
Effects of Anticoccidial Vaccination and Taraxacum officinale Extract on the Growth Performance, Biochemical Parameters, Immunity, and Intestinal Morphology of Eimeria-Challenged Chickens

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Birds, and Diets
2.2. Experimental Factors
2.3. Sample Collection and Analytical Procedure
2.4. Statistical Analysis
3. Results
3.1. Growth Performance
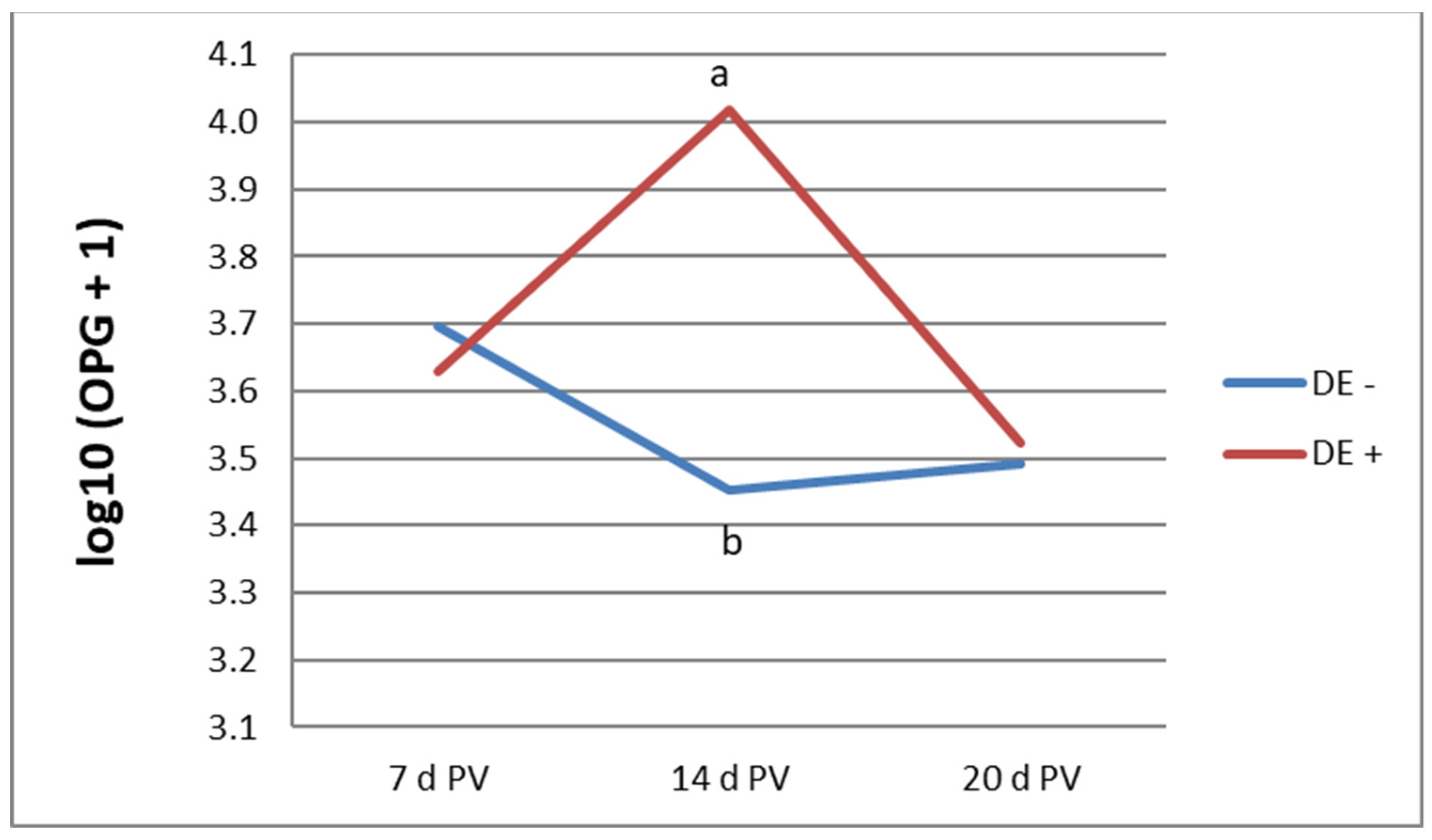
3.2. Oocyst Shedding and Lesion Score
3.3. Intestinal Morphology and Epithelial Barrier Characteristics
3.4. Immunoglobulin, Cytokines, and Acute Phase Protein Levels
3.5. Biochemical Parameters
3.6. Volatile Fatty Acids in Cecal Digesta
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohsin, M.; Li, Y.; Zhang, X.; Wang, Y.; Huang, Z.; Yin, G.; Zhang, Z. Development of CRISPR-CAS9 Based RNA Drugs against Eimeria tenella Infection. Genomics 2021, 113, 4126–4135. [Google Scholar] [CrossRef]
- Mohsin, M.; Li, L.; Huang, X.; Aleem, M.T.; Habib, Y.J.; Ismael, A.; Afzal, M.Z.; Abbas, R.Z.; Abbas, A.; Yin, G. Immunogenicity and Protective Efficacy of Probiotics with EtIMP1C against Eimeria tenella Challenge. Pak. Vet. J. 2021, 41, 274–278. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-Calculating the Cost of Coccidiosis in Chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D.; Cherry, T.E.; Danforth, H.D.; Richards, G.; Shirley, M.W.; Williams, R.B. Sustainable Coccidiosis Control in Poultry Production: The Role of Live Vaccines. Int. J. Parasitol. 2002, 32, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B. Intercurrent Coccidiosis and Necrotic Enteritis of Chickens: Rational, Integrated Disease Management by Maintenance of Gut Integrity. Avian Pathol. 2005, 34, 159–180. [Google Scholar] [CrossRef]
- Soutter, F.; Werling, D.; Tomley, F.M.; Blake, D.P. Poultry Coccidiosis: Design and Interpretation of Vaccine Studies. Front. Vet. Sci. 2020, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Yan, Y.; Jian, F.; Ning, C. Coccidia-Microbiota Interactions and Their Effects on the Host. Front. Cell. Infect. Microbiol. 2021, 11, 751481. [Google Scholar] [CrossRef]
- Lin, X. Evaluation of Immunogenicity and Protective Efficacy of Eimeria Maxima Immune Mapped Protein 1 with EDA Adjuvant in Chicken. Pak. Vet. J. 2020, 40, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.P.; Tomley, F.M. Securing Poultry Production from the Ever-Present Eimeria Challenge. Trends Parasitol. 2014, 30, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Peek, H.W.; Landman, W.J.M. Coccidiosis in Poultry: Anticoccidial Products, Vaccines and Other Prevention Strategies. Vet. Q. 2011, 31, 143–161. [Google Scholar] [CrossRef]
- Orso, C.; Stefanello, T.B.; Franceschi, C.H.; Mann, M.B.; Varela, A.P.M.; Castro, I.M.S.; Frazzon, J.; Frazzon, A.P.G.; Andretta, I.; Ribeiro, A.M.L. Changes in the Ceca Microbiota of Broilers Vaccinated for Coccidiosis or Supplemented with Salinomycin. Poult. Sci. 2021, 100, 100969. [Google Scholar] [CrossRef] [PubMed]
- Lehman, R.; Moran, E.T.; Hess, J.B. Response of Coccidiostat- versus Vaccination-Protected Broilers to Gelatin Inclusion in High and Low Crude Protein Diets. Poult. Sci. 2009, 88, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Broussard, C.; Fitz-Coy, S.; Burke, P.; Eckert, N.H.; Stevens, S.M.; Anderson, P.N.; Anderson, S.M.; Caldwell, D.J. Evaluation of Live Oocyst Vaccination or Salinomycin for Control of Field-Strain Eimeria Challenge in Broilers on Two Different Feeding Programs. J. Appl. Poult. Res. 2009, 18, 458–464. [Google Scholar] [CrossRef]
- Allen, P.C.; Fetterer, R.H. Recent Advances in Biology and Immunobiology of Eimeria Species and in Diagnosis and Control of Infection with These Coccidian Parasites of Poultry. Clin. Microbiol. Rev. 2002, 15, 58–65. [Google Scholar] [CrossRef]
- Lee, Y.; Lu, M.; Lillehoj, H.S. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines 2022, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Arczewska-Włosek, A.; Świątkiewicz, S. Nutrition as a Modulatory Factor of the Efficacy of Live Anticoccidial Vaccines in Broiler Chickens. Worlds Poult. Sci. J. 2014, 70, 81–92. [Google Scholar] [CrossRef]
- de Andrade, R.M.; Pagnussatt, H.; Talian, L.E.; Santo, A.D.; Ribeiro, A.B.; Leite, F.; Mis, G.; Hoinoski, G.; Aniecevski, E.; Fabiani, L.M.; et al. Interaction between Live Vaccines for Coccidiosis and Phytogenic Compounds in the Diet of Broilers. Parasitol. Int. 2022, 89, 102584. [Google Scholar] [CrossRef]
- Saeed, Z.; Alkheraije, K.A. Botanicals: A Promising Approach for Controlling Cecal Coccidiosis in Poultry. Front. Vet. Sci. 2023, 10, 1157633. [Google Scholar] [CrossRef]
- Park, I.; Nam, H.; Wickramasuriya, S.S.; Lee, Y.; Wall, E.H.; Ravichandran, S.; Lillehoj, H.S. Host-Mediated Beneficial Effects of Phytochemicals for Prevention of Avian Coccidiosis. Front. Immunol. 2023, 14, 1145367. [Google Scholar] [CrossRef]
- Yang, C.; Das, Q.; Rehman, M.A.; Yin, X.; Shay, J.; Gauthier, M.; Lau, C.H.-F.; Ross, K.; Diarra, M.S. Microbiome of Ceca from Broiler Chicken Vaccinated or Not against Coccidiosis and Fed Berry Pomaces. Microorganisms 2023, 11, 1184. [Google Scholar] [CrossRef]
- Qureshi, S.; Adil, S.; El-Hack, M.E.A.; Alagawany, M.; Farag, M.R. Beneficial Uses of Dandelion Herb (Taraxacum officinale) in Poultry Nutrition. Worlds Poult. Sci. J. 2017, 73, 591–602. [Google Scholar] [CrossRef]
- Grauso, L.; Emrick, S.; de Falco, B.; Lanzotti, V.; Bonanomi, G. Common Dandelion: A Review of Its Botanical, Phytochemical and Pharmacological Profiles. Phytochem. Rev. 2019, 18, 1115–1132. [Google Scholar] [CrossRef]
- Jędrejek, D.; Kontek, B.; Lis, B.; Stochmal, A.; Olas, B. Evaluation of Antioxidant Activity of Phenolic Fractions from the Leaves and Petals of Dandelion in Human Plasma Treated with H2O2 and H2O2/Fe. Chem. Biol. Interact. 2017, 262, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Noor, A.S.; Kadhim, A.H.; Ali, M.A. The Effect of Feeding Different Levels of Dandelion Leaf Powder (Taraxacum officinale) in the Diet on the Productive and Physiological Performance of Broiler Chickens, Strain Ross-308. IOP Conf. Ser. Earth Environ. Sci. 2021, 722, 012002. [Google Scholar] [CrossRef]
- Mao, J.; Wang, Y.; Wang, W.; Duan, T.; Yin, N.; Guo, T.; Guo, H.; Liu, N.; An, X.; Qi, J. Effects of Taraxacum mongolicum Hand.-Mazz. (Dandelion) on Growth Performance, Expression of Genes Coding for Tight Junction Protein and Mucin, Microbiota Composition and Short Chain Fatty Acids in Ileum of Broiler Chickens. BMC Vet. Res. 2022, 18, 180. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.; Banday, M.T.; Adil, S.; Shakeel, I.; Munshi, Z.H. Effect of Dandelion Leaves and Fenugreek Seeds with or without Enzyme Addition on Performance and Blood Biochemistry of Broiler Chicken, and Evaluation of Their in Vitro Antibacterial Activity. Indian J. Anim. Sci. 2015, 85, 1248–1254. [Google Scholar] [CrossRef]
- Smulikowska, S.; Rutkowski, A. Recommended Allowances and Nutritive Value of Feedstuffs. Poultry Feeding Standards, 4th ed.; The Kielanowski Institute of Animal Physiology and Nutrition, PAS, Polish Branch of WPSA: Jabłonna, Poland, 2005; ISBN 83-917097-7-9. (In Polish) [Google Scholar]
- FAO. Epidemiology, Diagnosis and Control of Poultry Parasites; Manuel FAO de Santé Animale; FAO: Rome, Italy, 1998; ISBN 92-5-104215-2. [Google Scholar]
- Johnson, J.; Reid, W.M. Anticoccidial Drugs: Lesion Scoring Techniques in Battery and Floor-Pen Experiments with Chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.; Świątkiewicz, S.; Arczewka-Włosek, A.; Muszyński, S.; Szymańczyk, S.; Arciszewski, M.B.; Siembida, A.Z.; Kras, K.; Piedra, J.L.V.; Schwarz, T.; et al. Modern Hybrid Rye, as an Alternative Energy Source for Broiler Chickens, Improves the Absorption Surface of the Small Intestine Depending on the Intestinal Part and Xylanase Supplementation. Animals 2021, 11, 1349. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Rudyk, H.; Dobrowolski, P.; Arciszewski, M.B.; Donaldson, J.; Kras, K.; Abramowicz, B.; Kuc, D.; Muszyński, S. Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins. Animals 2023, 13, 1538. [Google Scholar] [CrossRef] [PubMed]
- Kisielinski, K.; Willis, S.; Prescher, A.; Klosterhalfen, B.; Schumpelick, V. A Simple New Method to Calculate Small Intestine Absorptive Surface in the Rat. Clin. Exp. Med. 2002, 2, 131–135. [Google Scholar] [CrossRef]
- Muszyński, S.; Hułas-Stasiak, M.; Dobrowolski, P.; Arciszewski, M.B.; Hiżewska, L.; Donaldson, J.; Mozel, S.; Rycerz, K.; Kapica, M.; Puzio, I.; et al. Maternal Acrylamide Exposure Changes Intestinal Epithelium, Immunolocalization of Leptin and Ghrelin and Their Receptors, and Gut Barrier in Weaned Offspring. Sci. Rep. 2023, 13, 10286. [Google Scholar] [CrossRef] [PubMed]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2006; ISBN 978-0-935584-77-6. [Google Scholar]
- Sharman, P.A.; Smith, N.C.; Wallach, M.G.; Katrib, M. Chasing the Golden Egg: Vaccination against Poultry Coccidiosis. Parasite Immunol. 2010, 32, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peebles, E.D.; Kiess, A.S.; Wamsley, K.G.S.; Zhai, W. Effects of Coccidial Vaccination and Dietary Antimicrobial Alternatives on the Growth Performance, Internal Organ Development, and Intestinal Morphology of Eimeria-Challenged Male Broilers. Poult. Sci. 2019, 98, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Lee, K.W.; Bravo, D.; Lillehoj, E.P. Effects of Dietary Supplementation with Phytonutrients on Vaccine-Stimulated Immunity against Infection with Eimeria tenella. Vet. Parasitol. 2011, 181, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Kennes, Y.M.; Lepp, D.; Yin, X.; Wang, Q.; Yu, H.; Yang, C.; Gong, J.; Diarra, M.S. Effects of Encapsulated Cinnamaldehyde and Citral on the Performance and Cecal Microbiota of Broilers Vaccinated or Not Vaccinated against Coccidiosis. Poult. Sci. 2020, 99, 936–948. [Google Scholar] [CrossRef]
- Das, Q.; Shay, J.; Gauthier, M.; Yin, X.; Hasted, T.-L.; Ross, K.; Julien, C.; Yacini, H.; Kennes, Y.M.; Warriner, K.; et al. Effects of Vaccination Against Coccidiosis on Gut Microbiota and Immunity in Broiler Fed Bacitracin and Berry Pomace. Front. Immunol. 2021, 12, 621803. [Google Scholar] [CrossRef]
- Crouch, C.F.; Andrews, S.J.; Ward, R.G.; Francis, M.J. Protective Efficacy of a Live Attenuated Anticoccidial Vaccine Administered to 1-Day-Old Chickens. Avian Pathol. 2003, 32, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Livingston, M.L.; Nogal, B.; Hoang, V.; Wang, Y.-T.; Crespo, R.; Livingston, K.A. Effect of Coccidial Challenge and Vaccination on the Performance, Veterinary Postmortem Scores, and Blood Biochemistry of Broiler Chickens. Poult. Sci. 2020, 99, 3831–3840. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.F.; Allee, G.L.; Knight, C.D.; Dibner, J.J. Impact of Glutamine and Oasis Hatchling Supplement on Growth Performance, Small Intestinal Morphology, and Immune Response of Broilers Vaccinated and Challenged with Eimeria Maxima. Poult. Sci. 2005, 84, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum—A Review on Its Phytochemical and Pharmacological Profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Choct, M. Managing Gut Health through Nutrition. Br. Poult. Sci. 2009, 50, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.; Banday, M.T.; Shakeel, I.; Adil, S.; Mir, M.S.; Beigh, Y.A.; Amin, U. Histomorphological Studies of Broiler Chicken Fed Diets Supplemented with Either Raw or Enzyme Treated Dandelion Leaves and Fenugreek Seeds. Vet. World 2016, 9, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Jaramillo, F.X.; Kim, D.-H.; Lee, S.H.; Kwon, S.-K.; Jha, R.; Lee, K.-W. Role of Oregano and Citrus Species-Based Essential Oil Preparation for the Control of Coccidiosis in Broiler Chickens. J. Anim. Sci. Biotechnol. 2021, 12, 47. [Google Scholar] [CrossRef] [PubMed]
- Bedford, A.; Gong, J. Implications of Butyrate and Its Derivatives for Gut Health and Animal Production. Anim. Nutr. 2018, 4, 151–159. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of Cytokine Modulation of Epithelial Tight Junction Barrier. Front. Biosci. J. Virtual Libr. 2009, 14, 2765–2778. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric Pathogens and Their Toxin-Induced Disruption of the Intestinal Barrier through Alteration of Tight Junctions in Chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef]
- De Meyer, F.; Eeckhaut, V.; Ducatelle, R.; Dhaenens, M.; Daled, S.; Dedeurwaerder, A.; De Gussem, M.; Haesebrouck, F.; Deforce, D.; Van Immerseel, F. Host Intestinal Biomarker Identification in a Gut Leakage Model in Broilers. Vet. Res. 2019, 50, 46. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Min, W.; Dalloul, R.A. Recent Progress on the Cytokine Regulation of Intestinal Immune Responses to Eimeria1. Poult. Sci. 2004, 83, 611–623. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Yadav, S.; Castro, F.L.d.S.; Tompkins, Y.H.; Fuller, A.L.; Kim, W.K. Graded Eimeria Challenge Linearly Regulated Growth Performance, Dynamic Change of Gastrointestinal Permeability, Apparent Ileal Digestibility, Intestinal Morphology, and Tight Junctions of Broiler Chickens. Poult. Sci. 2020, 99, 4203–4216. [Google Scholar] [CrossRef]
- Castro, F.L.S.; Teng, P.-Y.; Yadav, S.; Gould, R.L.; Craig, S.; Pazdro, R.; Kim, W.K. The Effects of L-Arginine Supplementation on Growth Performance and Intestinal Health of Broiler Chickens Challenged with Eimeria spp. Poult. Sci. 2020, 99, 5844–5857. [Google Scholar] [CrossRef]
- Alagbe, E.O.; Schulze, H.; Adeola, O. Growth Performance, Nutrient Digestibility, Intestinal Morphology, Cecal Mucosal Cytokines and Serum Antioxidant Responses of Broiler Chickens to Dietary Enzymatically Treated Yeast and Coccidia Challenge. J. Anim. Sci. Biotechnol. 2023, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Zhang, Z.; Yin, G. Effect of Probiotics on the Performance and Intestinal Health of Broiler Chickens Infected with Eimeria tenella. Vaccines 2022, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.D.; Augustine, P.C. Effects of Coccidiosis on the Electrophoretic Pattern of Serum Proteins in Chickens. J. Parasitol. 1982, 68, 107–111. [Google Scholar] [CrossRef] [PubMed]
- El-Shall, N.A.; Abd El-Hack, M.E.; Albaqami, N.M.; Khafaga, A.F.; Taha, A.E.; Swelum, A.A.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; AbuQamar, S.F.; et al. Phytochemical Control of Poultry Coccidiosis: A Review. Poult. Sci. 2022, 101, 101542. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Chen, S.; Jia, R.; Huang, Y.; Zou, Y.; Li, L.; Zhao, X.; Yin, Z. Anticoccidial Effect of Herbal Powder “Shi Ying Zi” in Chickens Infected with Eimeria tenella. Animals 2020, 10, 1484. [Google Scholar] [CrossRef]
- Felici, M.; Tugnoli, B.; Ghiselli, F.; Baldo, D.; Ratti, C.; Piva, A.; Grilli, E. Investigating the Effects of Essential Oils and Pure Botanical Compounds against Eimeria tenella in Vitro. Poult. Sci. 2023, 102, 102898. [Google Scholar] [CrossRef]
- Remmal, A.; Achahbar, S.; Bouddine, L.; Chami, N.; Chami, F. In Vitro Destruction of Eimeria Oocysts by Essential Oils. Vet. Parasitol. 2011, 182, 121–126. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient [g/kg] | Starter, 1–10 d of Age | Grower 1, 11–19 d of Age | Grower 2, 20–35 d of Age | Finisher, 36–42 d of Age |
|---|---|---|---|---|
| Wheat | 100 | 100 | 100 | 100 |
| Maize | 384 | 404.7 | 413.9 | 436.9 |
| Rye | 100 | 100 | 100 | 100 |
| Soybean meal | 330 | 300 | 280 | 250 |
| Fish meal, 65% CP | 20 | 20 | 20 | 20 |
| Rapeseed oil | 30 | 41 | 52 | 60 |
| Limestone | 12 | 12 | 12 | 12 |
| Sodium chloride | 3 | 3 | 3 | 3 |
| Monocalcium phosphate | 13 | 12 | 11 | 11 |
| DL-Methionine | 2.5 | 2.3 | 2.4 | 2 |
| L-Lysine hydrochloride | 1 | 1.5 | 1.7 | 1.6 |
| L-Treonine | 0.5 | 0.5 | 1 | 0.5 |
| Vitamin–mineral premix * | 4 | 3 | 3 | 3 |
| Analyzed chemical composition (g/kg): | ||||
| Dry matter | 903 | 899 | 899 | 905 |
| Crude protein | 221 | 215 | 198 | 187 |
| Crude fat | 51.1 | 57.8 | 72.3 | 78.6 |
| Crude ash | 55.3 | 55.2 | 51.1 | 48.2 |
| Crude fiber | 29.6 | 28.3 | 2.22 | 2.18 |
| Calcium | 9.73 | 9.70 | 9.50 | 7.97 |
| Phosphorus | 6.83 | 6.71 | 6.41 | 5.98 |
| Aspartic acid | 20.93 | 19.98 | 19.94 | 17.5 |
| Threonine | 8.05 | 7.7 | 7.91 | 6.84 |
| Serine | 10.13 | 9.66 | 10.07 | 8.64 |
| Glutamic acid | 38.28 | 35.93 | 35.86 | 31.57 |
| Proline | 12.16 | 11.7 | 11.16 | 10.38 |
| Glycine | 9.23 | 9.2 | 9 | 8.12 |
| Alanine | 9.68 | 9.65 | 10.03 | 8.8 |
| Valine | 9.92 | 9.47 | 9.38 | 8.23 |
| Isoleucine | 8.66 | 8.23 | 8.18 | 7.2 |
| Leucine | 16.19 | 15.82 | 16.47 | 14.02 |
| Tyrosine | 7.27 | 7.25 | 7.54 | 6.11 |
| Phenylalanine | 11.42 | 10.73 | 10.59 | 9.32 |
| Histidine | 5.25 | 4.78 | 4.9 | 4.36 |
| Lysine | 13.24 | 12.06 | 11.99 | 10.53 |
| Arginine | 17.7 | 16.02 | 15.12 | 12.28 |
| Cysteine | 3.23 | 2.92 | 3.03 | 2.65 |
| Methionine | 4.97 | 4.87 | 5.04 | 4.06 |
| Tryptophan | 3.08 | 2.68 | 2.9 | 2.29 |
| 1–10 d (Starter) | 11–20 d (Grower 1) | 21–35 d (Grower 2) | 36–42 (Finisher) | 1–42 d | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Factors | BWG | FCR | FI | BWG | FCR | FI | BWG | FCR | FI | BWG | FCR | FI | BWG | FCR | FI | |
| ACV | DE | [g] | [g/g] | [g] | [g] | [g/g] | [g] | [g] | [g/g] | [g] | [g] | [g/g] | [g] | [g] | [g/g] | [g] |
| − | 305 | 1.28 | 389 | 706 | 1.45 b | 1024 b | 1464 b | 1.73 | 2531 | 623 | 2.28 | 1409 b | 3096 b | 1.74 | 5379 b | |
| + | 307 | 1.26 | 386 | 694 | 1.56 a | 1080 a | 1532 a | 1.72 | 2635 | 660 | 2.32 | 1519 a | 3193 a | 1.76 | 5626 a | |
| − | 313 a | 1.27 | 395 a | 692 | 1.51 | 1045 | 1490 | 1.73 | 2580 | 612 b | 2.37 | 1446 | 3107 | 1.76 | 5468 | |
| + | 299 b | 1.27 | 380 b | 707 | 1.50 | 1059 | 1506 | 1.72 | 2586 | 670 a | 2.22 | 1483 | 3182 | 1.74 | 5537 | |
| − | − | 311 | 1.28 | 399 | 692 | 1.48 | 1025 | 1445 | 1.76 | 2539 | 602 | 2.31 | 1379 | 3051 | 1.75 | 5348 |
| + | 298 | 1.27 | 379 | 719 | 1.42 | 1023 | 1482 | 1.70 | 2524 | 643 | 2.25 | 1439 | 3142 | 1.72 | 5411 | |
| + | − | 314 | 1.25 | 391 | 693 | 1.54 | 1065 | 1535 | 1.71 | 2621 | 622 | 2.44 | 1512 | 3163 | 1.77 | 5589 |
| + | 300 | 1.27 | 380 | 696 | 1.58 | 1096 | 1529 | 1.73 | 2649 | 697 | 2.19 | 1526 | 3222 | 1.76 | 5664 | |
| SEM | 2.07 | 0.029 | 2.63 | 4.08 | 0.019 | 11.5 | 15.4 | 0.019 | 35.8 | 14.6 | 0.040 | 20.2 | 23.9 | 0.013 | 53.5 | |
| p-value | ||||||||||||||||
| Effects | ACV | 0.536 | 0.164 | 0.405 | 0.120 | 0.003 | 0.012 | 0.027 | 0.821 | 0.178 | 0.178 | 0.608 | 0.004 | 0.036 | 0.362 | 0.022 |
| DE | 0.000 | 0.668 | 0.001 | 0.051 | 0.770 | 0.484 | 0.582 | 0.717 | 0.928 | 0.041 | 0.057 | 0.274 | 0.092 | 0.452 | 0.209 | |
| Interaction | ACV × DE | 0.912 | 0.213 | 0.267 | 0.112 | 0.129 | 0.408 | 0.456 | 0.327 | 0.774 | 0.520 | 0.215 | 0.496 | 0.705 | 0.671 | 0.888 |
| Factors | OPG | OPG | OPG | OPG | LS | LS | |
|---|---|---|---|---|---|---|---|
| ACV | DE | 5 d PI | 6 d PI | 7 d PI | 15 d PI | Duodenum | Jejunum/Ileum |
| − | 4.41 a | 3.74 a | 3.52 a | 2.77 | 0.76 a | 0.79 | |
| + | 2.62 b | 3.21 b | 3.17 b | 2.54 | 0.14 b | 0.41 | |
| − | 3.62 | 3.57 | 3.49 a | 3.06 a | 0.38 | 0.74 | |
| + | 3.41 | 3.39 | 3.20 b | 2.25 b | 0.53 | 0.46 | |
| − | − | 4.29 a | 3.80 | 3.66 | 3.01 | 0.70 | 0.93 |
| + | 4.53 a | 3.68 | 3.39 | 2.54 | 0.83 | 0.65 | |
| + | − | 2.95 b | 3.34 | 3.32 | 3.11 | 0.05 | 0.55 |
| + | 2.29 c | 3.09 | 3.01 | 1.96 | 0.23 | 0.27 | |
| SEM | 0.222 | 0.077 | 0.066 | 0.161 | 0.111 | 0.101 | |
| p-value | |||||||
| Effects | ACV | 0.000 | 0.000 | 0.001 | 0.392 | 0.003 | 0.059 |
| DE | 0.144 | 0.081 | 0.003 | 0.008 | 0.404 | 0.155 | |
| Interaction | ACV × DE | 0.004 | 0.504 | 0.865 | 0.226 | 0.873 | 0.982 |
| Factors | Longitudinal m. Lamina Thickness [μm] | Circular m. Lamina Thickness [μm] | Submucosa Thickness [μm] | Mucosa Thickness [μm] | Villus Height [μm] | Villus Thickness [μm] | Crypt Depth [μm] | Crypt Width [μm] | Villus Height/Crypt Depth Ratio | Mucosal Surface Absorptive Area [μm2] | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ACV | DE | ||||||||||
| − | 61.1 | 183 | 35.6 | 1365 | 1189 | 128 | 112 | 39.4 | 12.0 | 22.5 | |
| + | 64.1 | 165 | 25.7 | 1441 | 1268 | 136 | 106 | 41.4 | 12.3 | 22.5 | |
| − | 68.1 | 178 | 27.6 | 1453 | 1263 | 138 | 119 | 42.9 | 10.7 | 21.9 | |
| + | 57.0 | 169 | 33.7 | 1353 | 1193 | 127 | 99 | 37.9 | 13.6 | 23.2 | |
| − | − | 64.3 | 185 | 28.0 | 1447 | 1241 | 129 | 127 | 42.4 | 9.8 | 22.3 |
| + | 57.9 | 181 | 43.2 | 1283 | 1137 | 128 | 97 | 36.4 | 14.2 | 22.7 | |
| + | − | 71.9 | 172 | 27.2 | 1460 | 1286 | 148 | 111 | 43.5 | 11.5 | 21.4 |
| + | 56.2 | 157 | 24.3 | 1423 | 1250 | 125 | 100 | 39.4 | 13.0 | 23.7 | |
| SEM | 2.95 | 10.4 | 4.94 | 34.9 | 31.8 | 4.02 | 5.24 | 1.40 | 1.07 | 0.623 | |
| p-value | |||||||||||
| Effects | ACV | 0.604 | 0.423 | 0.343 | 0.273 | 0.234 | 0.303 | 0.549 | 0.455 | 0.914 | 0.981 |
| DE | 0.067 | 0.661 | 0.554 | 0.157 | 0.285 | 0.140 | 0.056 | 0.084 | 0.199 | 0.332 | |
| Interaction | ACV × DE | 0.416 | 0.794 | 0.383 | 0.360 | 0.597 | 0.159 | 0.361 | 0.721 | 0.506 | 0.465 |
| Factors | Longitudinal m. Lamina Thickness [μm] | Circular m. Lamina Thickness [μm] | Submucosa Thickness [μm] | Mucosa Thickness [μm] | Villus Height [μm] | Villus Thickness [μm] | Crypt Depth [μm] | Crypt Width [μm] | Villus Height/Crypt Depth Ratio | Mucosal Surface Absorptive Area [μm2] | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ACV | DE | ||||||||||
| − | 79.4 | 184 | 42.0 | 1273 | 1115 | 87.8 b | 141 a | 38.4 | 7.96 b | 25.5 | |
| + | 80.7 | 165 | 39.1 | 1249 | 1109 | 102.9 a | 119 b | 35.7 | 9.36 a | 24.6 | |
| − | 88.0 | 186 | 38.9 | 1213 | 1073 | 87.0 b | 128 | 35.2 | 8.48 | 25.9 | |
| + | 72.1 | 163 | 42.1 | 1309 | 1151 | 103.7 a | 132 | 38.9 | 8.84 | 24.3 | |
| − | − | 85.3 | 214 | 42.8 | 1212 | 1069 | 83.2 | 139 | 35.6 | 7.79 | 26.1 |
| + | 73.4 | 154 | 41.1 | 1334 | 1161 | 92.3 | 143 | 41.2 | 8.14 | 24.9 | |
| + | − | 90.7 | 157 | 35.1 | 1214 | 1077 | 90.8 | 118 | 34.8 | 9.17 | 25.6 |
| + | 70.7 | 172 | 43.0 | 1285 | 1141 | 115.0 | 121 | 36.6 | 9.54 | 23.7 | |
| SEM | 4.73 | 10.0 | 2.25 | 26.7 | 23.8 | 3.38 | 4.44 | 1.24 | 0.257 | 0.556 | |
| Significance (p-value) | |||||||||||
| Effects | ACV | 0.890 | 0.305 | 0.537 | 0.653 | 0.903 | 0.003 | 0.015 | 0.276 | 0.005 | 0.447 |
| DE | 0.111 | 0.241 | 0.507 | 0.084 | 0.122 | 0.001 | 0.647 | 0.146 | 0.413 | 0.180 | |
| Interaction | ACV × DE | 0.673 | 0.059 | 0.314 | 0.629 | 0.781 | 0.100 | 0.965 | 0.440 | 0.973 | 0.751 |
| Factors | Longitudinal m. Lamina Thickness [μm] | Circular m. Lamina Thickness [μm] | Submucosa Thickness [μm] | Mucosa Thickness [μm] | Villus Height [μm] | Villus Thickness [μm] | Crypt Depth [μm] | Crypt Width [μm] | Villus Height/Crypt Depth Ratio | Mucosal Surface Absorptive Area [μm2] | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ACV | DE | ||||||||||
| − | 82.9 | 201 | 35.1 | 845 | 698 | 96.1 | 108 b | 37.1 | 6.51 a | 15.8 | |
| + | 82.2 | 183 | 36.5 | 870 | 699 | 92.7 | 135 a | 35.9 | 5.17 b | 16.4 | |
| − | 90.6 a | 203 | 37.6 | 875 | 707 | 91.7 | 125 | 36.4 | 5.74 | 16.5 | |
| + | 74.5 b | 181 | 34.0 | 840 | 690 | 97.1 | 119 | 36.6 | 5.94 | 15.7 | |
| − | − | 90.5 | 207 | 36.5 | 910 | 747 | 99.0 | 114 | 38.8 | 6.56 | 16.2 |
| + | 75.4 | 195 | 33.6 | 780 | 650 | 93.2 | 102 | 35.3 | 6.45 | 15.4 | |
| + | − | 90.8 | 200 | 38.7 | 840 | 668 | 84.4 | 136 | 33.9 | 4.92 | 16.7 |
| + | 73.7 | 167 | 34.4 | 901 | 730 | 101.0 | 135 | 37.8 | 5.42 | 16.0 | |
| SEM | 3.45 | 8.70 | 2.02 | 23.1 | 22.6 | 3.63 | 4.17 | 0.925 | 0.230 | 0.482 | |
| p-value | |||||||||||
| Effects | ACV | 0.912 | 0.324 | 0.733 | 0.563 | 0.991 | 0.641 | 0.000 | 0.508 | 0.002 | 0.582 |
| DE | 0.022 | 0.210 | 0.406 | 0.425 | 0.701 | 0.464 | 0.293 | 0.912 | 0.595 | 0.463 | |
| Interaction | ACV × DE | 0.873 | 0.573 | 0.870 | 0.054 | 0.097 | 0.140 | 0.346 | 0.055 | 0.411 | 0.996 |
| Factors | ZO-1, Villi | Claudin-3, Crypts | |
|---|---|---|---|
| ACV | DE | (OD) | (OD) |
| − | 0.254 b | 0.238 a | |
| + | 0.339 a | 0.181 b | |
| − | 0.302 | 0.216 | |
| + | 0.291 | 0.203 | |
| − | − | 0.301 b | 0.223 ab |
| + | 0.208 c | 0.253 a | |
| + | − | 0.304 b | 0.210 b |
| + | 0.373 a | 0.153 c | |
| SEM | 0.016 | 0.010 | |
| p-value | |||
| Effects | ACV | 0.000 | 0.000 |
| DE | 0.529 | 0.210 | |
| Interaction | ACV × DE | 0.000 | 0.001 |
| Factors | IgY | IgM | IgA | TNF-α | IL-1β | IL-6 | Cp | Fb | |
|---|---|---|---|---|---|---|---|---|---|
| [mg/mL] | [mg/mL] | [µg/mL] | [pg/mL] | [pg/mL] | [pg/mL] | [µg/mL] | [mg/mL] | ||
| ACV | DE | ||||||||
| − | 10.1 | 6.56 b | 465 | 81.3 | 60.5 | 114.9 a | 77.9 | 5.28 | |
| + | 11.1 | 9.37 a | 430 | 68.2 | 44.3 | 92.6 b | 67.5 | 3.86 | |
| − | 10.7 | 6.59 a | 419 | 80.1 | 64.1 | 106.6 | 72.2 | 4.86 | |
| + | 10.5 | 9.34 b | 476 | 69.3 | 40.7 | 101.0 | 73.2 | 4.27 | |
| − | − | 12.2 ab | 6.39 b | 411 | 86.1 | 79.5 | 115.9 | 80.8 | 5.63 |
| + | 8.12 b | 6.72 b | 518 | 76.5 | 41.4 | 114.0 | 75.0 | 4.92 | |
| + | − | 9.31 ab | 6.79 b | 426 | 74.1 | 48.6 | 97.2 | 63.6 | 4.09 |
| + | 12.9 a | 11.96 a | 433 | 62.2 | 40.0 | 88.0 | 71.4 | 3.62 | |
| SEM | 0.803 | 0.707 | 30.2 | 4.78 | 6.74 | 4.61 | 3.39 | 0.398 | |
| p-value | |||||||||
| Effects | ACV | 0.511 | 0.014 | 0.580 | 0.186 | 0.208 | 0.015 | 0.142 | 0.087 |
| DE | 0.880 | 0.016 | 0.376 | 0.273 | 0.076 | 0.505 | 0.881 | 0.459 | |
| Interaction | ACV × DE | 0.018 | 0.031 | 0.432 | 0.901 | 0.247 | 0.664 | 0.327 | 0.881 |
| Factors | GLU | TG | TC | TP | ALT | AST | ALP | LDH | |
|---|---|---|---|---|---|---|---|---|---|
| ACV | DE | [mg/dL] | [mg/dL] | [mg/dL] | [g/dL] | [U/L] | [U/L] | [U/L] | [U/L] |
| − | 228 | 38.8 | 135 b | 2.58 b | 10.61 a | 241 | 3748 | 3041 | |
| + | 227 | 43.2 | 150.9 a | 2.85 a | 6.68 b | 251 | 4300 | 3008 | |
| − | 225 | 40.1 | 140.5 | 2.64 | 9.45 | 240 | 4132 | 2875 | |
| + | 230 | 41.9 | 145.4 | 2.79 | 7.83 | 252 | 3917 | 3174 | |
| − | − | 224 | 40.6 | 139.4 b | 2.66 b | 12.54 a | 239 | 3757 | 2789 |
| + | 231 | 37 | 130.6 b | 2.50 b | 8.67 b | 243 | 3739 | 3294 | |
| + | − | 225 | 39.6 | 141.6 ab | 2.62 b | 6.36 b | 242 | 4507 | 2962 |
| + | 229 | 46.8 | 160.2 a | 3.07 a | 7.00 b | 261 | 4094 | 3053 | |
| SEM | 3.30 | 1.91 | 3.79 | 2.58 | 0.728 | 7.12 | 212 | 172 | |
| p-value | |||||||||
| Effects | ACV | 0.944 | 0.257 | 0.022 | 0.003 | 0.002 | 0.501 | 0.223 | 0.927 |
| DE | 0.447 | 0.637 | 0.445 | 0.069 | 0.139 | 0.443 | 0.628 | 0.424 | |
| Interaction | ACV × DE | 0.834 | 0.168 | 0.044 | 0.001 | 0.045 | 0.633 | 0.657 | 0.577 |
| Factors | Acetic Acid | Propionic Acid | Isobutyric Acid | Butyric Acid | Isovaleric Acid | Valeric Acid | BCFAs | Total VFA | |
|---|---|---|---|---|---|---|---|---|---|
| [µmol/g] | [µmol/g] | [µmol/g] | [µmol/g] | [µmol/g] | [µmol/g] | [µmol/g] | [µmol/g] | ||
| ACV | DE | ||||||||
| − | 83.3 | 7.02 | 1.21 | 13.2 | 1.47 | 1.45 | 2.69 | 107.6 | |
| + | 77.3 | 7.27 | 1.07 | 11.8 | 1.27 | 1.27 | 2.33 | 99.9 | |
| − | 78.5 | 7.46 | 1.10 | 11.0 b | 1.30 | 1.30 | 2.40 | 100.6 | |
| + | 82.1 | 6.83 | 1.18 | 14.0 a | 1.44 | 1.42 | 2.62 | 107.0 | |
| − | − | 80.5 | 6.86 | 1.14 | 11.8 | 1.39 | 1.40 | 2.53 | 103.1 |
| + | 86.2 | 7.18 | 1.29 | 14.5 | 1.56 | 1.50 | 2.84 | 112.2 | |
| + | − | 76.4 | 8.06 | 1.05 | 10.1 | 1.22 | 1.20 | 2.27 | 98.1 |
| + | 78.1 | 6.48 | 1.08 | 13.5 | 1.32 | 1.34 | 2.40 | 101.8 | |
| SEM | 3.42 | 0.431 | 0.053 | 0.748 | 0.070 | 0.055 | 0.120 | 4.34 | |
| p-value | |||||||||
| Effects | ACV | 0.393 | 0.773 | 0.166 | 0.354 | 0.153 | 0.109 | 0.150 | 0.391 |
| DE | 0.605 | 0.477 | 0.424 | 0.048 | 0.330 | 0.292 | 0.359 | 0.478 | |
| Interaction | ACV × DE | 0.774 | 0.284 | 0.592 | 0.810 | 0.811 | 0.839 | 0.708 | 0.763 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arczewska-Włosek, A.; Świątkiewicz, S.; Tomaszewska, E.; Muszyński, S.; Dobrowolski, P.; Józefiak, D. Effects of Anticoccidial Vaccination and Taraxacum officinale Extract on the Growth Performance, Biochemical Parameters, Immunity, and Intestinal Morphology of Eimeria-Challenged Chickens. Life 2023, 13, 1927. https://doi.org/10.3390/life13091927
Arczewska-Włosek A, Świątkiewicz S, Tomaszewska E, Muszyński S, Dobrowolski P, Józefiak D. Effects of Anticoccidial Vaccination and Taraxacum officinale Extract on the Growth Performance, Biochemical Parameters, Immunity, and Intestinal Morphology of Eimeria-Challenged Chickens. Life. 2023; 13(9):1927. https://doi.org/10.3390/life13091927
Chicago/Turabian StyleArczewska-Włosek, Anna, Sylwester Świątkiewicz, Ewa Tomaszewska, Siemowit Muszyński, Piotr Dobrowolski, and Damian Józefiak. 2023. "Effects of Anticoccidial Vaccination and Taraxacum officinale Extract on the Growth Performance, Biochemical Parameters, Immunity, and Intestinal Morphology of Eimeria-Challenged Chickens" Life 13, no. 9: 1927. https://doi.org/10.3390/life13091927
APA StyleArczewska-Włosek, A., Świątkiewicz, S., Tomaszewska, E., Muszyński, S., Dobrowolski, P., & Józefiak, D. (2023). Effects of Anticoccidial Vaccination and Taraxacum officinale Extract on the Growth Performance, Biochemical Parameters, Immunity, and Intestinal Morphology of Eimeria-Challenged Chickens. Life, 13(9), 1927. https://doi.org/10.3390/life13091927