


Effects of Four Organic Carbon Sources on the Growth and Astaxanthin Accumulation of Haematococcus lacustris


 ,
, 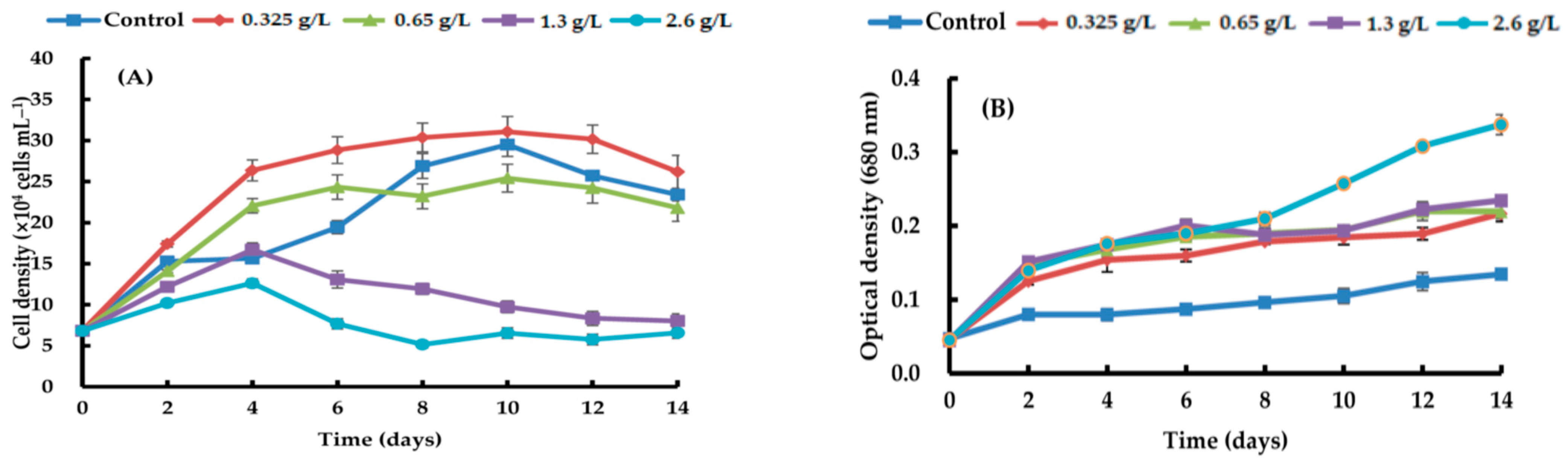{kind=link}
{kind=link}
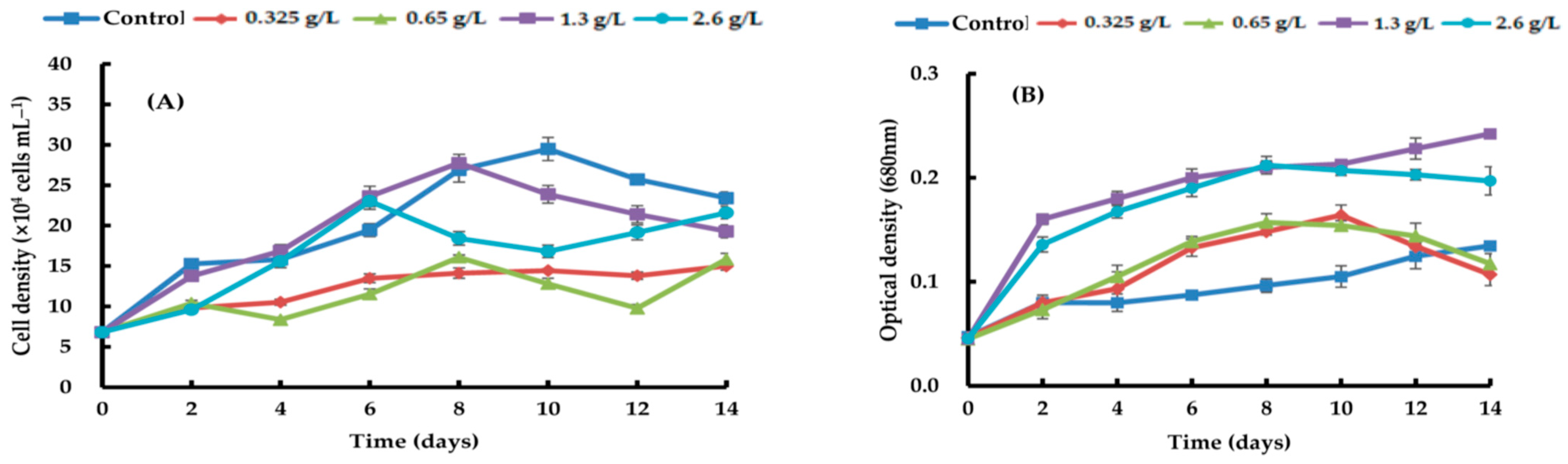{kind=link}
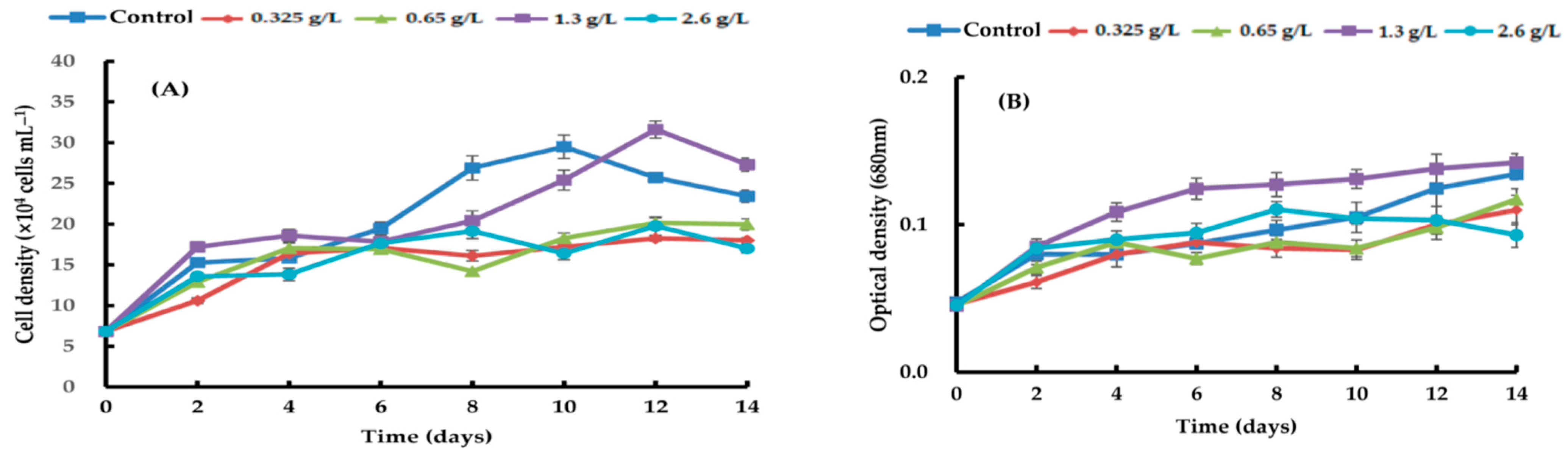{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Organic Carbon Sources
2.3. Analysis of Cell Growth
2.4. Analysis of Total Carotenoid Content
2.5. Analysis of Astaxanthin Content
2.6. Statistical Analysis
3. Results
3.1. Green Vegetative Stage of H. lacustris
3.2. Astaxanthin Accumulation According to Growth Stage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquac. 2018, 10, 738–773. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, C.S.; Lee, Y.J. Astaxanthin protects against MPTP/MPP+-induced mitochondrial dysfunction and ROS production in vivo and in vitro. Food Chem. Toxicol. 2011, 49, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Wang, W.; Wu, B.; Ji, S.; Xia, Q. Preparation and in vitro characterization studies of astaxanthin-loaded nanostructured lipid carriers with antioxidant properties. J. Biomater. Appl. 2023, 38, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Ahirwar, A.; Singh, S.; Lodhi, R.; Lodhi, A.; Rai, A.; Jadhav, D.A.; Harish; Varjani, S.; Singh, G.; et al. Astaxanthin as a King of Ketocarotenoids: Structure, Synthesis, Accumulation, Bioavailability and Antioxidant Properties. Mar. Drugs 2023, 21, 176. [Google Scholar] [CrossRef] [PubMed]
- Higuera-Ciapara, I.; Felix-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Onorato, C.; Rösch, C. Comparative life cycle assessment of astaxanthin production with Haematococcus pluvialis in different photobioreactor technologies. Algal Res. 2020, 50, 102005. [Google Scholar] [CrossRef]
- Li, Y.; Sommerfeld, M.; Chen, F.; Hu, Q. Effect of photon flux densities on regulation of carotenogenesis and cell viability of Haematococcus pluvialis (Chlorophyceae). J. Appl. Phycol. 2010, 22, 253–263. [Google Scholar] [CrossRef]
- Chekanov, K. Diversity and Distribution of Carotenogenic Algae in Europe: A Review. Mar. Drugs 2023, 21, 108. [Google Scholar] [CrossRef]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other nutrients from Haematococcus pluvialis—Multifunctional applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Yu, W.; Liu, J. Astaxanthin isomers: Selective distribution and isomerization in aquatic animals. Aquaculture 2020, 520, 734915. [Google Scholar] [CrossRef]
- Shah, M.M.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Hazen, T.E. The life history of Sphaerella lacustris (Haematococcus pluvialis). Mem. Torrey Bot. Club 1899, 6, 211–246. [Google Scholar]
- Elliott, A.M. Morphology and life history of Haematococcus pluvialis. Archiv. Protistekunde 1934, 82, 250–272. [Google Scholar]
- Wayama, M.; Ota, S.; Matsuura, H.; Nango, N.; Hirata, A.; Kawano, S. Three-dimensional ultrastructural study of oil and astaxanthin accumulation during encystment in the green alga Haematococcus pluvialis. PLoS ONE 2013, 8, e53618. [Google Scholar] [CrossRef] [PubMed]
- Imamoglu, E.; Sukan, F.V.; Dalay, M.C. Effect of different culture media and light intensities on growth of Haematococcus pluvialis. Int. J. Nat. Eng. Sci. 2007, 1, 5–9. [Google Scholar]
- Yu, X.; Ye, X.; Hu, C.; Xu, N.; Sun, X. Sodium acetate can promote the growth and astaxanthin accumulation in the unicellular green alga Haematococcus pluvialis as revealed by a proteomics approach. J. Oceanol. Limnol. 2022, 40, 2052–2067. [Google Scholar] [CrossRef]
- Chu, F.; Cheng, J.; Zhang, X.; Ye, Q.; Zhou, J. Enhancing lipid production in microalgae Chlorella PY-ZU1 with phosphorus excess and nitrogen starvation under 15% CO2 in a continuous two-step cultivation process. Chem. Eng. J. 2019, 375, 121912. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Oslan, S.N.; Mohamad, R.; Tan, J.S.; Yusoff, A.H.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Bioprocess Strategy of Haematococcus lacustris for Biomass and Astaxanthin Production Keys to Commercialization: Perspective and Future Direction. Fermentation 2022, 8, 179. [Google Scholar] [CrossRef]
- Chekanov, K.; Schastnaya, E.; Solovchenko, A.; Lobakova, E. Effects of CO2 enrichment on primary photochemistry, growth and astaxanthin accumulation in the chlorophyte Haematococcus pluvialis. J. Photochem. Photobiol. B 2017, 171, 58–66. [Google Scholar] [CrossRef]
- Wu, K.; Ying, K.; Liu, L.; Zhou, J.; Cai, Z. High irradiance compensated with CO2 enhances the efficiency of Haematococcus lacustris growth. Biotechnol. Rep. 2020, 26, e00444. [Google Scholar] [CrossRef]
- Wu, K.; Ying, K.; Zhou, J.; Liu, D.; Liu, L.; Tao, Y.; Hanotu, J.; Zhu, X.; Cai, Z. Optimizing the growth of Haematococcus pluvialis based on a novel microbubble-driven photobioreactor. iScience 2021, 24, 103461. [Google Scholar] [CrossRef]
- Rushan, N.H.; Said, F.M. The effect of culture medium on the oil yield and fatty acid methyl ester of freshwater microalgae Chlorella vulgaris. Chem. Eng. Commun. 2021, 208, 592–600. [Google Scholar] [CrossRef]
- Pang, N.; Chen, S. Effects of C5 organic carbon and light on growth and cell activity of Haematococcus pluvialis under mixotrophic conditions. Algal Res. 2017, 21, 227–235. [Google Scholar] [CrossRef]
- Pang, N.; Xie, Y.; Oung, H.M.O.; Sonawane, B.V.; Fu, X.; Kirchhoff, H.; Chen, S. Regulation and stimulation of photosynthesis of mixotrophically cultured Haematococcus pluvialis by ribose. Algal Res. 2019, 39, 101443. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Cheng, J.; Li, K.; Yang, Z.; Zhou, J.; Cen, K. Enhancing the growth rate and astaxanthin yield of Haematococcus pluvialis by nuclear irradiation and high concentration of carbon dioxide stress. Bioresour. Technol. 2016, 204, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Olguín, E.J.; Sánchez-Galván, G.; Arias-Olguín, I.I.; Melo, F.J.; González-Portela, R.E.; Cruz, L.; Adessi, A. Microalgae-Based Biorefineries: Challenges and future trends to produce carbohydrate enriched biomass, High-added value products and bioactive compounds. Biology 2022, 11, 1146. [Google Scholar] [CrossRef]
- Chakraborty, K.; Krishnan, S.; Chakraborty, R.D. Optimization of Pigment Extraction from Aristeus alcocki Shell Wastes via Different Solvent Systems. J. Aquat. Food Prod. Technol. 2023, 32, 185–195. [Google Scholar] [CrossRef]
- Ren, Y.; Deng, J.; Huang, J.; Wu, Z.; Yi, L.; Bi, Y.; Chen, F. Using green alga Haematococcus pluvialis for astaxanthin and lipid co-production: Advances and outlook. Bioresour. Technol. 2021, 340, 125736. [Google Scholar] [CrossRef]
- Ma, R.; Tao, X.; Chua, E.T.; Ho, S.H.; Shi, X.; Liu, L.; Chen, J. Enhancing astaxanthin production in Haematococcus pluvialis QLD by a pH steady NaHCO3-CO2-C/NH4Cl-N culture system. Algal Res. 2022, 64, 102697. [Google Scholar] [CrossRef]
- Joun, J.; Sirohi, R.; Sim, S.J. The effects of acetate and glucose on carbon fixation and carbon utilization in mixotrophy of Haematococcus pluvialis. Bioresour. Technol. 2023, 367, 128218. [Google Scholar] [CrossRef] [PubMed]
- Pang, N.; Gu, X.; Fu, X.; Chen, S. Effects of gluconate on biomass improvement and light stress tolerance of Haematococcus pluvialis in mixotrophic culture. Algal Res. 2019, 43, 101647. [Google Scholar] [CrossRef]
- Du, F.; Hu, C.; Sun, X.; Xu, N. Transcriptome analysis reveals pathways responsible for the promoting effect of sucrose on astaxanthin accumulation in Haematococcus pluvialis under high light condition. Aquaculture 2021, 530, 735757. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, L.; Zhao, J.; Liu, J. Enhancement of astaxanthin accumulation in Haematococcus pluvialis by exogenous oxaloacetate combined with nitrogen deficiency. Bioresour. Technol. 2022, 345, 126484. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Hejazi, M.A.; Hashemi, M. Supplementation with polyalcohols and sequential mixotrophy dilution photoinduction strategy boost the accumulation of astaxanthin by Haematococcus pluvialis. Aquaculture 2019, 511, 734225. [Google Scholar] [CrossRef]
- Niizawa, I.; Espinaco, B.Y.; Leonardi, J.R.; Heinrich, J.M.; Sihufe, G.A. Enhancement of astaxanthin production from Haematococcus pluvialis under autotrophic growth conditions by a sequential stress strategy. Prep. Biochem. Biotechnol. 2018, 48, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Orosa, M.; Franqueira, D.; Cid, A.; Abalde, J.J.B.T. Analysis and enhancement of astaxanthin accumulation in Haematococcus pluvialis. Bioresour. Technol. 2005, 96, 373–378. [Google Scholar] [CrossRef]
- Kobayashi, M.; Kakizono, T.; Nagai, S. Enhanced carotenoid biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular alga, Haematococcus pluvialis. Appl. Environ. Microbiol. 1993, 59, 867–873. [Google Scholar] [CrossRef]
- Droop, M.R. Carotenogenesis in Haematococcus pluvialis. Nature 1955, 175, 42. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Huisman, J.M.; Osborn, A. Culture of the astaxanthin-producing green alga Haematococcus pluvialis 1. Effects of nutrients on growth and cell type. J. Appl. Phycol. 1991, 3, 295–304. [Google Scholar] [CrossRef]
- Kakizono, T.; Kobayashi, M.; Nagai, S. Effect of carbon/nitrogen ratio on encystment accompanied with astaxanthin formation in a green alga, Haematococcus pluvialis. J. Ferment. Bioeng. 1992, 74, 403–405. [Google Scholar] [CrossRef]
- Steinbrenner, J.; Linden, H. Regulation of two carotenoid biosynthesis genes coding for phytoene synthase and carotenoid hydroxylase during stress-induced astaxanthin formation in the green alga Haematococcus pluvialis. Plant Physiol. 2001, 125, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, H.P.; Yu, X. The associative induction of succinic acid and hydrogen sulfide for high-producing biomass, astaxanthin and lipids in Haematococcus pluvialis. Bioresour. Technol. 2022, 358, 127397. [Google Scholar] [CrossRef]
- Minyuk, G.S.; Chelebieva, E.S.; Chubchikova, I.N.; Dantsyuk, N.V.; Drobetskaya, I.V.; Sakhon, E.G.; Chekanov, K.; Solovchenko, A.E. Stress-induced secondary carotenogenesis in Coelastrella rubescens (Scenedesmaceae, Chlorophyta), a producer of value-added keto-carotenoids. Algae 2017, 32, 245–259. [Google Scholar] [CrossRef]
- Andruleviciute, V.; Makareviciene, V.; Skorupskaite, V.; Gumbyte, M. Biomass and oil content of Chlorella sp., Haematococcus sp., Nannochloris sp. and Scenedesmus sp. under mixotrophic growth conditions in the presence of technical glycerol. J. Appl. Phycol. 2014, 26, 83–90. [Google Scholar] [CrossRef]
- Dechatiwongse, P.; Choorit, W. Mixotrophic Growth of Astaxanthin-Rich Alga Haematococcus pluvialis using Refined Crude Glycerol as Carbon Substrate: Batch and Fed-Batch Cultivations. Walailak J. Sci. Technol. 2021, 18, 7354-20. [Google Scholar] [CrossRef]
- Poddar, N.; Sen, R.; Martin, G.J.O. Glycerol and nitrate utilisation by marine microalgae Nannochloropsis salina and Chlorella sp. and associated bacteria during mixotrophic and heterotrophic growth. Algal Res. 2018, 33, 298–309. [Google Scholar] [CrossRef]
- Paranjape, K.; Leite, G.B.; Hallenbeck, P.C. Effect of nitrogen regime on microalgal lipid production during mixotrophic growth with glycerol. Bioresour. Technol. 2016, 214, 778–786. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef]
- Das, P.; Lei, W.; Aziz, S.S.; Obbard, J.P. Enhanced algae growth in both phototrophic and mixotrophic culture under blue light. Bioresour. Technol. 2011, 102, 3883–3887. [Google Scholar] [CrossRef]
- Da Silva, G.P.; Mack, M.; Contiero, J. Glycerol: A promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 2009, 27, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Lee, S.M. Biomass and oil content of microalgae under mixotrophic conditions. Environ. Eng. Res. 2015, 20, 25–32. [Google Scholar] [CrossRef]
- Sun, H.; Liu, B.; Lu, X.; Cheng, K.W.; Chen, F. Staged cultivation enhances biomass accumulation in the green growth phase of Haematococcus pluvialis. Bioresour. Technol. 2017, 233, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.; Tran, S.; Nguyen, P.; Mai, T. Growth, carotenoid production, antioxidant capacity and lipid accumulation of Haematococcus sp. under different light intensities. Am. J. Plant Biol. 2017, 2, 142–147. [Google Scholar]
- Xu, Y.F.; Létisse, F.; Absalan, F.; Lu, W.; Kuznetsova, E.; Brown, G.; Rabinowitz, J.D. Nucleotide degradation and ribose salvage in yeast. Mol. Syst. Biol. 2013, 9, 665. [Google Scholar] [CrossRef]
- Tonon, T.; Li, Y.; McQueen-Mason, S. Mannitol biosynthesis in algae: More widespread and diverse than previously thought. New Phytol. 2017, 213, 1573–1579. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byeon, H.; An, Y.; Kim, T.; Rayamajhi, V.; Lee, J.; Shin, H.; Jung, S. Effects of Four Organic Carbon Sources on the Growth and Astaxanthin Accumulation of Haematococcus lacustris. Life 2024, 14, 29. https://doi.org/10.3390/life14010029
Byeon H, An Y, Kim T, Rayamajhi V, Lee J, Shin H, Jung S. Effects of Four Organic Carbon Sources on the Growth and Astaxanthin Accumulation of Haematococcus lacustris. Life. 2024; 14(1):29. https://doi.org/10.3390/life14010029
Chicago/Turabian StyleByeon, Huijeong, Yunji An, Taesoo Kim, Vijay Rayamajhi, Jihyun Lee, HyunWoung Shin, and SangMok Jung. 2024. "Effects of Four Organic Carbon Sources on the Growth and Astaxanthin Accumulation of Haematococcus lacustris" Life 14, no. 1: 29. https://doi.org/10.3390/life14010029
APA StyleByeon, H., An, Y., Kim, T., Rayamajhi, V., Lee, J., Shin, H., & Jung, S. (2024). Effects of Four Organic Carbon Sources on the Growth and Astaxanthin Accumulation of Haematococcus lacustris. Life, 14(1), 29. https://doi.org/10.3390/life14010029