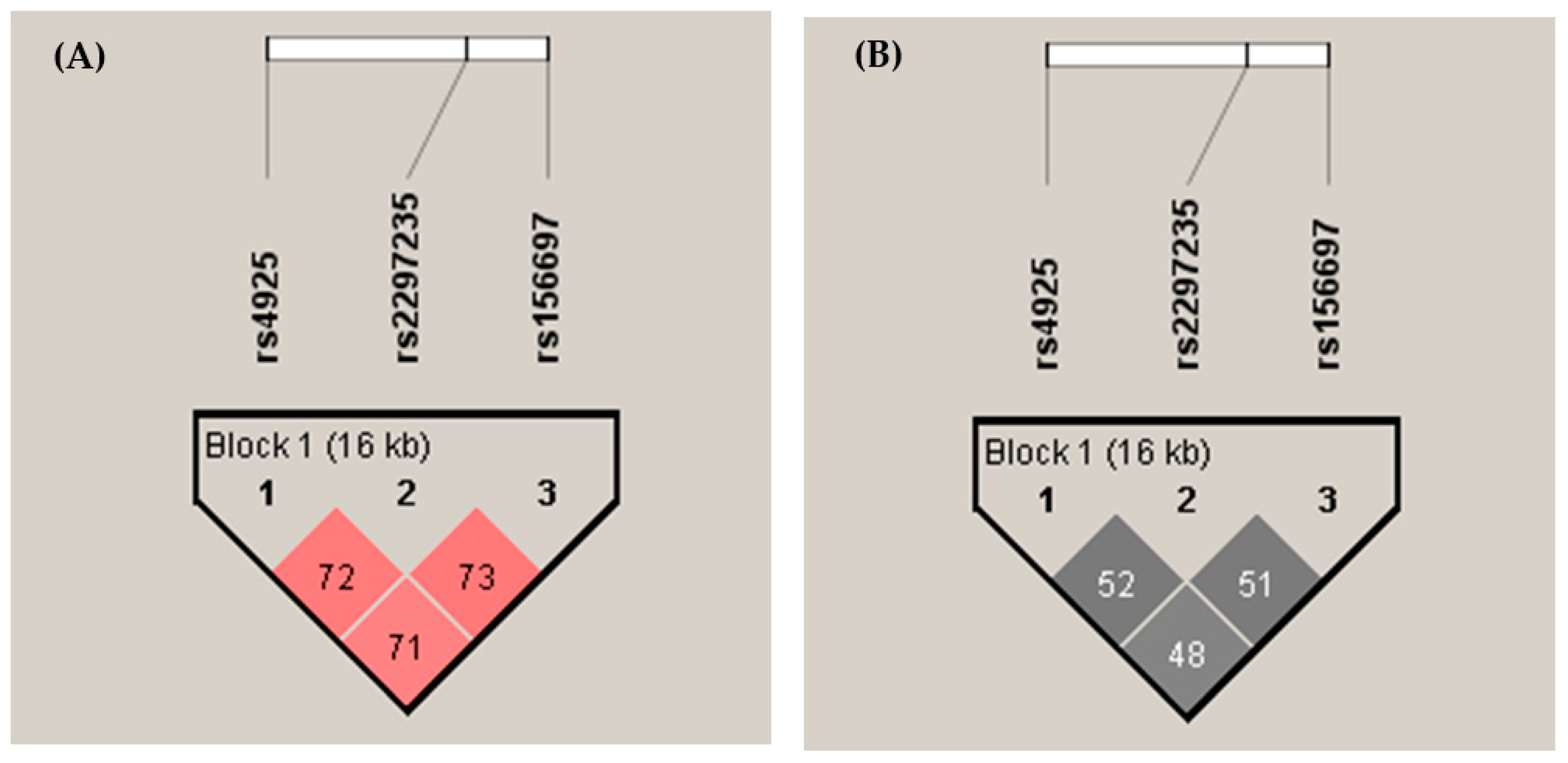{kind=link}
| Parameters | Testicular GCT Patients |
|---|---|
| Age (years) | 33.5 ± 8.7 1 33.5 (19–54) 2 |
| Risk factors for testicular GCT development, n (%) | |
| Cryptorchismus | 8 (9) |
| Infertility | 4 (5) |
| Testicular atrophy | 7 (8) |
| Testicular dysgenesis syndrome | 1 (1) |
| Positive family history | 4 (5) |
| Tumor type | |
| Seminoma | 52 (59) |
| Non-seminoma | 36 (41) |
| Clinical stage | |
| I | 61 (69) |
| II | 18 (21) |
| III | 9 (10) |
| Pathological stage | |
| pT1 | 40 (47) |
| pT2 | 39 (46) |
| pT3 | 5 (6) |
| pT4 | 1 (1) |
| Genotype | Patients n (%) | Controls n (%) | Crude OR1 1 (95% CI) | p | OR2 2 (95% CI) | p |
|---|---|---|---|---|---|---|
| GSTO1rs4925 3 | ||||||
| *A/A | 6 (7) | 13 (14) | 1.00 | - | 1.00 (reference group) | - |
| *C/A*C/C | 81 (93) | 82 (86) | 2.14 (0.77–5.90) | 0.142 | 3.20 (1.1–9.35) | 0.033 |
| GSTO2rs156697 4 | ||||||
| *A/A | 33 (39) | 50 (52) | 1.00 | - | 1.00 (reference group) | - |
| *A/G*G/G | 51 (61) | 46 (48) | 1.68 (0.93–3.04) | 0.087 | 1.34 (0.59–3.27) | 0.455 |
| GSTO2rs2297235 5 | ||||||
| *A/A | 31 (35) | 49 (51) | 1.00 | - | 1.00 (reference group) | - |
| *A/G*G/G | 57 (65) | 47 (49) | 1.92 (1.06–3.47) | 0.031 | 1.80 (0.75–4.27) | 0.186 |
| GSTO2rs156697/ GSTO2rs2297235 | Patients n (%) | Controls n (%) | Crude OR1 1 (95% CI) | p | OR2 2 (95% CI) | p |
|---|---|---|---|---|---|---|
| *A/A + *A/A | 29 (34) | 40 (42) | 1.00 | - | 1.00 | |
| *A/A + *A/G*G/G | 4 (5) | 10 (10) | 0.55 (0.16–1.93) | 0.354 | 0.59 (0.17–2.08) | 0.413 |
| *A/G*G/G + *A/A | 2 (3) | 9 (9) | 0.37 (0.06–1.52) | 0.149 | 0.31 (0.06–1.52) | 0.149 |
| *A/G*G/G + *A/G*G/G | 49 (58) | 37 (39) | 1.83 (0.96–3.47) | 0.065 | 2.49 (1.25–4.96) | 0.010 |
| Haplotype | GSTO1 rs4925 | GSTO2 rs2297235 | GSTO2 rs156697 | Frequency | Crude OR (95% CI) | p |
|---|---|---|---|---|---|---|
| H1 | *C | *A | *A | 0.563 | 1.00 | - |
| H2 | *A | *G | *G | 0.245 | 1.30 (0.79–2.14) | 0.312 |
| H3 | *A | *A | *A | 0.040 | 0.54 (0.15–1.94) | 0.355 |
| H4 | *C | *G | *A | 0.034 | 0.73 (0.22–2.42) | 0.619 |
| H5 | *C | *A | *G | 0.037 | 1.75 (0.52–5.87) | 0.374 |
| H6 | *A | *A | *G | 0.022 | 0.27 (0.04–1.67) | 0.166 |
| H7 | *C | *G | *G | 0.027 | 3.37 (0.60–18.78) | 0.174 |
| H8 | *A | *G | *A | 0.032 | 0.41 (0.07–2.31) | 0.315 |
| GSTO1rs4925/GSTO2rs156697/GSTO2rs2297235 | Patients n (%) | Controls n (%) | Crude OR1 1 (95% CI) | p |
|---|---|---|---|---|
| 1 | 29 (34) | 41 (43) | 1.00 (reference group) | |
| 2 | 12 (14) | 30 (32) | 0.57 (0.24–1.28) | 0.174 |
| 3 | 43 (51) | 24 (25) | 2.53 (1.27–5.04) | 0.008 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrovic, M.; Simic, T.; Djukic, T.; Radic, T.; Savic-Radojevic, A.; Zekovic, M.; Durutovic, O.; Janicic, A.; Milojevic, B.; Kajmakovic, B.; et al. The Polymorphisms in GSTO Genes (GSTO1 rs4925, GSTO2 rs156697, and GSTO2 rs2297235) Affect the Risk for Testicular Germ Cell Tumor Development: A Pilot Study. Life 2023, 13, 1269. https://doi.org/10.3390/life13061269
Petrovic M, Simic T, Djukic T, Radic T, Savic-Radojevic A, Zekovic M, Durutovic O, Janicic A, Milojevic B, Kajmakovic B, et al. The Polymorphisms in GSTO Genes (GSTO1 rs4925, GSTO2 rs156697, and GSTO2 rs2297235) Affect the Risk for Testicular Germ Cell Tumor Development: A Pilot Study. Life. 2023; 13(6):1269. https://doi.org/10.3390/life13061269
Chicago/Turabian StylePetrovic, Milos, Tatjana Simic, Tatjana Djukic, Tanja Radic, Ana Savic-Radojevic, Milica Zekovic, Otas Durutovic, Aleksandar Janicic, Bogomir Milojevic, Boris Kajmakovic, and et al. 2023. "The Polymorphisms in GSTO Genes (GSTO1 rs4925, GSTO2 rs156697, and GSTO2 rs2297235) Affect the Risk for Testicular Germ Cell Tumor Development: A Pilot Study" Life 13, no. 6: 1269. https://doi.org/10.3390/life13061269
APA StylePetrovic, M., Simic, T., Djukic, T., Radic, T., Savic-Radojevic, A., Zekovic, M., Durutovic, O., Janicic, A., Milojevic, B., Kajmakovic, B., Zivkovic, M., Bojanic, N., Bumbasirevic, U., & Coric, V. (2023). The Polymorphisms in GSTO Genes (GSTO1 rs4925, GSTO2 rs156697, and GSTO2 rs2297235) Affect the Risk for Testicular Germ Cell Tumor Development: A Pilot Study. Life, 13(6), 1269. https://doi.org/10.3390/life13061269