
Approaches to Characterize and Quantify Extracellular Vesicle Surface Conjugation Efficiency



Abstract
:1. Introduction
2. Conjugation Methods
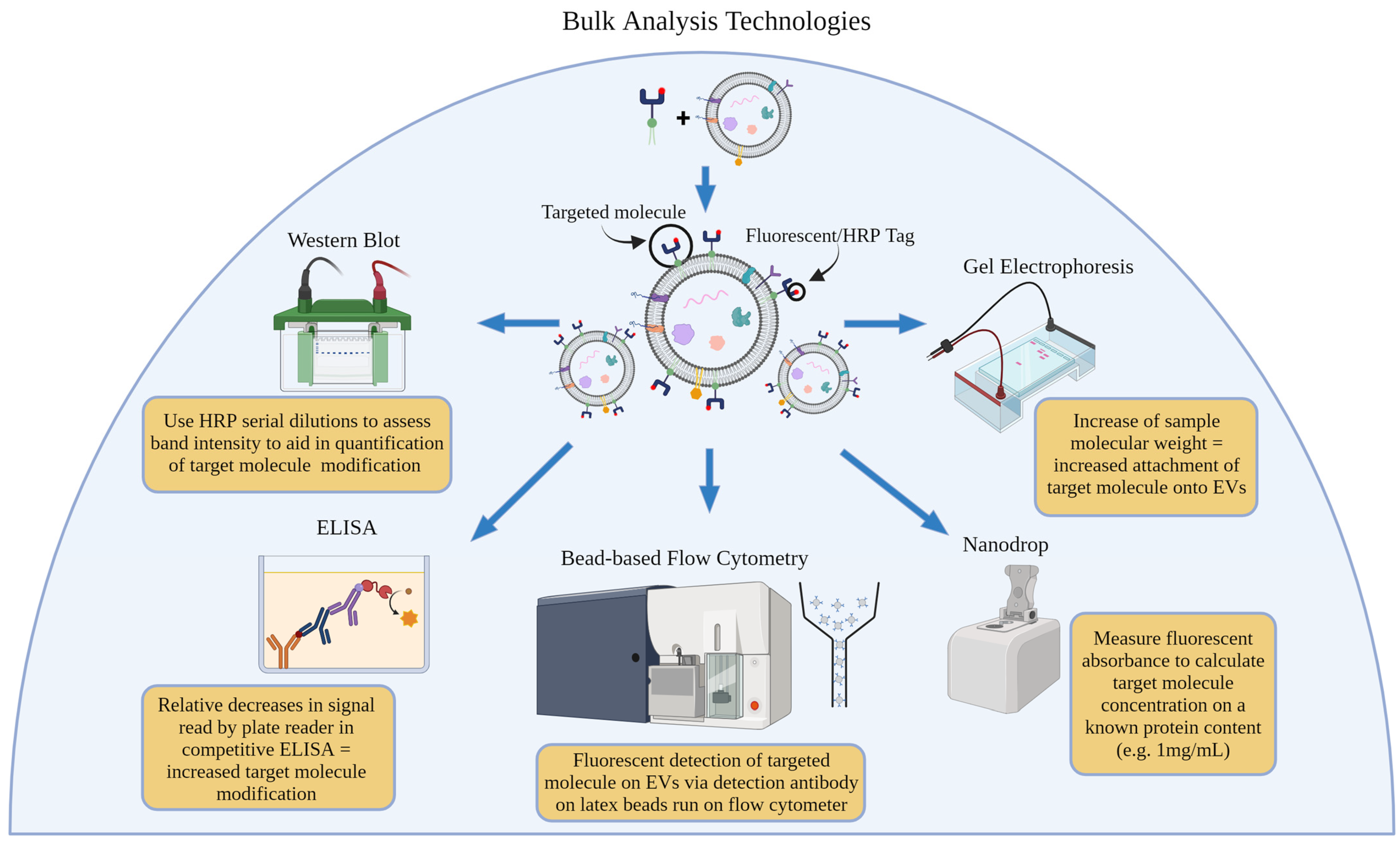
3. Bulk Analysis Technologies
3.1. Western Blot
Western Blot (Serial Dilution of HRP)
3.2. Bead-Based Flow Cytometry
3.3. Absorbance
3.3.1. Nanodrop/BCA
3.3.2. Fluorescence Correlation Spectroscopy/Plate Reader/ELISA
3.3.3. Gel Electrophoresis
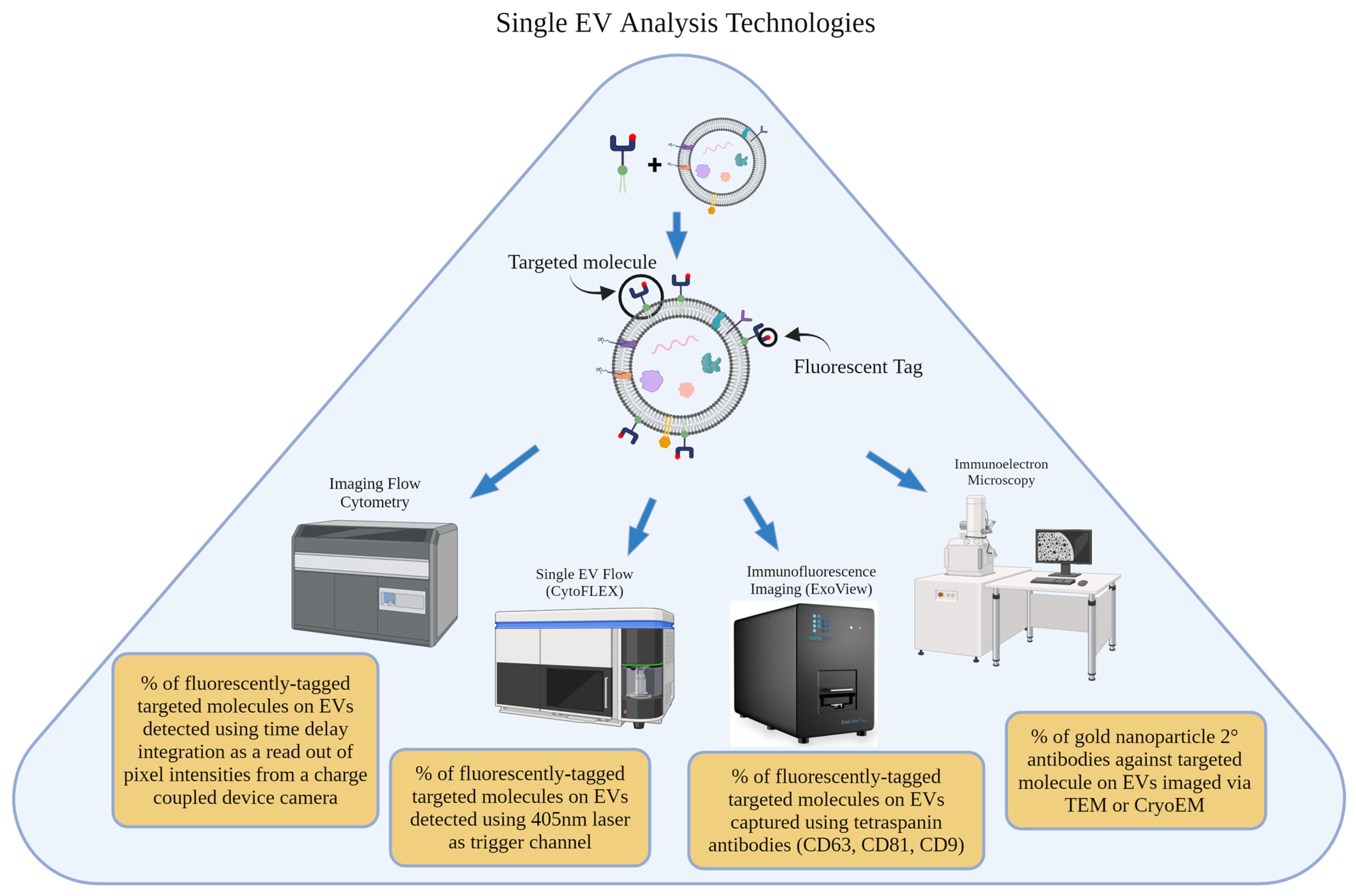
4. Single-Nanovesicle Analysis Technologies
4.1. Single EV Flow Cytometry
4.2. ExoView/Single Particle Interferometric Reflectance Imaging (SP-IRIS)
4.3. Immunoelectron Microscopy/Immunogold Labeling
5. Improved Targeting/Uptake/Biodistribution
6. How Conjugation Efficiency Calculations Differ Based on Confirmation Method
6.1. Sampling of Different Total Populations
6.2. Detection Threshold of Technology
7. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Margolis, L.; Sadovsky, Y. The biology of extracellular vesicles: The known unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wang, Y.; Xia, X.; Zheng, J.C. Exosome engineering: Current progress in cargo loading and targeted delivery. NanoImpact 2020, 20, 100261. [Google Scholar] [CrossRef]
- Burrello, J.; Monticone, S.; Gai, C.; Gomez, Y.; Kholia, S.; Camussi, G. Stem Cell-Derived Extracellular Vesicles and Immune-Modulation. Front. Cell Dev. Biol. 2016, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Kholia, S.; Ranghino, A.; Garnieri, P.; Lopatina, T.; Deregibus, M.C.; Rispoli, P.; Brizzi, M.F.; Camussi, G. Extracellular vesicles as new players in angiogenesis. Vascul. Pharmacol. 2016, 86, 64–70. [Google Scholar] [CrossRef] [PubMed]
- De Jong, O.G.; van Balkom, B.W.M.; Schiffelers, R.M.; Bouten, C.V.C.; Verhaar, M.C. Extracellular Vesicles: Potential Roles in Regenerative Medicine. Front. Immunol. 2014, 5, 608. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tan, X.; Tan, Y.; Li, Q.; Ma, J.; Wang, G. Mesenchymal Stem Cell Derived Exosomes in Cancer Progression, Metastasis and Drug Delivery: A Comprehensive Review. J. Cancer 2018, 9, 3129–3137. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Vermesh, O.; Mani, V.; Ge, T.J.; Madsen, S.J.; Sabour, A.; Hsu, E.-C.; Gowrishankar, G.; Kanada, M.; Jokerst, J.V.; et al. The Exosome Total Isolation Chip. ACS Nano 2017, 11, 10712–10723. [Google Scholar] [CrossRef]
- Lai, C.P.; Mardini, O.; Ericsson, M.; Prabhakar, S.; Maguire, C.A.; Chen, J.W.; Tannous, B.A.; Breakefield, X.O. Dynamic biodistribution of extracellular vesicles in vivo using a multimodal imaging reporter. ACS Nano 2014, 8, 483–494. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and delivery efficiency of unmodified tumor-derived exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Grange, C.; Tapparo, M.; Bruno, S.; Chatterjee, D.; Quesenberry, P.J.; Tetta, C.; Camussi, G. Biodistribution of mesenchymal stem cell-derived extracellular vesicles in a model of acute kidney injury monitored by optical imaging. Int. J. Mol. Med. 2014, 33, 1055. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.C.; Kim, S.R.; Yoon, Y.J.; Park, K.-S.; Kim, J.H.; Lee, J.; Kim, O.Y.; Choi, E.-J.; Kim, D.-K.; Choi, D.-S.; et al. In vivo Kinetic Biodistribution of Nano-Sized Outer Membrane Vesicles Derived from Bacteria. Small 2015, 11, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Nishikawa, M.; Shinotsuka, H.; Matsui, Y.; Ohara, S.; Imai, T.; Takakura, Y. Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J. Biotechnol. 2013, 165, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ramasubramanian, L.; Kumar, P.; Wang, A. Engineering Extracellular Vesicles as Nanotherapeutics for Regenerative Medicine. Biomolecules 2019, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, M.K.; Tan, M.; Peng, B.; Yang, Y.; Sethi, G.; Pirisinu, M.; Le, M.T. New approaches in extracellular vesicle engineering for improving the efficacy of anti-cancer therapies. Semin. Cancer Biol. 2021, 74, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhao, Y.; Xue, F.; Zheng, Y.; Huang, H.; Wang, W.; Chang, Y.; Yang, H.; Zhang, J. Exosomal DNA Aptamer Targeting α-Synuclein Aggregates Reduced Neuropathological Deficits in a Mouse Parkinson’s Disease Model. Mol. Ther. Nucleic Acids 2019, 17, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Rayamajhi, S.; Aryal, S. Surface functionalization strategies of extracellular vesicles. J. Mater. Chem. B 2020, 8, 4552–4569. [Google Scholar] [CrossRef]
- Vandergriff, A.; Huang, K.; Shen, D.; Hu, S.; Hensley, M.T.; Thomas, G. Theragnostic Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics 2018, 8, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pol, J.; Gosselet, F.; Duban-Deweer, S.; Pottiez, G.; Karamanos, Y. Targeting and Crossing the Blood-Brain Barrier with Extracellular Vesicles. Cells 2020, 9, 851. [Google Scholar] [CrossRef] [PubMed]
- Iavorovschi, A.M.; Wang, A. Engineering mesenchymal stromal/stem cell-derived extracellular vesicles with improved targeting and therapeutic efficiency for the treatment of central nervous system disorders. Neural Regen. Res. 2020, 15, 2235–2236. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, X.; Liang, Y.; Wen, C.; Ouyang, K.; Huang, J.; Xiao, Y.; Deng, X.; Xia, J.; Duan, L. Osteoclast-targeted delivery of anti-miRNA oligonucleotides by red blood cell extracellular vesicles. J. Ophthalmol. Clin. Res. 2023, 358, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.A.; Aleza, C.G.; Roffler, S.R.; van Solinge, W.W.; Vader, P.; Schiffelers, R.M. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting. J. Extracell. Vesicles 2016, 5, 31053. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, L.; Zhu, C.; Zheng, Q.; Wang, G.; Tong, J.; Fang, Y.; Xia, Y.; Cheng, G.; He, X.; et al. Aptamer-conjugated extracellular nanovesicles for targeted drug delivery. Cancer Res. 2018, 78, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, M.K.; Pirisinu, M.; Yang, Y.; Peng, B.; Pham, T.T.; Lee, C.Y.; Tan, M.; Vu, L.T.; Dang, X.T.T.; Pham, T.C.; et al. Surface-engineered extracellular vesicles for targeted delivery of therapeutic RNAs and peptides for cancer therapy. Theranostics 2022, 12, 3288–3315. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Lu, L.; Song, H.; Duan, Y.; Chen, J.; Carney, R.; Li, J.J.; Zhou, P.; Nolta, J.; Lam, K.S.; et al. Engineered extracellular vesicles with high collagen-binding affinity present superior in situ retention and therapeutic efficacy in tissue repair. Theranostics 2022, 12, 6021–6037. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Yoshida, S.; Sawada, S.I.; Sasaki, Y.; Akiyoshi, K. Fusogenic Hybrid Extracellular Vesicles with PD-1 Membrane Proteins for the Cytosolic Delivery of Cargos. Cancers 2022, 14, 2635. [Google Scholar] [CrossRef]
- Pham, T.C.; Jayasinghe, M.K.; Pham, T.T.; Yang, Y.; Wei, L.; Usman, W.M.; Chen, H.; Pirisinu, M.; Gong, J.; Kim, S.; et al. Covalent conjugation of extracellular vesicles with peptides and nanobodies for targeted therapeutic delivery. J. Extracell. Vesicles 2021, 10, e12057. [Google Scholar] [CrossRef]
- Greene, M.K.; Richards, D.A.; Nogueira, J.C.F.; Campbell, K.; Smyth, P.; Fernández, M.; Scott, C.J.; Chudasama, V. Forming next-generation antibody–nanoparticle conjugates through the oriented installation of non-engineered antibody fragments. Chem. Sci. 2017, 9, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Florinas, S.; Teitgen, A.; Xu, Z.-Q.; Gao, C.; Wu, H.; Kataoka, K.; Cabral, H.; Christie, R.J. Controlled Fab installation onto polymeric micelle nanoparticles for tuned bioactivity. Sci. Technol. Adv. Mater. 2017, 18, 666–680. [Google Scholar] [CrossRef]
- Choi, E.S.; Song, J.; Kang, Y.Y.; Mok, H. Mannose-Modified Serum Exosomes for the Elevated Uptake to Murine Dendritic Cells and Lymphatic Accumulation. Macromol. Biosci. 2019, 19, e1900042. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, E.; Abnous, K.; Farzad, S.A.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Targeted doxorubicin-loaded mesenchymal stem cells-derived exosomes as a versatile platform for fighting against colorectal cancer. Life Sci. 2020, 261, 118369. [Google Scholar] [CrossRef] [PubMed]
- Yerneni, S.S.; Lathwal, S.; Shrestha, P.; Shirwan, H.; Matyjaszewski, K.; Weiss, L.; Yolcu, E.S.; Campbell, P.G.; Das, S.R. Rapid On-Demand Extracellular Vesicle Augmentation with Versatile Oligonucleotide Tethers. ACS Nano 2019, 13, 10555–10565. [Google Scholar] [CrossRef] [PubMed]
- Kirpotin, D.; Park, J.W.; Hong, K.; Zalipsky, S.; Li, W.-L.; Carter, P.; Benz, A.C.C.; Papahadjopoulos, D. Sterically stabilized anti-HER2 immunoliposomes: Design and targeting to human breast cancer cells in vitro. Biochemistry 1997, 36, 66–75. [Google Scholar] [CrossRef]
- Chen, C.; Sun, M.; Liu, X.; Wu, W.; Su, L.; Li, Y.; Liu, G.; Yan, X. General and mild modification of food-derived extracellular vesicles for enhanced cell targeting. Nanoscale 2021, 13, 3061–3069. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Ohno, S.I.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically Injected Exosomes Targeted to EGFR Deliver Antitumor MicroRNA to Breast Cancer Cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef]
- Kooijmans, S.A.A.; Gitz-Francois, J.J.J.M.; Schiffelers, R.M.; Vader, P. Recombinant phosphatidylserine-binding nanobodies for targeting of extracellular vesicles to tumor cells: A plug-and-play approach. Nanoscale 2018, 10, 2413–2426. [Google Scholar] [CrossRef]
- Antes, T.J.; Middleton, R.C.; Luther, K.M.; Ijichi, T.; Peck, K.A.; Liu, W.J.; Valle, J.; Echavez, A.K.; Marbán, E. Targeting extracellular vesicles to injured tissue using membrane cloaking and surface display. J. Nanobiotechnol. 2018, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Altinoglu, S.; Takeda, Y.S.; Xu, Q. Integrating Protein Engineering and Bioorthogonal Click Conjugation for Extracellular Vesicle Modulation and Intracellular Delivery. PLoS ONE 2015, 10, e0141860. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.A.; Fliervoet, L.A.L.; van der Meel, R.; Fens, M.H.A.M.; Heijnen, H.F.G.; van Bergen En Henegouwen, P.M.P.; Vader, P.; Schiffelers, R.M. PEGylated and targeted extracellular vesicles display enhanced cell specificity and circulation time. J. Control. Release 2016, 224, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhang, H.-X.; He, C.-P.; Fan, S.; Zhu, Y.-L.; Qi, C.; Huang, N.-P.; Xiao, Z.-D.; Lu, Z.-H.; Tannous, B.A.; et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials 2018, 150, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Sajeesh, S.; Camardo, A.; Dahal, S.; Ramamurthi, A. Surface-Functionalized Stem Cell-Derived Extracellular Vesicles for Vascular Elastic Matrix Regenerative Repair. Mol. Pharm. 2023, 20, 2801–2813. [Google Scholar] [CrossRef]
- Kanada, M.; Linenfelser, L.; Cox, E.; Gilad, A.A. A Dual-Reporter Platform for Screening Tumor-Targeted Extracellular Vesicles. Pharmaceutics 2022, 14, 475. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Zhang, L.; Wang, S.; Liu, Y.; Wu, C.; Cui, C.; Sun, H.; Shi, M.; Jiang, Y.; Li, L.; et al. Molecular Recognition-Based DNA Nanoassemblies on the Surfaces of Nanosized Exosomes. J. Am. Chem. Soc. 2017, 139, 5289–5292. [Google Scholar] [CrossRef]
- Morishita, M.; Takahashi, Y.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Exosome-based tumor antigens–adjuvant co-delivery utilizing genetically engineered tumor cell-derived exosomes with immunostimulatory CpG DNA. Biomaterials 2016, 111, 55–65. [Google Scholar] [CrossRef]
- Smyth, T.; Petrova, K.; Payton, N.M.; Persaud, I.; Redzic, J.S.; Graner, M.W.; Smith-Jones, P.; Anchordoquy, T.J. Surface functionalization of exosomes using click chemistry. Bioconjug. Chem. 2014, 25, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.W.; Li, F.-X.; Liu, Y.-W.; Rao, S.-S.; Yin, H.; Huang, J.; Chen, C.-Y.; Hu, Y.; Zhang, Y.; Tan, Y.-J.; et al. Aptamer-functionalized exosomes from bone marrow stromal cells target bone to promote bone regeneration. Nanoscale 2019, 11, 20884–20892. [Google Scholar] [CrossRef]
- Mukhopadhya, A.; Tsiapalis, D.; McNamee, N.; Talbot, B.; O’Driscoll, L. Doxorubicin Loading into Milk and Mesenchymal Stem Cells’ Extracellular Vesicles as Drug Delivery Vehicles. Pharmaceutics 2023, 15, 718. [Google Scholar] [CrossRef] [PubMed]
- Kolb, K.B.; Finn, H.C.; Sharpless, M.G. Click Chemistry: Diverse Chemical Function from a Few Good Reactions. Angew. Chemie Int. Ed. 2001, 40, 2004–2021. [Google Scholar] [CrossRef]
- Chen, C.; Wang, J.; Sun, M.; Li, J.; Wang, H.M.D. Toward the next-generation phyto-nanomedicines: Cell-derived nanovesicles (CDNs) for natural product delivery. Biomed. Pharmacother. 2022, 145, 112416. [Google Scholar] [CrossRef] [PubMed]
- Kowal, E.J.K.; Ter-Ovanesyan, D.; Regev, A.; Church, G.M. Extracellular Vesicle Isolation and Analysis by Western Blotting. Methods Mol. Biol. 2017, 1660, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Bostancioglu, R.B.; Welsh, J.A.; Zickler, A.M.; Murke, F.; Corso, G.; Felldin, U.; Hagey, D.W.; Evertsson, B.; Liang, X.-M.; et al. Systematic Methodological Evaluation of a Multiplex Bead-Based Flow Cytometry Assay for Detection of Extracellular Vesicle Surface Signatures. Front. Immunol. 2018, 9, 1326. [Google Scholar] [CrossRef] [PubMed]
- Suárez, H.; Gámez-Valero, A.; Reyes, R.; López-Martín, S.; Rodríguez, M.J.; Carrascosa, J.L.; Cabañas, C.; Borràs, F.E.; Yáñez-Mó, M. A bead-assisted flow cytometry method for the semi-quantitative analysis of Extracellular Vesicles. Sci. Rep. 2017, 7, 11271. [Google Scholar] [CrossRef] [PubMed]
- Florinas, S.; Liu, M.; Fleming, R.; Van Vlerken-Ysla, L.; Ayriss, J.; Gilbreth, R.; Dimasi, N.; Gao, C.; Wu, H.; Xu, Z.-Q.; et al. A Nanoparticle Platform to Evaluate Bioconjugation and Receptor-Mediated Cell Uptake Using Cross-Linked Polyion Complex Micelles Bearing Antibody Fragments. Biomacromolecules 2016, 17, 1818–1833. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.; Lantz, L.M.; Fowlkes, B.J.; Schmid, I.; Giorgi, J.V. Preparation of Cells and Reagents for Flow Cytometry. Curr. Protoc. Immunol. 2001, 44, 5.3.1–5.3.24. [Google Scholar] [CrossRef] [PubMed]
- Van der Pol, E.; Coumans, F.A.W.; Grootemaat, A.E.; Gardiner, C.; Sargent, I.L.; Harrison, P.; Sturk, A.; van Leeuwen, T.G.; Nieuwland, R. Particle size distribution of exosomes and microvesicles determined by transmission electron microscopy, flow cytometry, nanoparticle tracking analysis, and resistive pulse sensing. J. Thromb. Haemost. 2014, 12, 1182–1192. [Google Scholar] [CrossRef]
- Lannigan, J.; Erdbruegger, U. Imaging flow cytometry for the characterization of extracellular vesicles. Methods 2017, 112, 55–67. [Google Scholar] [CrossRef]
- Morales-Kastresana, A.; Jones, J.C. Flow Cytometric Analysis of Extracellular Vesicles; Humana Press: New York, NY, USA, 2017; pp. 215–225. [Google Scholar]
- Morales-Kastresana, A.; Telford, B.; Musich, T.A.; McKinnon, K.; Clayborne, C.; Braig, Z.; Rosner, A.; Demberg, T.; Watson, D.C.; Karpova, T.S.; et al. Labeling extracellular vesicles for nanoscale flow cytometry. Sci. Rep. 2017, 7, 1878. [Google Scholar] [CrossRef]
- Welsh, J.A.; Jones, J.C.; Tang, V.A. Fluorescence and Light Scatter Calibration Allow Comparisons of Small Particle Data in Standard Units across Different Flow Cytometry Platforms and Detector Settings. Cytom. Part A 2020, 97, 592–601. [Google Scholar] [CrossRef]
- Shen, W.; Guo, K.; Adkins, G.B.; Jiang, Q.; Liu, Y.; Sedano, S.; Duan, Y.; Yan, W.; Wang, S.E.; Bergersen, K.; et al. A Single Extracellular Vesicle (EV) Flow Cytometry Approach to Reveal EV Heterogeneity. Angew. Chem. Int. Ed. 2018, 57, 15675–15680. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Ma, L.; Wang, S.; Chen, C.; Zhang, W.; Yang, L.; Hang, W.; Nolan, J.P.; Wu, L.; Yan, X. Light-scattering detection below the level of single fluorescent molecules for high-resolution characterization of functional nanoparticles. ACS Nano 2014, 8, 10998–11006. [Google Scholar] [CrossRef]
- Mizenko, R.R.; Brostoff, T.; Rojalin, T.; Koster, H.J.; Swindell, H.S.; Leiserowitz, G.S.; Wang, A.; Carney, R.P. Tetraspanin immunocapture phenotypes extracellular vesicles according to biofluid source but may limit identification of multiplexed cancer biomarkers. bioRxiv 2021, bioRXiv:2021.03.02.433595. [Google Scholar] [CrossRef]
- Price, J.M.J.; Hisada, Y.; Hazeldine, J.; Bae-Jump, V.; Luther, T.; Mackman, N.; Harrison, P. Detection of tissue factor–positive extracellular vesicles using the ExoView R100 system. Res. Pract. Thromb. Haemost. 2023, 7, 100177. [Google Scholar] [CrossRef]
- Hefley, B.S.; Deighan, C.; Vasini, B.; Khan, A.; Hjortdal, J.; Riaz, K.M.; Liu, Y.; Karamichos, D. Revealing the presence of tear extracellular vesicles in Keratoconus. Exp. Eye Res. 2022, 224, 109242. [Google Scholar] [CrossRef]
- Skovronova, R.; Grange, C.; Dimuccio, V.; Deregibus, M.C.; Camussi, G.; Bussolati, B. Surface marker expression in small and medium/large mesenchymal stromal cell-derived extracellular vesicles in naive or apoptotic condition using orthogonal techniques. Cells 2021, 10, 2948. [Google Scholar] [CrossRef]
- Baschong, W.; Stierhof, Y.-D. Preparation, Use, and Enlargement of Ultrasmall Gold Particles in Immunoelectron Microscopy. Microsc. Res. Tech. 1998, 42, 66–79. [Google Scholar] [CrossRef]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and tracking of tumour extracellular vesicle delivery and RNA translation using multiplexed reporters. Nat. Commun. 2015, 6, 7029. [Google Scholar] [CrossRef]
- Tian, Y.; Li, S.; Song, J.; Ji, T.; Zhu, M.; Anderson, G.J.; Wei, J.; Nie, G. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014, 35, 2383–2390. [Google Scholar] [CrossRef]
- Welsh, J.A.; Van Der Pol, E.; Bettin, B.A.; Carter, D.R.F.; Hendrix, A.; Lenassi, M.; Langlois, M.-A.; Llorente, A.; Van De Nes, A.S.; Nieuwland, R.; et al. Towards defining reference materials for measuring extracellular vesicle refractive index, epitope abundance, size and concentration. J. Extracell. Vesicles 2020, 9, 1816641. [Google Scholar] [CrossRef]
- Yamashita, T.; Takahashi, Y.; Nishikawa, M.; Takakura, Y. Effect of exosome isolation methods on physicochemical properties of exosomes and clearance of exosomes from the blood circulation. Eur. J. Pharm. Biopharm. 2016, 98, 1–8. [Google Scholar] [CrossRef]
- Phan, J.; Kumar, P.; Hao, D.; Gao, K.; Farmer, D.; Wang, A. Engineering mesenchymal stem cells to improve their exosome efficacy and yield for cell-free therapy. J. Extracell. Vesicles 2018, 7, 1522236. [Google Scholar] [CrossRef]
- Arab, T.; Mallick, E.R.; Huang, Y.; Dong, L.; Liao, Z.; Zhao, Z.; Gololobova, O.; Smith, B.; Haughey, N.J.; Pienta, K.J.; et al. Characterization of extracellular vesicles and synthetic nanoparticles with four orthogonal single-particle analysis platforms. J. Extracell. Vesicles 2021, 10, e12079. [Google Scholar] [CrossRef]
- Brittain, G.C.; Chen, Y.Q.; Martinez, E.; Tang, V.A.; Renner, T.M.; Langlois, M.-A.; Gulnik, S. A Novel Semiconductor-Based Flow Cytometer with Enhanced Light-Scatter Sensitivity for the Analysis of Biological Nanoparticles. Sci. Rep. 2019, 9, 16039. [Google Scholar] [CrossRef]
- Bachurski, D.; Schuldner, M.; Nguyen, P.; Malz, A.; Reiners, K.S.; Grenzi, P.C.; Babatz, F.; Schauss, A.C.; Hansen, H.P.; Hallek, M.; et al. Extracellular vesicle measurements with nanoparticle tracking analysis—An accuracy and repeatability comparison between NanoSight NS300 and ZetaView. J. Extracell. Vesicles 2019, 8, 1596016. [Google Scholar] [CrossRef]
- Cizmar, P.; Yuana, Y. Detection and Characterization of Extracellular Vesicles by Transmission and Cryo-Transmission Electron Microscopy. Methods Mol. Biol. 2017, 1660, 221–232. [Google Scholar] [CrossRef]
- Singh, R.K.; Santos, M.F.; Herndon, C.; Gieler, B.A.; Lee, I.; Chen, J.; Lorico, A. Detection by super-resolution microscopy of viral proteins inside bloodborne extracellular vesicles. Extracell. Vesicles Circ. Nucleic Acids 2023, 4, 557–567. [Google Scholar] [CrossRef]
- Silva, A.M.; Lázaro-Ibáñez, E.; Gunnarsson, A.; Dhande, A.; Daaboul, G.; Peacock, B.; Osteikoetxea, X.; Salmond, N.; Friis, K.P.; Shatnyeva, O.; et al. Quantification of protein cargo loading into engineered extracellular vesicles at single-vesicle and single-molecule resolution. J. Extracell. Vesicles 2021, 10, e12130. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ref. | Application | EV Source/ Nanoparticle | Targeting Molecule | Conjugation Method | Conjugation Confirmation Method | Reported Conjugation Efficiency |
|---|---|---|---|---|---|---|
| [38] | Alzheimer’s | Immature dendritic cells (DCs) | RVG; MSP; FLAG | Cell transfection with lamp2b | qPCR; WB | N/A |
| [30] | Lung Cancer | Red blood cells (RBCs) | EGFR-targeting peptide (ET); TR5 | Sortase A & OaAEP1 ligase-aided conjugation | WB (HRP serial dilution); Bead-based FC; Single EV FC | 380 ET peptides/EV; 77.2% TR5-ligated EV |
| [39] | Breast Cancer | HEK293 cells | GE11; EGF | Cell transfection with pDisplay | WB; Bead-based FC; Immunogold labeling + TEM | 15.3% (GE11); 21.2% (EGF) |
| [25] | Tumors/ Cancer | Neuro2A cells | EGa1 (anti-EGFR) nanobodies | Cell transfection, anchored to glycosylphosphatidylinositol (GPI) | WB; Immunogold labeling + TEM | 15–25% EGa1 |
| [40] | Tumors/ Cancer | RBCs; Neuro2A cells | EGa1 | Direct binding to phospholipid phosphatidylserine (PS) via lactadherin (C1C2) | WB; Immunogold labeling + TEM | Amount of EGa1-C1C2 conjugated to EVs increased in concentration dependent manner |
| [41] | Cardiovascular disease | Cardiosphere- derived cells | ASSLNIA (muscle homing peptide); CSTSMLKAC (ischemic targeting peptide) | Lipid insertion; Streptavidin-biotin binding | Cell Uptake | N/A |
| [42] | Melanoma | B16F10 cells | Cy3 fluorophore | Metabolic labeling with azide; Click chemistry | Cy3 absorption using Nanodrop; Bead-based FC | 790 nM AHA-Cy3/EVs (1mg/mL); 440 nM ManNAz-Cy3/EVs (1mg/mL) |
| [43] | Tumors/ Cancer | Neuro2A cells | EGFR ligand (EGa1) | Micelle formation by DSPE-PEG-maleimide + SH-ligand; Fusion | WB; Immunogold labeling + TEM | 7–14% EGa1; 0.4–4 ligands/EV |
| [44] | Cerebral Ischemia | Bone marrow mesenchymal stromal cells (BMSCs) | αvβ3-targeting peptide c(RGDyK) | Click chemistry; Amine crosslinking | Standard curve determined by FITC-labeled c(RGDyK) | 263 peptides/EV |
| [33] | Immunotherapy | Bovine serum | Mannose | Amine crosslinking (NHS-PEG-biotin); Lipid Insertion (DSPE-PEG-biotin or DSPE-PEG-mannose) | Gel electrophoresis; Biotin quantification kit | 40% NHS-PEG-bio on EVs; 70% DSPE-PEG-bio in EVs; ~12-250 nmol/mg EV (DSPE-PEG-biotin); ~40-400 nmol/mg EV (NHS-PEG-biotin) * depending on incubation ratios |
| [35] | Immunotherapy | THP1; J774A.1 cells | FasL; AS1411 aptamer; Cy5 | Cholesterol modification + ssDNA tether | Bead-based FC (CD63 antibody); Fluorescence microscopy | 1800–6900 ssDNA tethers/EV * depending on concentration of Chol-DNA |
| [45] | Abdominal aortic aneurysms | BMSCs | Cathepsin K binding peptide (CKBP) | Click chemistry; Amine crosslinking | Fluorescent plate reader (standard curve) | 0.22 μg CKBP per 10 μg EV |
| [36] | HER2-overexpressing cancer | Liposomes | Anti-HER2 Fab’ fragments | Lipid Insertion; Maleimide-thiol crosslinking to Fab’ fragments | Dye binding (protein concentration) assay | 100–120 Fab′ fragments/liposome |
| [28] | Muscular damage, inflammation, cirrhosis | Placental-derived MSCs (PMSCs) | SILY peptide | Click chemistry; Amine crosslinking chemistry | Fluorescence microscopy; ExoView | 70% SILY peptide |
| [27] | Myeloid leukemia | RBCs | B-TL5; B-T140 | OaAEP1 ligase-aided conjugation; Streptavidin-biotin binding | WB (HRP serial dilution); Single EV FC; Competitive ELISA | 95% B-TL5; 99% B-T140; 351 B-TL5/EV; 1000-1402 B-T140/EV |
| [37] | Tumors/ Cancer | Food-derived (milk & plant cells) | Alexa Fluor 555; Transferrin | Disulfide reduction via mild reducing agent TCEP; Thiol-maleimide conjugation | Fluorescence spectrometry + NTA; ELISA; NanoFCM | Max of 1965/mEV; 85% Alexa Fluor 555; 74% Transferrin |
| [46] | Breast Cancer | HEK293FT cells | Tumor-homing peptides (THPs) – PDL1; uPAR; EGFR | Lipid Insertion; Amine-crosslinking chemistry | N/A | N/A |
| [47] | Liver Cancer | HepG2 cells | Nanoassembly (NA)-(HepG2 EV-binding aptamer LZH8 & M1/M2 monomers) | HepG2 EV-binding aptamer LZH8; DNA Hybridization Chain Reaction (HCR) | Bead-based FC; Immunogold labeling + TEM | N/A |
| [29] | Cancer | Sf9 cells | Programmed cell death 1 (PD-1) protein; Baculoviral fusogenic glycoprotein gp64 | Cell transfection with plasmid transformed into recombinant baculoviruses | WB; Imaging FC (IFC) | 37% fusion efficiency with PD-1 EVs and cargo-loaded liposomes |
| [31] | Breast Cancer | PLGA–PEG nanoparticles (NP) | Monoclonal antibody TRAZ (antibody fragment) | Disulfide-selective pyridazinedione linkers; Click chemistry | BCA (standard curve); Surface plasmon resonance (SPR) | 193.1 TRAZ F(ab) pmoles/mg PLGA-PEG NPs; 18.4% modified TRAZ F(ab) |
| [32] | Cancer | Polyion complex (PIC) micelles | Anti-EphA2 (antibody fragment) | Maleimide-thiol crosslinking; Click chemistry | UV-vis absorbance | 1.5–3.5 Fabs/micelle |
| [48] | Cancer Immunotherapy | B16BL6 cells | CpG DNA | Streptavidin (SAV) cell transfection; Streptavidin-biotin binding | Fluorescence microscopy | 287 CpG DNA molecules/EV |
| [34] | Colorectal Cancer | BM-MSCs | MUC1 aptamer | EDC/NHS chemistry | Nanodrop; Gel retardation assay (gel electrophoresis) | 44.78 μg MUC1 aptamer/100 μg EV |
| [24] | Osteoporosis | RBCs | TBP | CD63 receptor binding through CP05 peptide | Bead-based FC | N/A |
| [26] | Cancer | Immature mouse dendritic cells | Nucleolin-targeting aptamer AS1411 | Cholesterol anchor on cells; Extrusion to form exosome-mimetic extracellular nanovesicles (ENVs) | Fluorescence microscopy & Dot blot using Cy5-labeled complementary DNA probe of AS1411 | 15–25% on cells prior to ENV extrusion * depending on chol-PEG2000 concentration |
| [49] | General targeting | 4T1 cells | Fluor 545 | EDC/NHS chemistry; Click chemistry | HPLC/UV–vis absorbance | 1.5 alkynes available to react to azide-fluor 545/EV (150kDa protein) |
| [50] | Osteoporosis and fracture | BMSCs | BMSC-targeting DNA aptamer | Amine-based Schiff base reaction | Bead-based FC; Cell Uptake | N/A |
| Technology | Detection Thresholds (nm) |
|---|---|
| SP-IRIS (ExoView) | 50 nm |
| Single Flow Cytometry (CytoFLEX) | 70 nm |
| NanoFCM | 40 nm |
| Conventional Flow Cytometers (Bead-based FC) | 200 nm |
| TEM | 1 nm |
| NTA | 70 nm |
| MRPS | 50 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldbloom-Helzner, L.; Bains, H.; Wang, A. Approaches to Characterize and Quantify Extracellular Vesicle Surface Conjugation Efficiency. Life 2024, 14, 511. https://doi.org/10.3390/life14040511
Goldbloom-Helzner L, Bains H, Wang A. Approaches to Characterize and Quantify Extracellular Vesicle Surface Conjugation Efficiency. Life. 2024; 14(4):511. https://doi.org/10.3390/life14040511
Chicago/Turabian StyleGoldbloom-Helzner, Leora, Harjn Bains, and Aijun Wang. 2024. "Approaches to Characterize and Quantify Extracellular Vesicle Surface Conjugation Efficiency" Life 14, no. 4: 511. https://doi.org/10.3390/life14040511
APA StyleGoldbloom-Helzner, L., Bains, H., & Wang, A. (2024). Approaches to Characterize and Quantify Extracellular Vesicle Surface Conjugation Efficiency. Life, 14(4), 511. https://doi.org/10.3390/life14040511